
不同地理种源杉木根叶功能性状与碳氮磷化学计量分析
2022-08-31 06:08:36王利艳马祥庆
生态学报 2022年15期
徐 睿,刘 静,王利艳,颜 耀,马祥庆,2,李 明,2,*
1 福建农林大学林学院,福州 350002 2 国家林草局杉木工程技术研究中心,福州 350002
植物功能性状是指植物体所具有的一系列与其定植、存活、生长和死亡密切相关的核心植物属性[1]。作为连接植物与环境的桥梁,植物功能性状反映了植物不同物种或种群在长期进化过程中对生存环境的生长适应和竞争能力[2]。植物中碳(C)、氮(N)磷(P)元素的化学计量特征能够反映植物的生长速率、养分利用效率和限制性元素及植物自身随环境的变化,是探讨生态系统和植物关系的新思路[3- 4],也为研究植物应对不同生境条件的功能性状权衡关系拓宽思路,因此,近年来被频繁应用于植物功能性状的研究中。地理分布广泛的同种植物在漫长的进化和演替过程中,会对长期气候地理环境变化产生适应性响应,从而形成特定的变异类型或地理种源,这也体现了植物对生长环境的长期适应策略[5]。植物比叶面积、比根长等功能性状和C、N、P含量及其化学计量比在不同地理生境和不同物种之间变化很大,这些功能性状的变化,体现了植物体在生境差异较大的不同地理种源之间的适应性分化和表型可塑性。
植物根系和叶片是植物最为重要的功能器官,其功能性状和碳氮磷化学计量特征差异最能反映植物对异质性生存环境的适应策略和竞争能力[6]。C、N、P是植物体所必须的生命元素,也是植物生长发育的关键性限制元素,C、N、P化学元素在植物生长代谢水平上的限制作用可通过化学计量来衡量[7- 8]。植物根系既要为植物从土壤中吸收养分和水分,又要将光合作用的产物转化成有机质返还土壤[3]。当土壤环境发生变化时,根系会通过改变自身形态生理性状的方式来提高对其土壤养分的吸收能力[9],同时也会使植物根系的化学计量随之改变[10],从而有效提高植物对环境的适应性。尤其以直径小于2mm的细根对外界环境响应最为敏感,也最能反映植物根系随外界环境的变化[11]。叶片在植物各功能器官中与环境接触面积最大、对环境响应最敏感[12],可塑性较大,易受环境因子影响[13]。叶片也是光合作用发生的器官,对植物的养分循环以及贮存养分有着重要作用[14]。植物叶片功能性状具有相对稳定性、容易测量、并能很快量化的特性,且能反映植物对各种环境因子的生态适应性[15],叶片结构和化学计量特征是反应植物生长地理环境的关键因素[16]。
杉木(Cunninghamialanceolata)是我国南方人工造林的重要树种,水平分布在我国整个亚热带、热带北缘、暖温带南缘等气候区的19个省份,垂直分布在海拔130- 2900m的丘陵山地,其面积和蓄积量约占全国人工乔木林主要优势树种的1/5和1/4[17- 18]。目前杉木地理分布格局并不是按照经度和纬度进行地带性分化,而是表现出“多中心起源”的特点,俞新妥等将全国杉木划分为9个种源区和“三带五区”的分布区[19]。