
杭州午潮山天然次生林木本幼苗组成及其时空分布格局
2022-08-31 06:08:30岳春雷李贺鹏
生态学报 2022年15期
朱 弘,杨 乐,岳春雷,李贺鹏
浙江省林业科学研究院,杭州 310023
森林的更新和演替是一个复杂的生态学过程,也是森林生态领域长期关注的热点问题之一[1]。其中,木本植物实生幼苗天然更新是森林植被恢复与演替的主要方式,木本幼苗通过定植、生长、扩散,对未来森林群落的物种构成、种群结构、分布格局与演替进程带来深远的影响[2]。森林更新受树种生理特性、地理环境、自然干扰、人为干扰等因素及其相互作用的影响,这些生物的和非生物的因素随时间和空间而不断变化,构成了森林的空间异质性和时间异质性,使森林更新具有时空变化的特点[3]。
杭州午潮山开发利用历史悠久,近代由于过度森林砍伐和不合理的土地利用活动,导致区域内原始植被遭到了严重破坏;1960年后,浙江省林业科学研究院将其作为科研基地加以保护,经过长期的封山育林,演替形成了以木荷(Schimasuperba)、青冈(Cyclobalanopsisglauca)、苦槠(Castanopsissclerophylla)等为建群种的浙北典型天然次生林[4]。近年来,有关午潮山森林的群落结构、物种组成、功能性状等方面已经取得了一定的进展[5—6],然而相关研究主要针对乔木大树,对其林下幼苗的关注度明显不足,特别是在大样地的实践中,往往被选择性忽视,从而限制了对该区域森林生态系统的全面认识。
幼苗阶段被认为是森林演替过程中最重要和最敏感的阶段,其种类构成、数量和存活动态等方面的变化也最为明显[7]。基于幼苗层物种的空间分布规律和自然更新的探究,有助于预测森林演替方向和进程[8—10]。鉴于此,本研究重点以杭州午潮山天然次生林天然更新幼苗为研究对象,关注以下3方面问题:1)林下木本幼苗物种组成和数量特征;2)主要优势树种幼苗基径结构;3)优势木本幼苗在不同发育阶段的空间分布规律和小尺度空间效应。研究结果旨在为为丰富北亚热带森林物种多样性构建与群落演替机制资料,也为加快该区域的植被恢复重建和森林经营管理提供依据。
1 材料与方法
1.1 研究区概况
