
加工方式对黄精多糖的结构和活性影响的研究进展
2022-08-27 13:57:42吴伟菁陈家凤赵海军黄俊鹏黄宁炜吴兰兰赵海伊
食品工业科技 2022年17期
吴伟菁,陈家凤,赵海军, ,黄俊鹏,黄宁炜,吴兰兰,赵海伊
(1.厦门医学院,福建厦门 361023;2.厦门医学院,天然化妆品福建省高校工程研究中心,福建厦门 361023;3.福建省食品微生物与酶工程重点实验室,福建厦门 361018)
黄精为百合科植物,具有良好的保健功能。药食同源目录中收录的黄精有3 种,分别为黄精(Polygonatum sibiricumRed.)、滇黄精(Polygonatum kingianumColl. et Hemsl)、多花黄精(Polygonatum cyrtonemaHua)。按形状不同,习称“鸡头黄精”、“大黄精”、“姜形黄精”。黄精的活性成分包括多糖[1-4]、皂苷[5]、黄酮[1,6]、多酚[7-10]等。生黄精辛辣,其生用对人咽喉有强烈的刺激性,易引发过敏反应,且生用补性较差,故黄精需要炮制后入药或食用,以达到减毒增效和提质增效,且炮制后的黄精口感和质地均佳。但是熟黄精的活性成分发生显著变化[11]。多糖是黄精中核心功能性成分,中国药典仅对于多糖进行定量评价:生黄精多糖含量要求≥7%,炮制后的黄精多糖含量要求≥4%[12]。然而加工引起的黄精多糖含量的降低可能进一步引起多糖结构及生理活性的变化,但该问题并未引起重视,这也是黄精产业的瓶颈所在。因此,本文综述黄精常用的加工方式以及加工对三个品种黄精中多糖的含量、结构及生理活性的影响,为黄精资源在加工中的高效利用和精准控制提供参考。
1 黄精的加工方式
黄精加工的初衷是去除“刺人咽喉”的不良反应。有研究说明,与生黄精相比,加工后的黄精可显著降低溶血率,因此刺激性降低[13]。从古至今,黄精的炮制方法及工艺多样,其炮制工艺经历生用→单蒸(出于《雷公炮制论》)→重蒸(出于《千金翼方》)→九蒸九晒(出于《食疗本草》)的演变[11,14]。以“九蒸九制”为代表的反复蒸晒的方法是黄精沿用已久方法,也是现阶段黄精研究采用的最普遍的加工方式。但《中国药典》及各地的炮制规范中未收录该方法。