不同种源杉木的生长、叶片光合、木材密度、养分利用效率等功能性状均存在较大差异[20]。这些不同地理种源杉木功能性状的差异,其实质可能为历史上分布极为广泛的杉木对不同生存环境的长期适应性响应而产生的分化。以往对不同地理种源杉木的研究中,通常较为关注其引种栽培后的速生性状差异,而忽略了不同种源间根、叶功能性状及化学计量特征的差异,也缺乏对这些性状特征与环境因子关系的探讨。因此,本研究选取采集自树龄百年以上、种源地较为明确的杉木王群体同质园扦插苗为研究对象,对7个不同地理种源的3年生杉木幼树进行根、叶功能性状及C、N、P化学计量分析,以期解决以下3个问题:(1)种源地气候是否可以解释不同种源的杉木根、叶主要功能性状与C、N、P计量的差异?(2)主要功能性状间是否存在权衡关系? (3)主要功能性状与C、N、P计量是否受种源地气候环境的影响?杉木王记录了当地气候环境因子的变化,形成了适应当地地理环境的独特功能性状和化学计量特征,对这些性状特征与气候环境因子相关性的分析能够探究不同种源杉木的适应性特征和功能性状权衡关系,也有助于认识当前杉木地理分布格局的形成原因[21]。
1 研究方法
1.1 样品地理信息
试验所选用材料于2016年采集自7个不同地理种源的杉木王,2016年,取不同地理种源杉木王顶端带顶芽的2- 3节作为插穗,带回福建农林大学同质园圃进行扦插,将插穗根部蘸浓度为100mg/L的ABT生根粉的泥浆后,插入营养土育苗基质,等苗高达40cm后进行栽植。该同质园圃位于杉木中心产区,属中亚热带季风气候,温暖湿热、降水充沛、光照充足、土层深厚,适宜于杉木生长。本研究选取对杉木生长与分布有重要影响的种源地经纬度、海拔、积温、降水等气候环境因子进行功能性状与种源地地理环境因子的相关性分析。样品种源地经纬度和海拔通过手持GPS数据测量获得,气象数据来源于国家气象局气象数据中心(http://data.cma.cn/)采样点气象数据的近30年平均值获得(表1)。
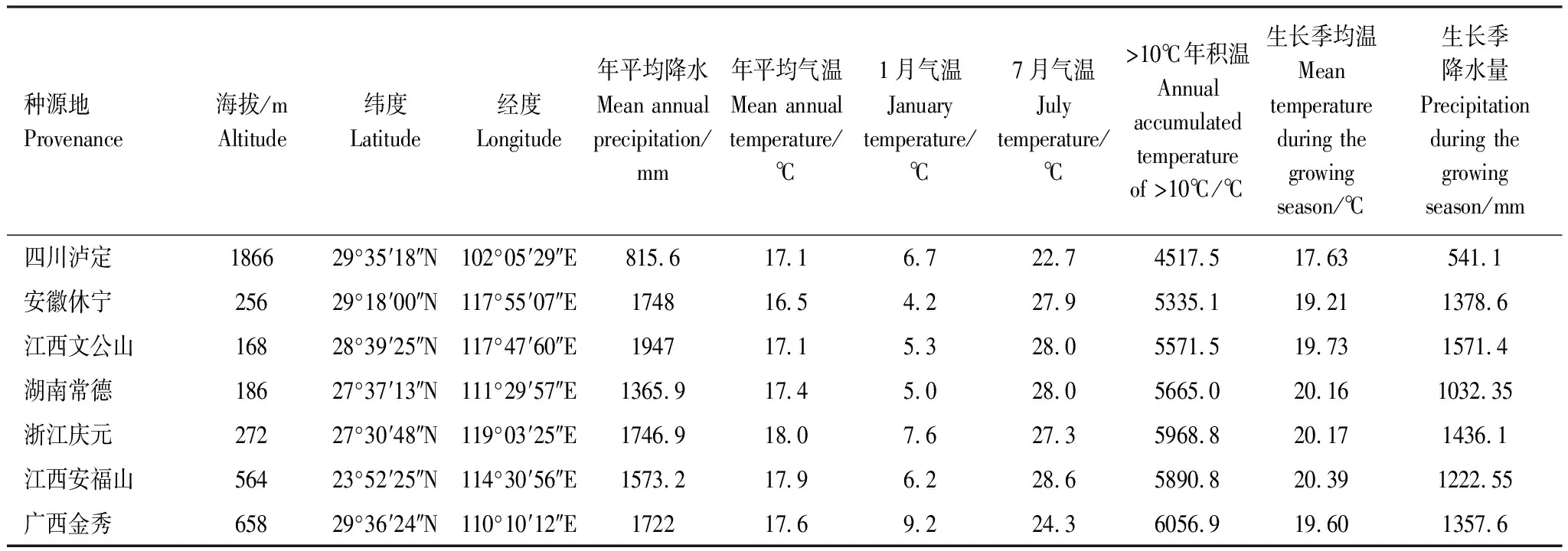
表1 不同杉木王及种源地特征
1.2 样品采集
2019年11月在3年生的不同种源杉木同质园圃中分别选取5株标准株,每株取从顶端向下数的第2节上20片中等大小的健康叶片用于叶片功能性状和C、N、P元素含量测定。采用收获法采集标准株直径小于2mm的细根,用于根功能性状测量及C、N、P元素含量测定。
1.3 功能性状的测量
使用千分之一天平测量杉木叶饱和鲜重,再使用ImageJ分析叶片扫描图以获得叶面积。叶片经80℃烘干至恒重后测量干重,计算比叶面积、叶组织密度和比叶重。其中,比叶面积(Specific leaf area,SLA)=叶面积/叶干重,叶组织密度(Leaf tissue density,LTD)=叶干重/(叶面积×叶厚度)[22],比叶重(Leaf mass per area,LMA)=叶干重/叶面积,叶干物质含量(Leaf dry matter content,LDMC)=叶干重/叶饱和鲜重[23]。