研究区地处杭州西郊,属天目山余脉,平均海拔264 m,主峰504 m。气候带属亚热带季风气候,其中年平均气温15.4℃,极端最高温39℃,极端最低温-9.4℃,年均降水量为1 992.5 mm,年无霜期240 d,年平均相对湿度81%;森林植被类型涵盖常绿阔叶林、落叶阔叶林、针阔混交林、针叶林、竹叶林和灌木林6大类型,森林覆盖率达98.2%。土壤类型为红壤[6,11]。
1.2 样地设置和幼苗调查
2021年3—4月,在前期林分踏查的基础上,从幼苗分布较为丰富的典型沟谷地(30.19°N,120.00°E,海拔237 m)设置一个与底部溪流方向大致平行的矩形样带,再在样带内采用相邻格子法划分出6个5 m × 5 m的小样方,调查面积共计150 m2。以样方左下角为坐标原点建立直角坐标系,调查和记录样方内胸径(Diameter at Breast Height,DBH)<1 cm的林下所有幼苗的物种名称和空间二维坐标(x,y),并利用卷尺和数显游标卡尺(桂林广陆数字测控有限公司)测量每个个体的株高和基径(Stem base diameter, SBD)。
1.3 数据处理
1.3.1幼苗数量特征
幼苗密度=幼苗总株数/样方面积(m2)
(1)
幼苗丰富度=物种数/样方面积(m2)
(2)
幼苗重要值(IV)=(相对多度+相对频度)/2
(3)
式中,相对多度(RA)=(某个种的株数/所有幼苗总株数) × 100%,频度(F)=某物种出现的样地数/样地总数× 100%,相对频度(RF)=(某一物种的频度/所有物种的频度之和) × 100%[12—13]
1.3.2优势木本幼苗的确定和不同径级结构的划分
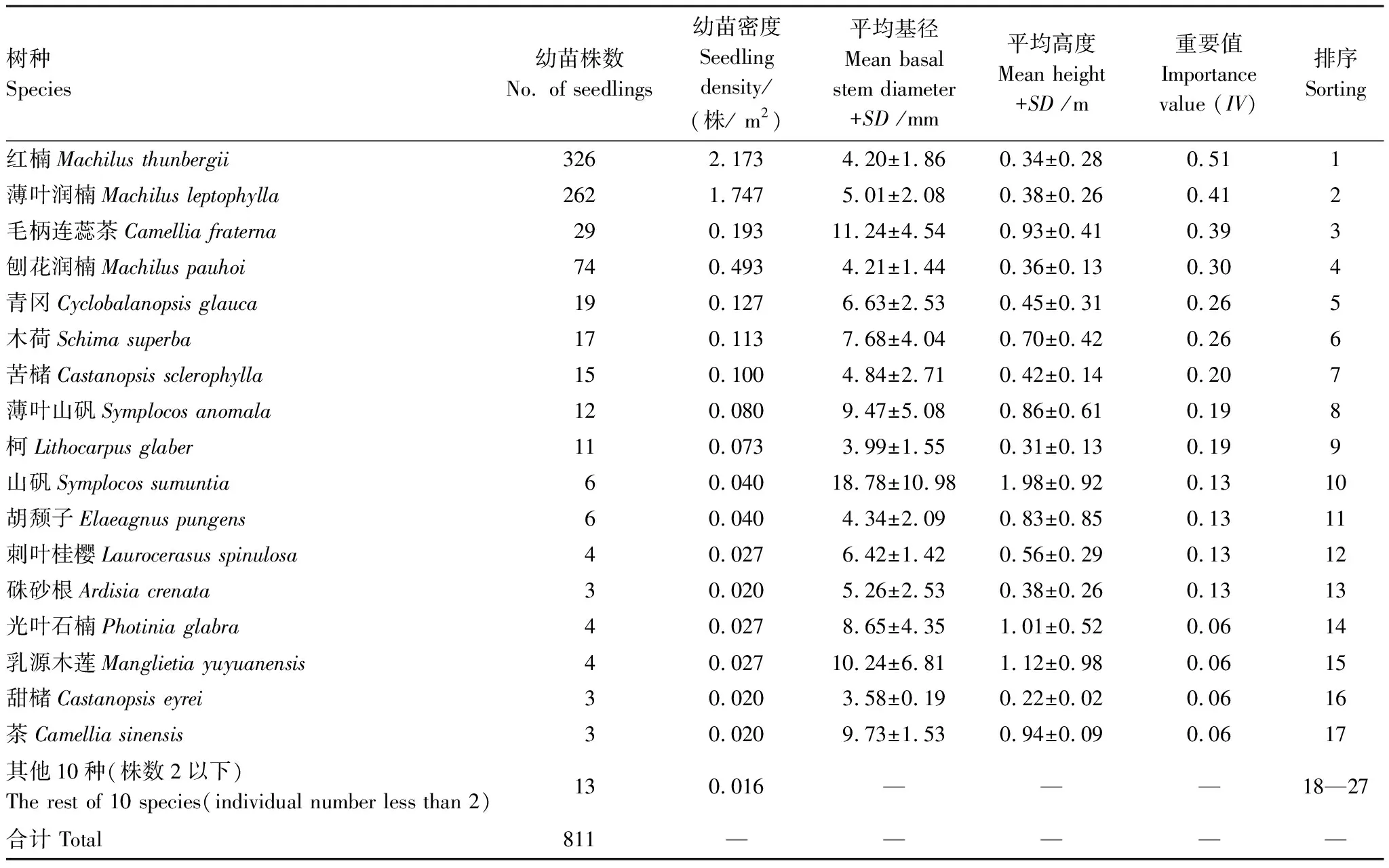
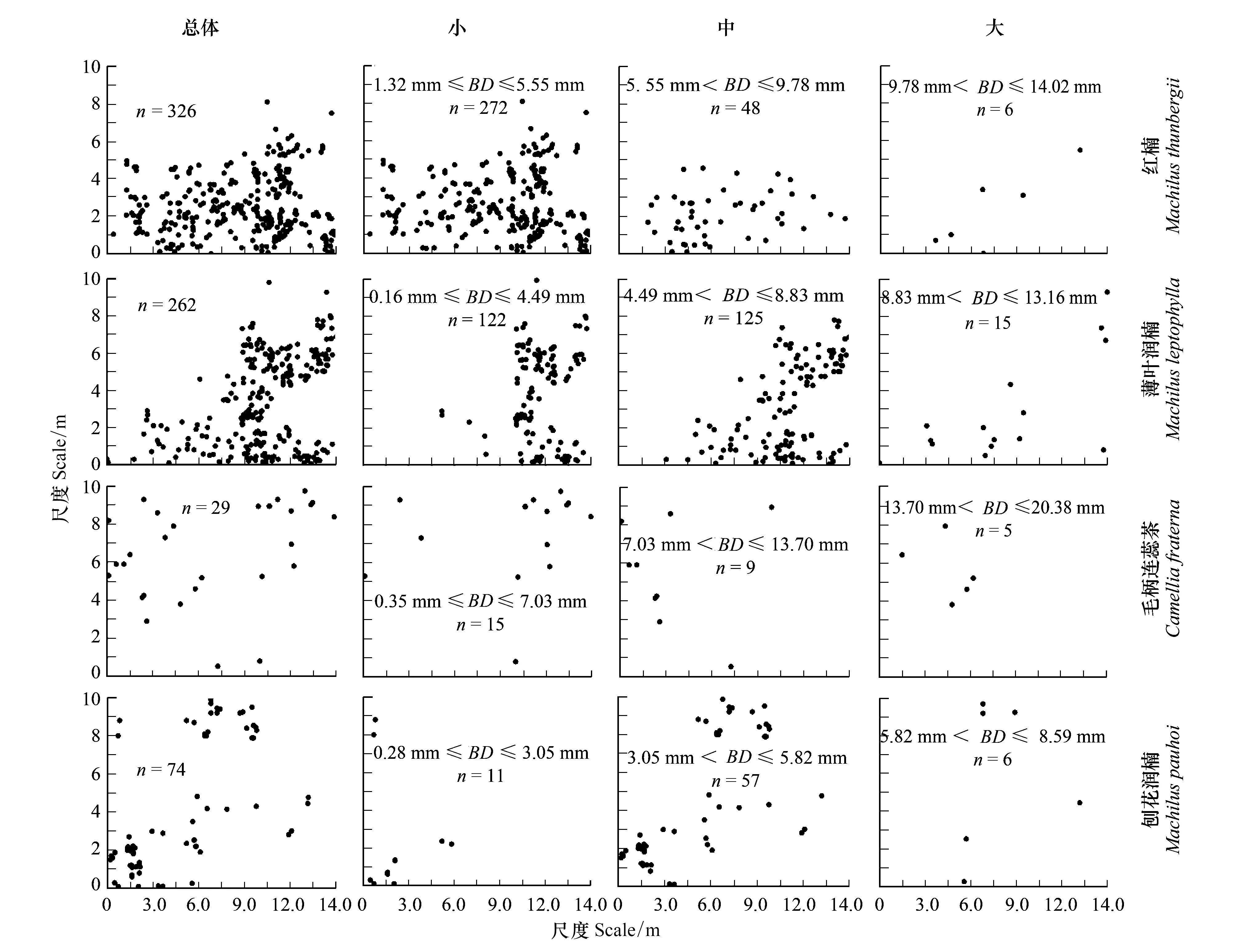
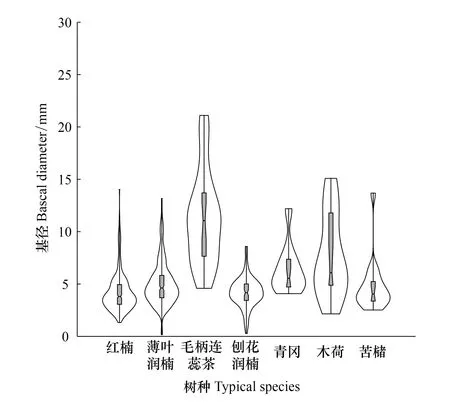
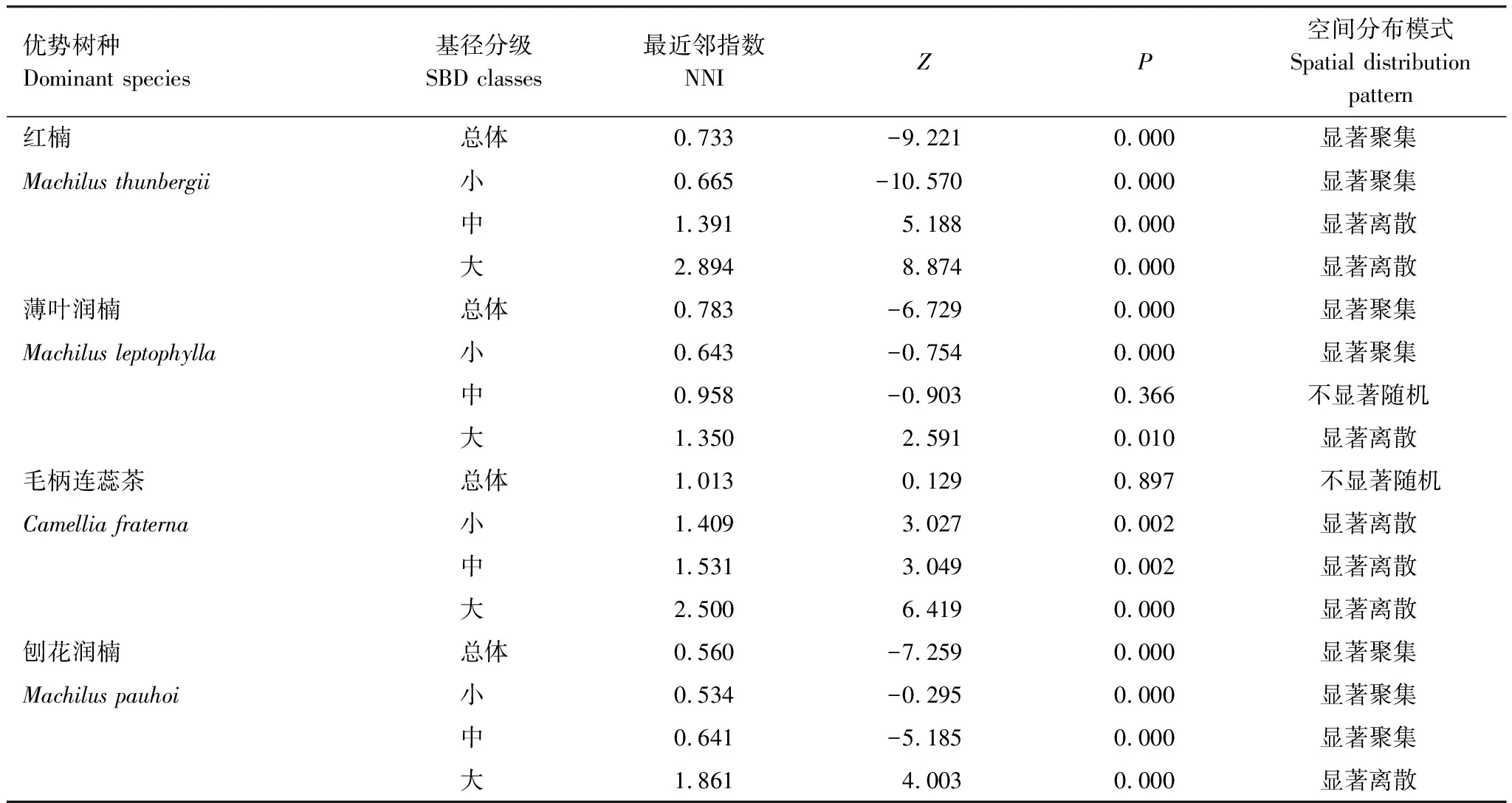
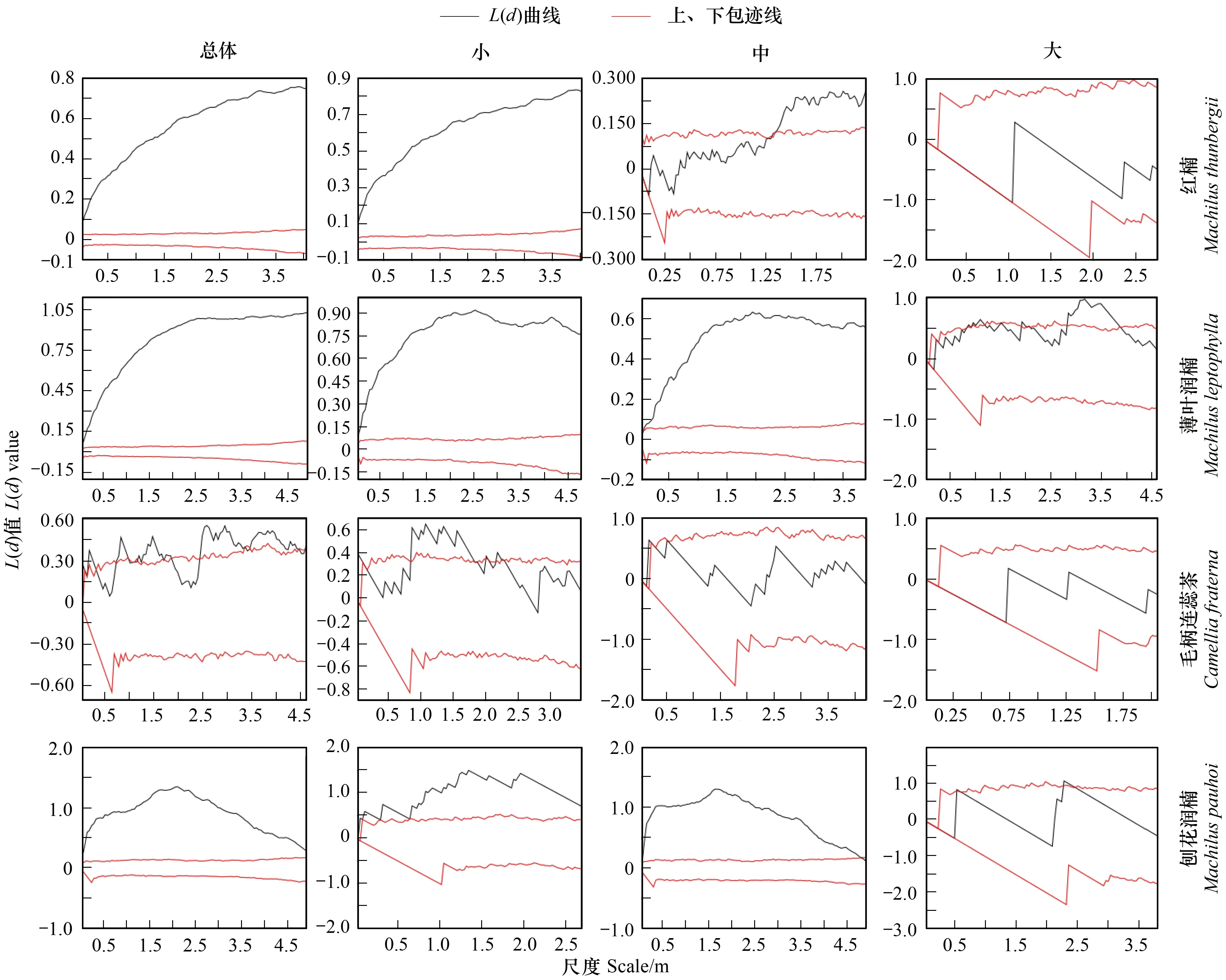