如表1 所示,黄精的古今炮制法存在一脉相承的相似性,即蒸制是黄精的基本炮制方法[14],单次蒸制时间一般为4~6 h。也有研究采用高压蒸制来提高效率[15-18]。另外,还可采用炖煮的方式[17,19-20]。除此之外,《中国药典》中黄精饮片经典的炮制方法将黄精与黄酒拌匀后共同蒸制,俗称酒制[12,17,20-24]。其他辅料还包括姜、黑豆、白酒、蜂蜜等[14,25]。为了保证黄精能长期保存,传统黄精加工通过晒制来降低水分活度。但现代加工常采用烘箱烘干的方式。
目前黄精的加工工艺研究未成体系,但其原因是无可控的炮制工艺参数。另外,黄精炮制终点的判断依靠主观经验判断。例如,药典中黄精炮制终点描述为炖透或蒸透[12],指标过于宽泛。因此,通过深入研究加工过程中黄精活性成分变化规律从而规范其工艺参数显得尤为必要。
2 不同加工方式对黄精多糖含量的影响
目前加工方式对黄精中多糖含量的影响的研究多基于“九蒸九制”的方法。基于不同方式(清蒸、炖煮、酒蒸等)的反复处理均能引起黄精中多糖含量的急剧降低。多数研究中,熟黄精中多糖含量均降低近一半以上(表1)。其次,加以辅料对多糖的下降无影响,清蒸和酒制均可引起多糖含量随着蒸制次数增加而下降[22],并且最终多糖含量相近(清蒸:5.5%~6.9%,酒蒸:5.6%~6.5%)[24]。三类黄精中的多糖显著下降多发生在前三蒸,随后多糖含量稳定。蒸制引起多糖含量下降可能与时间有关而非次数。以滇黄精为例,多糖下降发生在前48 h,随后增加时间对多糖含量无影响[26]。因此,减少蒸制时间可有效减少多糖损失[27]。炮制引起的多糖损失与传统认为的炮制补益且增强药效存在差距[14]。相对于蒸制,黄精加工耗时最长的干燥对多糖的影响研究较少。如表1 所示,黄精干燥通常采用热风干燥,温度为45~70 ℃,烘干时间大多≥12 h。也有研究采用105 ℃烘干黄精[26],但黄精多糖在高温烘干中是否受到影响值得进一步研究。相比于热风干燥,微波真空干燥更有利于保存多花黄精中多糖含量[28-29],说明烘干的温度对于黄精多糖的保留也很重要。
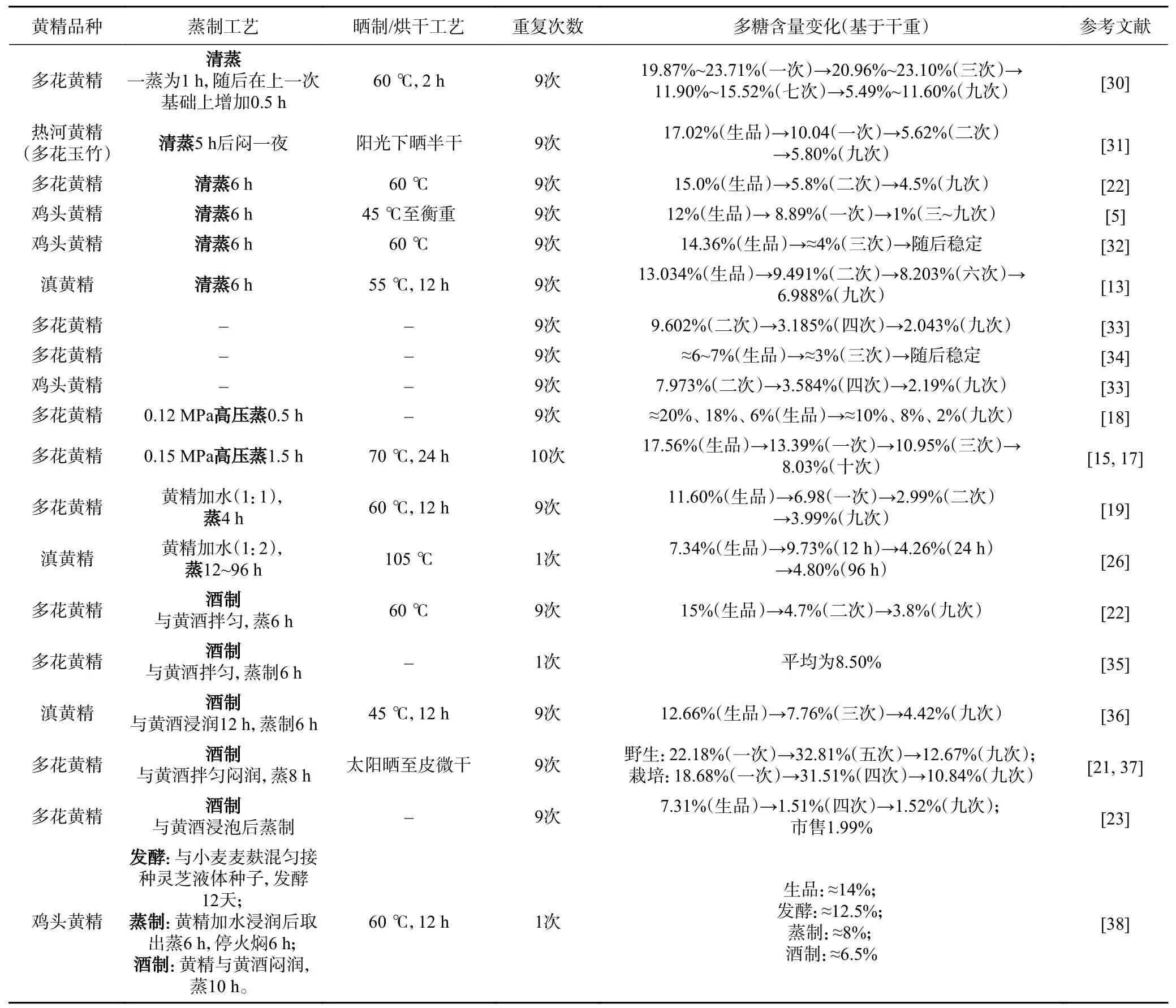
表1 不同加工方式对黄精中多糖含量的影响Table 1 Effects of different processing methods on polysaccharides content in Polygonatum spp.