使用千分之一天平称量杉木根饱和鲜重,再使用WINRHIZO根系分析系统扫描分析根表面积。根经80℃烘干至恒重后测量干重,计算比根长、根比表面积和根干物质含量。其中,比根长(specific root length,SRL)=根长/根干重[24],根比表面积(root specific surface area,RSA)=根表面积/根干物质重,根干物质含量(Root dry matter content,RDMC)=根干重/根饱和鲜重[25]。
1.4 C、N、P含量测定
烘干至恒重后的杉木根、叶样品磨碎成粉末,后过0.15mm筛,使用Vario Max碳氮元素分析仪(德国Elementar)测定C、N含量,并计算碳氮比。粉碎过筛后的根、叶样品使用ETHOS UP微波消解仪(美国Milestone)加入硝酸5mL、30%过氧化氢1mL进行消解后,使用钼锑抗比色法测得全磷含量,并计算碳磷比和氮磷比。
1.5 数据分析
采用SPSS 22.0和Canoco 5 软件对数据进行统计分析。采用单因素方差分析(one-way ANOVA) 和Duncan法进行多重比较(α=0.05)对不同地理种源杉木王幼树功能性状进行显著性分析,用Pearson 法分析各功能性状间的相关性,用主成分分析法(PCA)筛选出对杉木王幼树影响较大的代表性环境因子,用冗余分析(RDA)分析各环境因子对杉木王幼树功能性状的影响。利用SPSS 22.0和Canoco 5 软件作图。图表中数据为平均值±标准差。
2 结果与分析
2.1 不同地理种源杉木王幼树功能性状及C、N、P化学计量特征差异
对不同地理种源杉木王幼树功能性状及化学计量进行方差分析,结果显示各种源间叶厚、比叶面积、叶组织密度、比叶重、叶干物质含量、叶N含量、叶C∶N、根N含量、根C∶N、C∶P均存在极显著差异,根比表面积、叶P含量、叶C∶P、根P含量存在显著差异(表2)。

表2 不同地理种源杉木王幼树功能性状及C、N、P化学计量方差分析
不同地理种源杉木种源间变异系数为7.3%—18.9%,其中比叶重变异系数最大(18.9%),其次是叶组织密度(17.9%)、叶厚(17.7%)、比叶面积(17.4%)、比根长(15.6%)、根比表面积(11.5%)、叶干物质含量(9.2%)、叶面积(7.5%),最小是根干物质含量(7.3%)。不同地理种源杉木叶厚、比叶面积、叶组织密度、比叶重、叶干物质含量、N含量、叶C∶N、根N含量、根C∶N、根C∶P差异极显著,根比表面积、叶P含量、叶C∶P、根P含量差异显著(表3)。
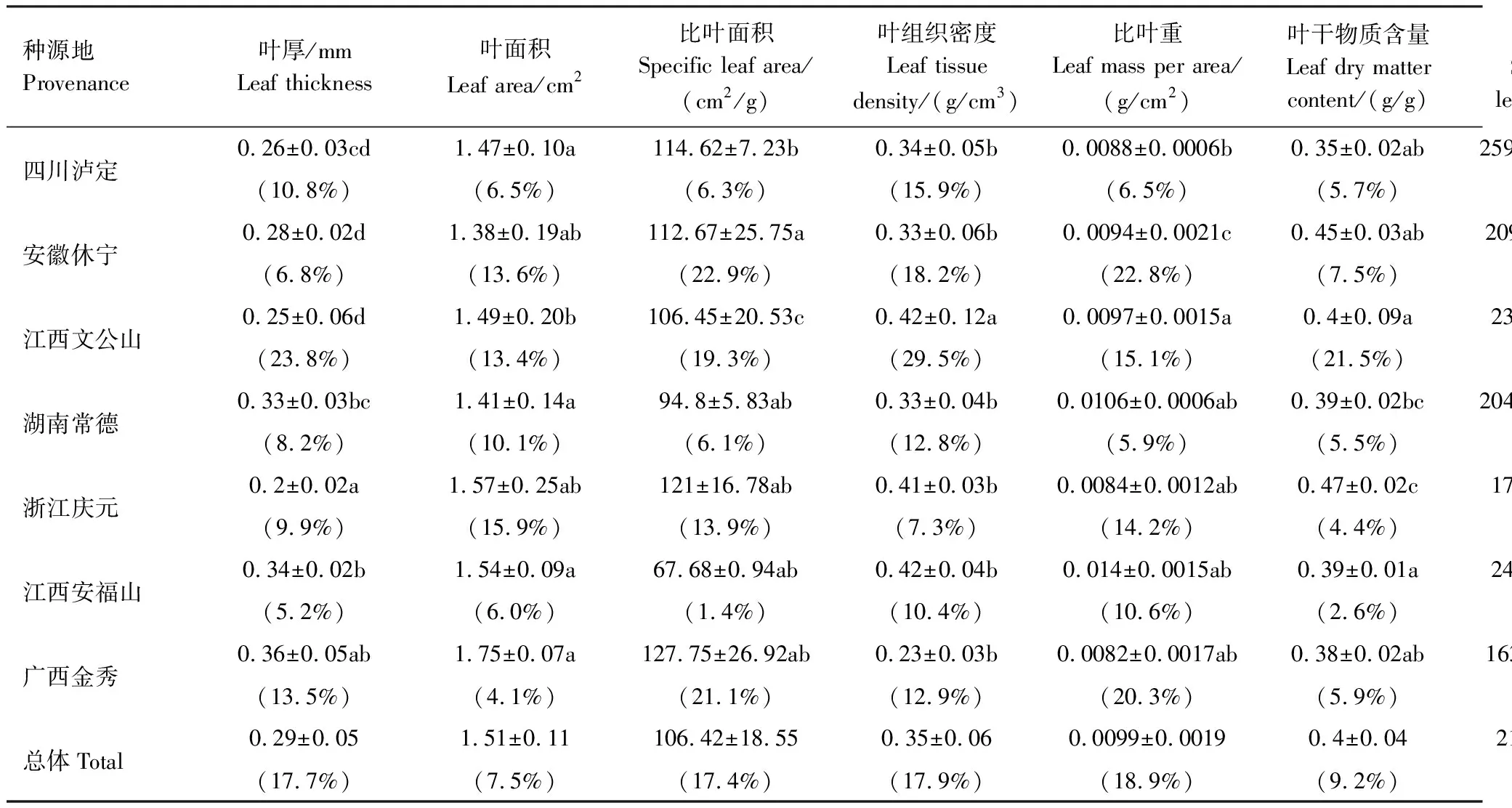
表3 不同地理种源杉木王幼树功能性状差异