按重要值大小进行排序(表1),筛选IV≥0.3的物种作为优势木本幼苗作为后续研究对象。依据“空间替代年龄”的原理和方法,即将基径从小到大的顺序视为时间顺序关系,首先绘基径制频率直方图(Frequency histogram),将频率分布较大转折点作为龄级划分的标准[14],同时结合样地内幼苗个体数及自身特性的实际情况,最终将不同优势树种基径(BD)划分为小径级(Small-size classes)、中径级(Medium-size classes)和大径级(Large-size classes)3个径级,视为不同生长发育阶段。其中红楠(Machilusthunbergii)3个径级依次为:1.32 mm≤BD≤5.55 mm、5.55 mm 1.3.3最近邻分析 利用最近邻分析(Nearest Neighbor Analysis, NNA)法分析不同物种在不同径级下的空间分布格局。它最早于1954年由密歇根大学的Clark和Evans提出[15],是一种经典的点格局分析方法。其原理是将随机分布模式下的预期最近邻距离与实际观测得到的最近点的平均距离进行比值,用得到的最近邻指数(Nearest neighbor index, NNI)来作出种群的分布类型[14]。公式如下: (4) 式中,A是研究区的总面积,di是i第棵树与其最近树之间的距离,n为每类树的总株数。当NNI接近1时,呈随机分布;NNI<1时,呈聚集分布;NNI>1时,呈离散分布。为了更好地反映实测平均距离和预期平均距离的偏离程度,利用正态分布检验得出Z值及其95%置信水平。Z值为负且值越小,个体越趋于聚集分布;Z值正值且越大则显著偏向均匀分布;Z值位于二者之间则为随机分布。 1.3.4Ripley′s K函数分析 由于种群空间格局特征具有很强的尺度依赖性,因此进一步利用Ripley′sK函数对不同物种在不同距离尺度下的空间分布格局进一步分析。该方法自1977年由Ripley[16]首次提出,已经在国内外广泛使用。公式如下: (5) 式中,d为空间尺度,A为研究样地面积,n为样地内某一种植物的个体总数,uij为个体i到个体j之间的距离,Wij为边界效应校正系数。