综上,不同加工方式均能引起黄精中多糖含量显著下降,而未来的研究可以多糖含量为质量控制标准,明晰引起多糖下降的关键控制因素(温度、水分、时间等)。同时,以水分活度为标准,控制烘干条件(方式、温度、时间),以提高黄精的加工效率。
3 黄精多糖结构及其在加工中的变化
三类黄精(生品)多糖的结构有所不同(表2)。生多花黄精多糖以中性多糖为主的果聚糖[3,39-45]。也有报道多花黄精多糖的单糖组成以葡萄糖为主[45],这与单糖检测方法有关。果糖在检测前处理(强酸水解)下,可以转变为甘露糖及葡萄糖,因此果糖无法被识别[46]。生鸡头黄精多糖是以半乳糖、甘露糖为主形成的中性多糖[22,47-54]。生滇黄精多糖是以葡萄糖为主形成的中性多糖[2]。三种不同来源黄精多糖的单糖组成有较大不同,因而三种黄精多糖指纹图谱上存在差异[36,55],可能对其活性功能有一定的影响。
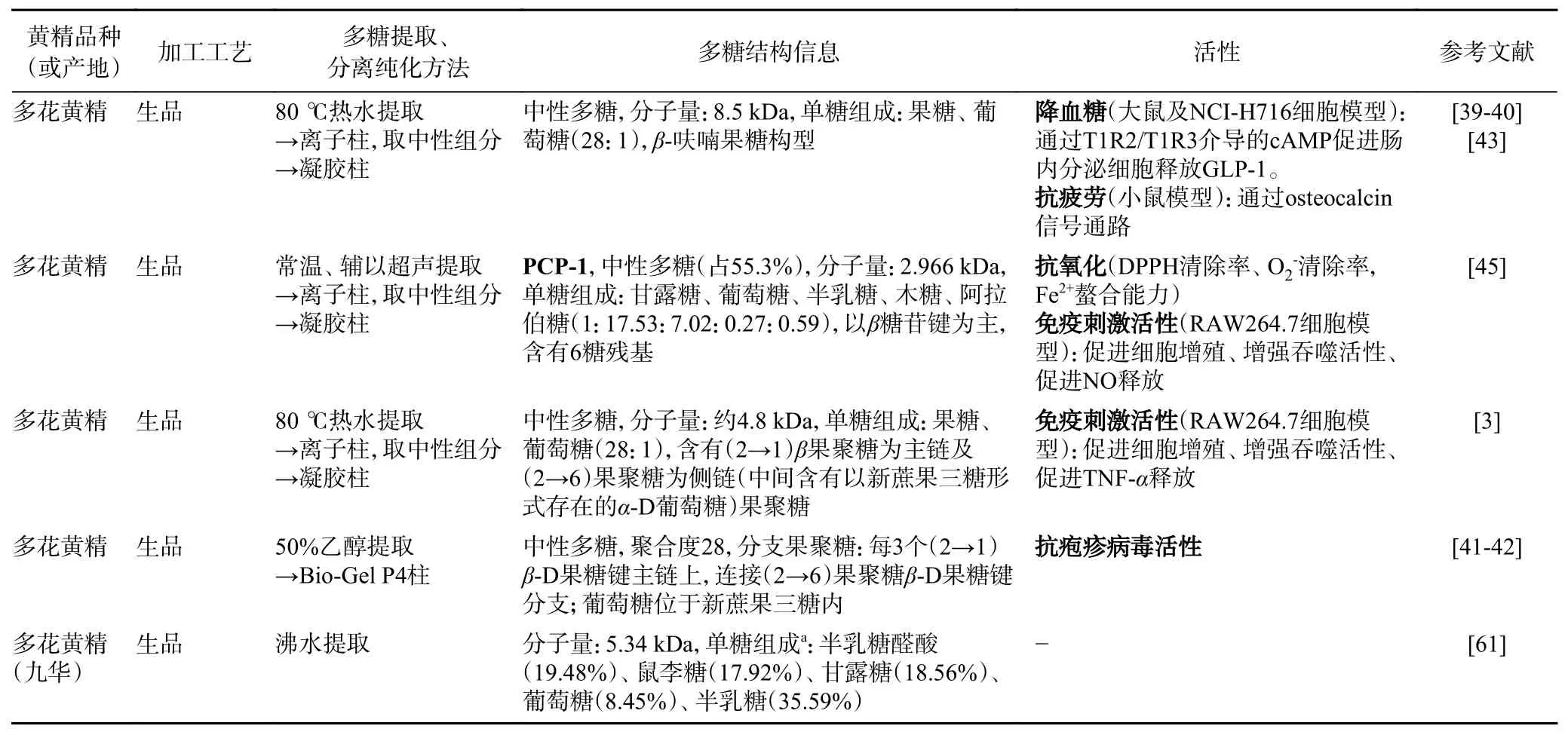
表2 黄精多糖结构、活性及其在加工中的变化Table 2 Structure and bioactivity of polysaccharides from Polygonatum spp. and their changes during processing