对不同地理种源杉木王幼树功能性状进行分析,结果显示江西文公山的比根长、根干物质含量最小;安徽休宁叶面积、叶组织密度、比叶重、根比表面积最大、比叶面积最小;江西安福山比根长最大;浙江庆元叶干物质含量最大,叶厚、根比表面积最小;广西金秀叶厚、叶面积、比叶面积最大,叶组织密度、比叶重最小;四川泸定叶干物质含量最小(表3)。
对不同地理种源杉木王幼树C、N、P化学计量特征进行分析,结果显示种源间变异系数为1.6%—18.9%,其中根N含量最大(18.9%),其次为根C∶N(18.3%)、叶C∶P(17.3%)、根C∶P(15.9%)、叶P含量(15.5%)、根P含量(15.2%)、叶N∶P(11.4%)、根N∶P(10.2%)、叶N含量(9.8%)、叶C∶N(8.0%)、根C含量(4.3%),叶C含最小(1.6%)。江西文公山叶C含量、根N∶P最大;安徽休宁叶C含量、C∶P、根C∶N、N∶P最小,根N、P含量最高;四川泸定叶N、P、细根C∶P含量最高,叶C∶N最小;湖南常德叶C∶P最大;浙江庆元叶C∶N最大、N、P含量最低;江西安福山根C∶N最大,叶N∶P、根C、N、P含量、C∶N、C∶P最小;广西金秀根C含量最高(表4、表5)。

表4 不同地理种源杉木王幼树叶化学计量差异
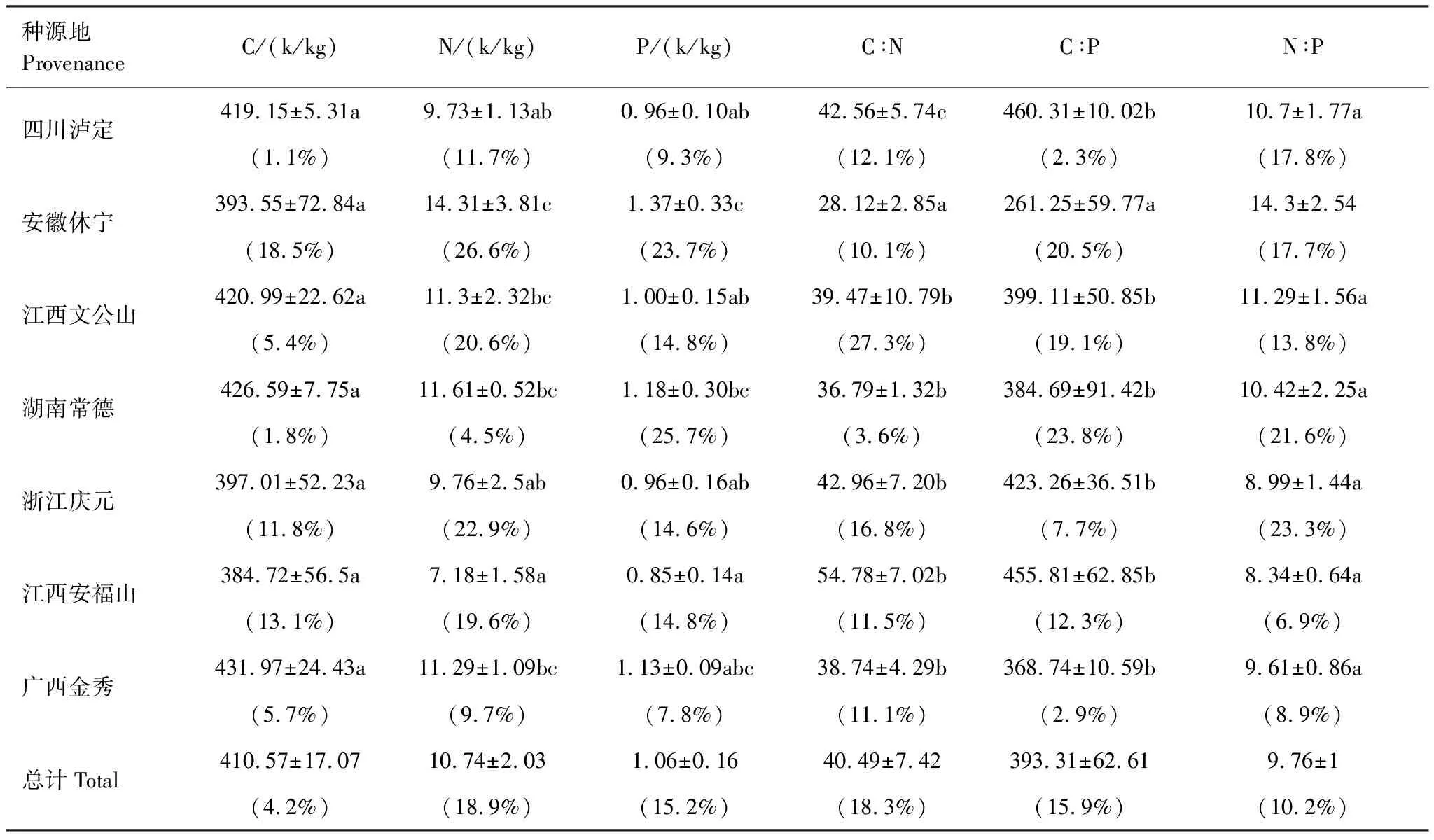
表5 不同地理种源杉木王幼树根化学计量差异
2.2 不同地理种源杉木王幼树功能性状及化学计量与地理因子冗余分析
对选取的10个种源地环境因子进行主成分(PCA)分析(表6),筛选其中单一变量解释度>10%的5环境因子,即年平均气温、海拔、经度、生长季降水量、年平均降水量,进行冗余(RDA)分析,结果显示年平均气温与根C∶P、叶C∶N、根干物质含量呈显著正相关,与根C含量呈极显著负相关;海拔与比根长呈极显著正相关性,与叶面积、叶P含量呈显著正相关,与叶干物质含量、叶N∶P呈极显著负相关;经度与叶组织密度、叶C∶P含量呈极显著正相关,与叶N含量、根比表面积呈极显著负相关;生长季降水及年平均降水量均与比叶重呈极显著正相关,叶干物质含量、根N∶P、叶N∶P呈显著正相关,与比根长呈极显著负相关(图1)。

表6 种源地环境因子对杉木王种源功能性状及化学计量解释度

图1 不同地理种源杉木王功能性状及化学计量与地理环境因子RDA分析Fig.1 Redundancy analysis functional traits andstoichiometryof C. lanceolata from different provenancesLT:叶厚 leaf thickness; LA:叶面积 leaf area; SLA:比叶面积 specific leaf area; LTD:叶组织密度Leaf tissue density; LMA:比叶重Leaf mass per area; LDMC∶叶干物质含量 leaf dry matter content; SRL:比根长 specific root length; RSA:根比表面积 root specific surface area; PDMC∶根干物质重Root