当uij≤d时,Ir(uij)=1;当uij>d时,Ir(uij)=0;为保持方差稳定,对K(d)作开平方的线性变换如下: (6) 在随机分布下,L(d)在所有空间尺度下均等于0;当L(d)>0,表示幼苗个体服从聚集分布;当L(d)<0,表示幼苗个体服从均匀分布。 将实测格局与选定的零模型(null models)通过Monte-Carlo法绘制空间格局,拟合上、下两条包迹线,重复验证99次,确定95%的置信水平。用实际数据计算不同尺度的L(d)值,若L(d)在包迹线以内,则呈随机分布;若L(d)值在包迹线以上,则呈显著聚集分布;若L(d)在包迹线以下,则呈显著均匀分布;上述两种点格局分析法的运行和出图均在PAST version 4.03统计软件包(http://folk.uio.no/ohammer/past/)中实现。 研究样地内共记录到811株实生幼苗,隶属12科20属27种(表1),其中樟科(Lauraceae)(5)、壳斗科(Fagaceae)(4)、山茶科(Theaceae)(3)和山矾科(Symplocaceae)(3)为排名前4的科;前4个优势种(IV≥0.3)为红楠、薄叶润楠、毛柄连蕊茶、刨花润楠累计个体总数为691株,占样地内所有幼苗总株数的85.20%;优势种各级散布模式和聚集程度明显不同(图1);样地总体幼苗密度和幼苗丰富度分别为5.4株/m2和0.18种/m2。 表1 午潮山林下幼苗物种组成及数量特征 图1 午潮山150 m2样地内4个优势木本幼苗不同基径结构的空间分布图Fig.1 Spatial distribution of four dominant tree seedlings at different growth stages in Wuchao Mountain with 150 m2 plot 