熟黄精不仅多糖含量显著降低,而还原糖、总体单糖含量显著上升[23,26,56],说明黄精多糖发生降解。有研究发现(如图1 所示),在反复蒸制过程中,多花黄精多糖可先降解为寡聚糖,随后进一步降解成单糖[18]。与此同时,黄精多糖中单糖组成结构上发生变化。首先,蒸制使得三类黄精的多糖从以中性多糖为主转为含有酸性多糖。熟黄精多糖中半乳糖醛酸摩尔质量比增加至6.1%~41.4%[44,48,57-58]。其次,黄精多糖的单糖组成在蒸制中发生变化,生多花黄精多糖以果糖为主,占96.56%mol[39-40],但熟多花黄精多糖中果糖摩尔质量占比降低至24.6%,半乳糖醛酸及半乳糖的摩尔质量比也分别增加至20.5%及41.4%[44]。鸡头黄精多糖随着蒸制次数增加,甘露糖含量不断下降(52.47%→18.86%),葡萄糖含量先降低后增加(36.84%→32.17%→52.20%),半乳糖及阿拉伯糖含量均增加(7.32%→21.77%及3.37%→7.58%)[32]。也有研究说明鸡头黄精多糖随着蒸制次数增加,甘露糖的摩尔质量占比不断下降(51.2%→17.5%),葡萄糖摩尔质量占比先增加后降低(21.4%→65.5%→46.5%),半乳糖的摩尔质量占比先降低