dry matter content; LC∶叶C含量 leaf C correlationson; LN∶叶N含量 leaf N correlationson; LP:叶P含量 leaf P correlationson; LC∶N∶叶C∶N leaf C∶N; LC∶P:叶C∶P leaf C∶P; LN∶P:叶N∶P leaf N∶P; RC∶根C含量root C correlationson; RN:根N含量 root N correlationson; RP:根P含量 root P correlationson; RC∶N根C∶N root C∶N; RC∶P:根C∶P root C∶P; RN∶P:根N∶P root N∶P; MAT:年平均气温 Mean annual temperature; ALT 海拔 Altitude; LNG:经度 Longitude; GSP:生长季降水量 Growing season precipitation; MAP:年平均降水 Mean annual precipitation
2.3 不同地理种源杉木王幼树功能性状及化学计量相关性分析
对不同地理种源杉木王幼树功能性状和碳氮磷化学计量间相关性进行分析,结果显示叶厚与叶组织密度、叶干物质含量呈极显著负相关,与比叶面积、叶C∶N呈显著负相关,与比叶重、叶N含量呈显著正相关;叶面积与叶组织密度呈极显著负相关,与比叶重、叶干物质含量呈显著负相关,与比叶面积呈显著正相关;比叶面积与叶组织密度、比叶重呈极显著负相关,与根比表面积、根N含量呈显著负相关,与叶C∶N、根C∶N、C∶P呈显著正相关;叶组织密度与比叶重、叶干物质含量呈极显著正相关,与根C∶P呈显著负相关;比叶重与根比表面积呈极显著正相关,与根N、P含量呈显著正相关,与根C∶N、C∶P呈极显著负相关;叶干物质含量与叶N、P含量呈显著负相关,与叶C∶N、C∶P呈显著正相关;比根长与根比表面积呈极显著正相关;根比表面积与叶C含量、根C∶N、C∶P呈极显著负相关,与根N含量呈显著正相关;根干物质含量与根C含量呈显著负相关;叶片N含量与P含量呈极显著正相关、与叶C∶N、C∶P呈极显著负相关;叶片P含量与叶C∶N、C∶P、N∶P呈极显著负相关;叶C∶P与叶N∶P呈极显著正相关;根C含量与N∶P呈极显著正相关;根N含量与根C、P含量、N∶P呈极显著正相关、与根C∶N、C∶P呈极显著负相关;根P含量与根C∶N、C∶P呈极显著负相关;根C∶N与根C∶P、N∶P呈极显著正相关(表7)。

表7 不同地理种源杉木功能性状及化学计量相关性分析
3 讨论
3.1 不同地理种源杉木叶、根功能性状及化学计量差异