从主要优势木本幼苗基径分布的小提琴图(图2),可以看到不同树种基径分布有一定的差异:红楠和薄叶润楠总体基径呈典型的正偏态分布,差别在于红楠小径级个体数量优势明显占其总株数的83.44%,预示着其种群结构属于快速增长型;而薄叶润楠小径级与中径级的比例接近1∶1,两端存在更多的离散值,种群结构属于稳定增长型;毛柄连蕊茶总体呈现略微正偏分布的双峰型,各径级数量分布较为分散,小基径占其总株数的51.72%,且起始值较大,种群结构属于缓慢增长型;刨花润楠总体基径近似正态分布,中径级占其总株数的77.02%,种群结构属于稳定型。 图2 午潮山林下代表性木本幼苗的基径分布的小提琴图 Fig.2 Violin plots of stem base diameter (SBD) classes distribution of understory seedling for typical tree species in Wuchao Mountain 利用最近邻分析分别对4个优势种3个径级的空间分布格局进行空间分布格局的判定,结果(表2)表明:红楠总体和小径级的NNI指数<1,Z值均为负值,在空间上呈显著聚集分布;中径级和大径级的NNI指数>1,Z值均为正值,在空间上呈显著离散分布。薄叶润楠总体和小径级的NNI指数<1,Z值均为负值,在空间上呈显著聚集分布;中径级的NNI指数接近1,Z值为负值,未能拒绝在P<0.05的水平上随机零假设,在空间上呈不显著随机分布;大径级NNI指数>1,Z值均为正值,在空间上呈显著离散分布。毛柄连蕊茶总体的NNI指数接近1,Z值为正值,未能拒绝在P<0.05的水平上随机零假设,在空间上呈不显著随机分布;小、中、和大径级的NNI指数均>1,Z值均为正值,在空间上呈显著离散分布。刨花润楠总体、小径级和中径级的NNI指数均<1,Z值均为负值,在空间上呈显著聚集分布;大径级的NNI指数>1,Z值均为正值,在空间上呈显著离散分布。 表2 午潮山4个优势木本幼苗在不同生长阶段的最近邻分析结果 Ripley′sK函数分析结果(图3)显示,红楠总体和小径级在0—4 m范围内均呈聚集分布;中径级在0—1.25 m范围内呈随机分布,在>1.25 m呈聚集分布;大径级在0—2.5 m范围内均呈随机分布。薄叶润楠总体和小径级在0—4.9 m范围内均呈现强烈聚集分布;中径级在0—3.9 m范围内呈随机分布;大径级在0—2.8 m和3.8—4.5 m范围内呈随机分布,在2.8—3.8 m范围内呈聚集分布。