后增加(26.9%→8.97%→30.1%),阿拉伯糖的摩尔质量占比在蒸制过程中有增有减,但熟鸡头黄精多糖中阿拉伯糖摩尔质量比例显著高于生鸡头黄精多糖(0.46%(生黄精),5.89%(九蒸九制))[54]。
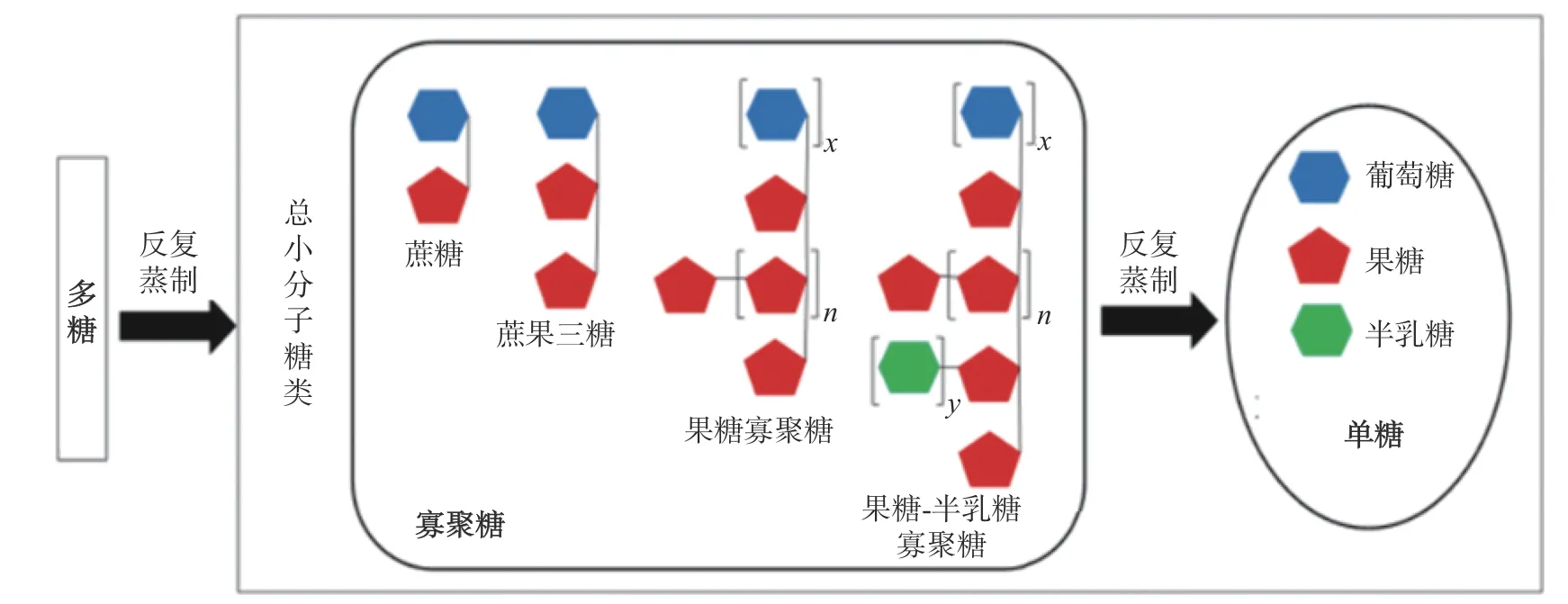
图1 多花黄精多糖在“九蒸九制”过程中的变化[18]Fig.1 Changes of polysaccharides from Polygonatum cyrtonema Hua processed by different times of repeated steaming [18]
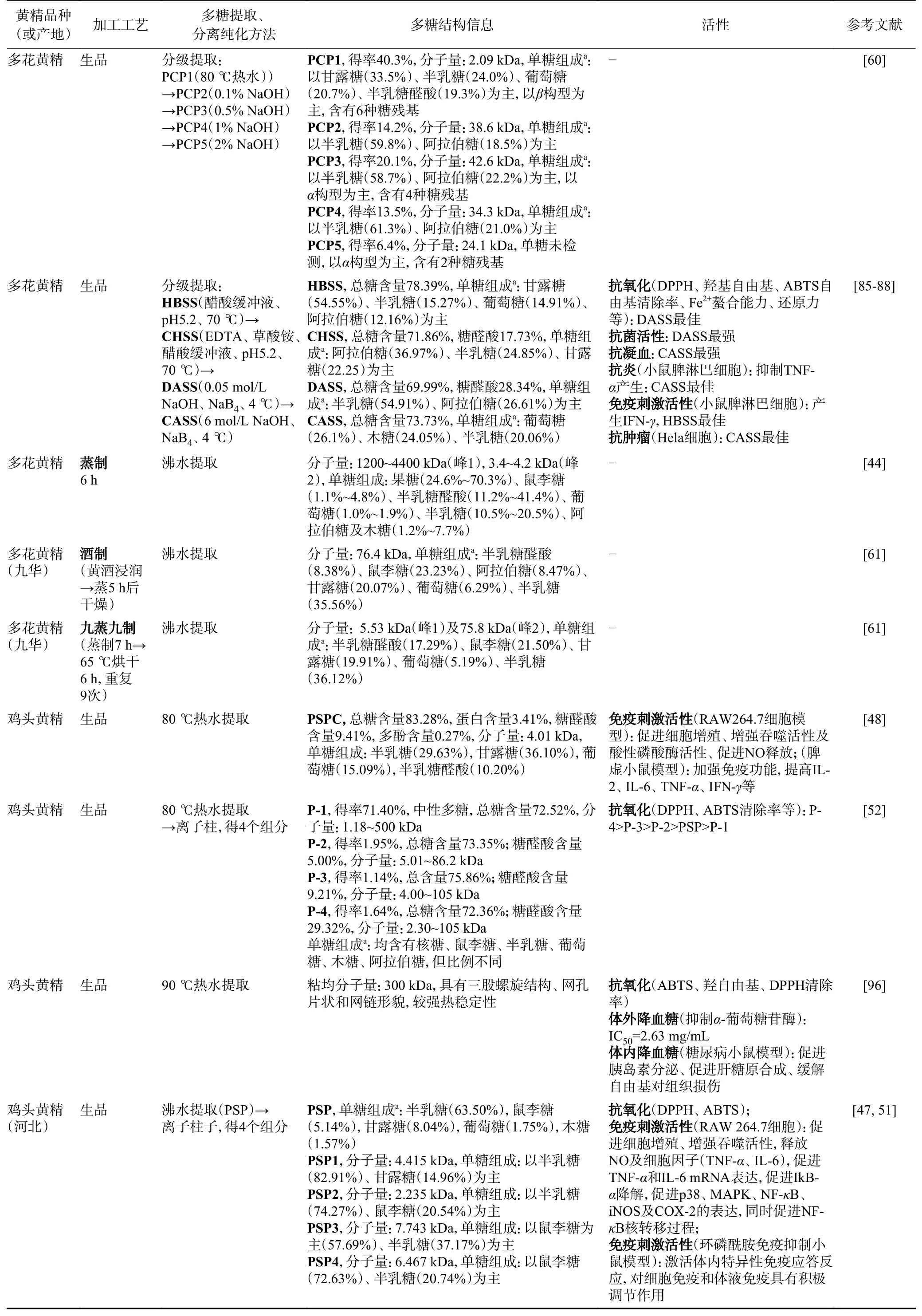
续表 2
但是,黄精多糖蒸制过程中降解的同时还发生了聚集。蒸制可导致三种黄精多糖分子量显著增加[36,44,48,59]。生多花黄精多糖的分子量为2.09~8.5 kDa[3,39-40,45,60-61]。而蒸制6 h 后的多花黄精多糖中除了低分子量的组分(3.4~4.0 kDa)之外,还含有高分子量组分(1200~4400 kDa)[44]。另外,“九蒸九制”的多花黄精的多糖有2 个片段,分别为5.53 kDa及75.8 kDa,酒黄精多糖中有一个高分子片段76.4 kDa[61]。