处于不同生态环境的同种物种,经过长期对当地生境的适应,发生地理变异,形成不同的地理种源[26],但每个物种都具有一定的适应范围,其功能性状的变异具有一定的限度[27],本研究结果显示,不同地理种源杉木功能性状种源间变异系数为7.3%—18.9%,化学计量种源间变异系数为1.6%—18.9%,均小于20%,叶厚、比叶面积、叶组织密度、比叶重等性状变异系数较大且种源间差异极显著,说明这些功能性状对环境的适应较为敏感。比叶面积表示植物单位叶片干重对光的接受和截获面积,体现植物对环境资源的利用能力和对获得资源的保存能力,与植物的同化率和生存对策密切相关[28]。以往研究表明,在资源丰富的环境中往往具有较高的比叶面积[29- 30],当植物受养分限制或种间竞争时,通过增加比根长、根比表面积提高对养分的获取能力或竞争力。本研究中,广西金秀和浙江庆元种源杉木比叶面积最大,而比根长和根比表面积均很小,表明这两地杉木受当地养分条件限制较小。C元素是植物各种生理生化过程的底物和能量来源,N、P是各种蛋白质和遗传物质的重要组成元素[31],植物C∶ N和C∶ P反映植物对N、P元素的利用效率,而N∶ P代表植物养分限制状况[32]。本研究结果显示,各种源地杉木叶C、N、P含量均大于根,除安徽休宁外,其它种源地C∶N、C∶P均为根大于叶,而叶、根N∶P均为呈现显著差异,这说明杉木根对N、P的利用率大于叶,且各种源地杉木生长均为受到养分限制。由于叶片对C、N、P利用效率低于根,所以当叶片生长造成细胞快速分裂使得叶片产生大量的养分需求,光合作用所产生的C逐渐积累,根也向叶片输送更多的N、P用于合成蛋白质和核酸,使得杉木叶片N、P含量较高[33]。不同地理种源杉木叶、根C含量未呈现显著差异,且变异系数最小。C的吸收主要是通过植物光合作用,而杉木体内的C含量保持在稳定水平,这与张婷婷等对植物生态化学计量内稳性特征的研究结果一致[34]。不同种源地杉木叶、根C∶N、C∶P及N、P含量均呈现显著或极显著差异,表明杉木叶、根N、P含量随生境变化趋势相同,以保证N∶P不变,这一结论与Pearson相关性分析一致,即叶片与细根N、P含量均呈极显著正相关。叶片和细根养分元素间的变异格局存在明显的一致性,这一结论与Zhao[35]和Liu[36]等分别就中国东部样地带和内蒙古草地植物中叶片-细根间的养分分配关系的研究结果一致。
3.2 不同地理种源杉木叶、根功能性状及化学计量相关性
叶组织密度反映了植物叶片的承载力和防御力,与叶片周转生长速度密切相关,叶组织密度的增大有利于增强植物对非生物因素的防御能力[37]。比叶重是比叶面积的反比,其大小受叶片厚度和密度的影响,比叶重越大表明单位叶面积积累的干物质越多,对贫瘠环境的耐受能力随之增强[38]。通过对不同地理种源杉木功能性状相关性分析表明,比叶重与叶厚、叶组织密度呈显著正相关,与比叶面积呈极显著负相关。因此,叶厚与叶组织密度同时增大,则叶干物质含量增大,比叶面积减小,比叶重增大,杉木对恶劣环境适应性越强。叶干物质含量与叶N、P含量呈显著负相关,与叶C∶N、C∶P呈显著正相关;比根长与根比表面积呈极显著正相关。叶片N含量与P含量呈极显著正相关、与叶C∶N、C∶P呈极显著负相关,N、P元素共同作用于杉木蛋白质与核酸的合成,故而呈极显著正相关。