毛柄连蕊茶总体在0—4.5 m范围内呈集中和随机的波动模式;小径级在0.8—2.2 m范围内呈集中分布,伴有少量随机分布;中径级和大径级分布在0—4 m和0—2 m范围呈随机分布。刨花润楠总体和中径级在0—5 m范围内呈集中分布,当d=2 m左右,L(d)=1.23,聚集强度分别达到最大;小径级在0—2.7 m范围内呈集中分布;大径级基本在0—3.9 m范围内呈随机分布。 图3 午潮山样地内4个优势木本幼苗不同径发育阶段的Ripley′s K函数分析Fig.3 The Ripley′s K analysis of four dominant tree seedlings at different growth stages in Wuchao Mountain 研究森林群落组成、结构随演替的变化规律,对于揭示森林群落构建与物种共存机制具有重要意义[17—20]。基于小尺度的样方调查表明,午潮山乔木幼苗树种组成较为丰富,群落整体分布密度较高,表明林下自然更新良好。但不同种间的个体数量分配差异较大,表明不同种的繁殖能力与环境适应是不同的。径级结构在一定程度上反映植物种群的年龄结构特征和生长发育状况,成为预测种群和群落的动态发展趋势的一种有效方法。在所有调查的木本幼苗中,红楠和薄叶润楠是重要值排序前两位的树种,也是中至北亚热带的优势种[21],由于储备较多的小径级幼苗个体,属于增长型种群,或将成为下一阶段午潮山群落演替的建群树种。毛柄连蕊茶是亚热带山区常见伴生种,通常以常绿灌木或小乔木的形式生长,也是早春重要的蜜源植物之一[22]。依靠其显著的基径和高度,重要值排名维持至第3,虽然个体数量不多,但生长良好,推测未来仍将作为林下伴生树种与其它物种长期混交共存。刨花润楠的重要值排名第4,该种是亚热带重要的山地乡土树种,亦是集经济用材价值和园林观赏价值为一体的珍贵特色树种[23],其幼苗各径级呈近正态分布的样式反映出该种生长发育对本区域有着很好的适应性。总之,午潮山木本幼苗是午潮山森林资源的重要组成部分,上述优势物种通过占据不同水平与垂直空间,采取不同的生活史对策,在群落中得以共存,并维持群落的稳定性。 