各种源地杉木叶片C含量差异不显著,N、P含量升高,C∶P、N∶P均下降,这与李世杰[39]对贵州东南部常见森林植物叶片研究不同,这也说明C、N、P化学计量在同种植物内与多种植物间的权衡存在差异。不同地理种源杉木叶N、P含量呈极显著相关,这与张蕾蕾[40]等对刨花楠的研究结果一致,但王楚楚[41]等对翅荚木的研究结果不同,这说明不同植物叶片对环境适应性策略也存在差异。杉木根N、P含量呈极显著正相关,这一结果与王珊[42]对不同降水条件下的红砂、珍珠的研究一致,说明不同植物根系在适应不同生境条件过程中的权衡机制相同。比叶面积与叶N、P含量未成显著相关性,比根长与根N、P含量也未呈现显著相关,与李曼[43]对武夷山森林植物的研究结果不同,但比叶面积与比根长之间不存在显著相关性的结论相同,这说明不同生境条件植物的响应机制不完全相同。徐冰[44]等的研究指出,叶片与细根性状的关系,主要受细根性状的影响,而细根性状在不同环境、生活型及功能群中均存在较大的差异,且根比表面积与比叶面积呈显著负相关,与比叶重呈极显著正相关,说明杉木根系面积生长制约叶面积生长,而根长对叶面积并无显著关联。比根长与根比表面积呈极显著正相关,根N含量与根P含量呈极显著正相关,与李淑英[45]对杉木混交林研究结果一致,这说明杉木根长、根面积生长趋势相同。
3.3 种源地环境因子对不同种源地杉木种源叶、根功能性状及化学计量的影响
根据主成分(PCA)分析筛选后可知,年平均气温、海拔、经度、生长季降水量和年平均降水量等5个环境因子对杉木功能性状及C、N、P化学计量有较大影响。连政华[46]对蒙古栎、水曲柳、山杨的研究结果表明,年平均气温与比叶面积呈显著负相关、与比叶重呈显著正相关。Reich[47]等对全球多种植物研究表明,随温度升高,植物叶N、P含量减少,N∶P增加。陈嘉静[48]等的刨花楠同质园实验表明,比叶重与纬度呈显著负相关,即随温度的升高而上升。本研究冗余(RDA)分析结果显示,不同地理种源杉木比叶面积、叶N、P含量变化趋势与上述结论一致,但显著性不明显,而比叶重未成明显趋势,这可能是由于本研究选取的7个种源地年平均气温相差不大导致的。杉木叶P含量与年平均降水及生长季降水呈极显著负相关,与叶N含量相关性不显著,但是经度影响降水,我国降水趋势从东到西,随经度减小而降低,杉木叶N含量与经度呈极显著负相关,也就说明叶N含量随降水减少而升高,这与Santiago[49]等对巴拿马森林的研究结论一致。已有结果表明,植物在适应干旱的生境得过程中为了保水或提高水分利用效率,会把更多的氮投入到叶片中[50]。植物叶片氮是光合物质代谢和植物生长的关键性因子,是合成叶绿素和有关光合蛋白的重要成分[51],磷是核酸、磷脂的组分,对植物的光合能力具有重要影响[52]。降水量增大,光照强度降低,植物为提高光合作用效率,大量合成叶绿素和光合蛋白,因而N含量升高,而P含量降低。