种群分布格局的变化反映了植物生存策略、适应机制,进而影响群落的发展趋势[24—27]。利用最近邻分析可以分析全局尺度下树种的空间分布格局,而基于Ripley′sK函数分析能够进一步量化研究区域内植物个体在不同空间尺度上的分布格局,从而揭示植物种群的生态学特性和形成机制[28]。在本研究中,红楠、薄叶润楠和刨花润楠3个润楠属在整体和小径级个体均呈聚集分布,随着径级和空间尺度的增大,聚集强度达到最强后逐渐降低,常呈随机或均匀分布。说明不同树种在不同发育阶段、不同龄级也会形成不同的空间格局,这通常是自身生物学特性和环境因素共同作用的结果。可能的原因如下:首先,3种植物均隶属于樟科润楠属植物,幼苗期耐阴蔽,喜湿润肥沃土壤,研究地处低海拔沟谷,水热充沛,较厚的林下凋落物层有利于种子冬季储存及打破生理休眠,从而获得较高的种子萌发率,促进幼苗的集中大量萌发和生长;其次,核果球形,推测扩散传播以重力传播和水流传播为主,因而野外观察到大部分种子集中散落在母树周围及沿河流沟谷两侧;此外,3种植物低龄级幼苗个体较小,处在森林群落的下层,对光照、水分、土壤等环境因子的竞争能力较弱,聚集分布的幼苗在母树遮蔽下可以形成幼苗群,能够形成有利于其生长发育的微环境,从而提高个体竞争能力及成活率。但是当种群年龄和空间尺度逐渐增大,种子有效传播能力和繁殖能力不断减弱,种内种间对资源和空间的需求不断激增,加上病虫害、草食动物取食用等侵扰加剧带来的负密度制约效应影响[29],导致大量个体死亡,种群密度下降,聚集强度随之减弱,种群由聚集分布向随机分布或离散分布转变[30]。此外,还可以看到,红楠与薄叶润楠的分布格局较为接近,而与刨花润楠相差较大,推测前两者具有更相似的生物学特征,如均耐阴喜湿、更大的比叶面积、幼枝轮生等特征,相似的生态位导致生境选择的趋同,从而导致两者常常相伴而生。 相较而言,毛柄连蕊茶的个体数量较少、种群密度较低,分布格局由整体上的不显著随机分布向显著离散分布转变,这一现象的产生,一方面,受限于自身较慢的生长速率,导致其在幼苗层群落中的竞争力有所弱化,林中大量草本和藤本生长占据地表空间,使得许多的毛柄连蕊茶小径级幼苗容易被掩盖,影响生长发育。