本研究中,不同杉木种源间年平均降水量和生长季降水量与比叶面积、叶组织密度呈显著正相关,这与袁泉[53]针叶林有较低比叶面积和叶组织密度体现对干旱生境适应的结果一致。本研究中,不同杉木种源间海拔与比叶重、叶干物质含量呈极显著负相关,与叶面积呈显著正相关,与比叶面积、叶组织密度呈负相关、叶厚呈正相关。KÖrner[54]等对阿尔卑斯山植物的研究表明,随海拔升高和气温的降低,叶片厚度增加,密度减小,干物质含量减小,比叶面积减小,这一结果与本研究结论一致。但是,由于本研究选取的7个种源地年平均气温相差不大,因而海拔对气温的影响较小,所以年平均气温与比叶面积、叶组织密度、比叶重影响均不显著。
4 结论
对不同地理种源杉木根、叶功能性状的研究表明,不同地理种源杉木的叶功能形状及化学计量相较于根差异性更显著,这说明叶功能性状比根更具有可塑性,对环境的适应性变化更明显。总体上,不同地理种源间杉木根功能性状的稳定性要优于叶片。C是杉木体内最稳定的元素,杉木根对N、P的利用效率要高于叶片,且叶片和细根养分的变异格局存在一致性。对不同地理种源杉木根、叶主要功能性状的权衡关系研究表明,根比表面积与比叶面积呈显著负相关,与比叶重呈极显著正相关,比根长与根比表面积呈极显著正相关,叶片及细根N含量与P含量均呈极显著正相关,杉木叶、根面积存在关联性,根长与根面积生长趋势相同,根叶对N、P元素含量的响应也相同。对不同地理种源杉木功能性状和C、N、P计量与种源地气候环境的关系研究表明,随着温度梯度升高,不同种源杉木根C含量降低,根干物质含量升高;随着经度梯度升高,不同种源杉木叶C∶P、叶组织密度升高,叶根N含量降低;随着降水梯度升高,不同种源杉木叶根N∶P、比叶面积、比叶重升高;随着海拔梯度升高,不同种源杉木比根长、叶P含量升高、比叶重降低。
总体上,本研究初步揭示了不同地理种源杉木根、叶功能性状和C、N、P化学计量的差异、权衡关系及与种源地气候环境因子的关联。然而,受限于种源地数量和气候环境因子调查数据的不足,本研究结果仍然未能充分阐明气候环境对杉木地理分布格局的影响。将调查的种源地覆盖杉木全部栽培分布区,并补充种源地土壤营养、日照时长、坡向等微地形与微环境因子,将有助于提高研究数据的全面性和准确性。
猜你喜欢
种子(2023年7期)2023-09-24 07:40:54
特产研究(2022年6期)2023-01-17 05:05:02
天津医科大学学报(2021年1期)2021-12-05 11:11:05
辽宁林业科技(2021年1期)2021-03-16 09:17:08
农民致富之友(2020年8期)2020-05-11 05:54:38
现代园艺(2017年23期)2018-01-18 06:58:24
现代园艺(2017年23期)2018-01-18 06:58:19
中国医学影像技术(2017年11期)2017-01-16 12:54:05
考试周刊(2016年63期)2016-08-15 14:33:26
科技视界(2016年1期)2016-03-30 14:08:41