另一方面,由于常年处于林冠下层,郁闭度较高,在授粉期光照不足情况下,毛柄连蕊茶植株的开花结实率有下降的可能。此外,所在生境还频繁受到人为干预,如当地林场工人的砍伐、挖笋、采茶等活动导致小苗很难正常生长,最终影响了该物种种群结构的正常演替。 综合上述研究结果,建议在森林经营过程中:1)可对小尺度范围内集中分布和长势不良(如病虫害或者顶芽受损)的红楠和薄叶润楠幼苗进行合理间伐,降低小径级阶段自疏的比例,提高中、高径级的比例。2)对林下郁闭度较高、枯死木、病株、攀援藤本(如南五味子Kadsuralongipedunculata、鸡矢藤Paederiascandens等)所在区域进行及时清理,有利于林窗的快速形成,从而提高林下木本幼苗(如毛柄连蕊茶)的更新率和成活率。3)对壳斗科、山茶科等亚热带常绿阔叶树种(如表2中重要值排序稍靠后的青冈、苦槠、木荷等树种)幼苗进行适当抚育或补植工作,优化种群结构,加速群落向顶级群落的演替过程,最终促进整体森林群落结构的正向更新,提高群落的稳定性与物种多样性。2 结果与分析
2.1 林下幼苗组成与数量特征
2.2 优势树种幼苗的基径结构
2.3 优势种幼苗不同径级个体的空间分布格局
2.4 优势种幼苗不同径级个体多尺度特征分析
3 讨论
猜你喜欢
广西林业科学(2022年4期)2022-11-07 01:50:34
林业调查规划(2022年3期)2022-06-13 09:31:04
广西林业科学(2021年6期)2022-01-21 07:39:10
四川林业科技(2019年5期)2019-07-07 07:14:30
安徽农学通报(2019年1期)2019-02-14 02:25:08
湖北林业科技(2019年6期)2019-01-06 02:14:29
老年教育(2018年12期)2018-12-29 12:43:00
现代园艺(2017年19期)2018-01-19 02:49:51
现代园艺(2017年21期)2018-01-03 06:41:58
湖北林业科技(2017年2期)2017-05-25 18:44:32