蒸制后的鸡头黄精分子量增加[44,48,54]。例如,酒蒸6 h 后的鸡头黄精多糖的分子量也从4.01 kDa 增加至14.2 kDa[48]。也有研究发现在“九蒸九制”过程中,鸡头黄精分子量从6.06 kDa 先增至10.3 kDa(一蒸),随后降至4.63~5.10 kDa(二~四蒸),随后分子量显著增至44.3~75.3 kDa(五蒸~九蒸)[54]。与此同时,通过PCA 分类可得出,鸡头黄精(生品)多糖、一至四蒸的鸡头黄精多糖、五至九蒸的鸡头黄精多糖分别为三个品类的多糖[54]。滇黄精(生品)多糖分子量为14.05 kDa[2],而滇黄精(饮片)多糖分子量为149.4 kDa[58]。有研究说明,滇黄精(生品)中多糖及八至九蒸的熟黄精中多糖分别为两类多糖,而中间一至七蒸的熟黄精多糖差异较大,较为分散[36]。黄精多糖在蒸制过程分子量增加可能是由于多糖分子含有大量的羟基,由于分子间相互作用力,尤其是酸性多糖容易形成不同程度的聚集体,导致多糖分子量增加。
综上,由于三种生黄精多糖结构上有较大区别,因而在研究中需要区别不同黄精品种。进一步,在加工过程中,三种熟黄精中多糖的结构与生黄精中多糖具有显著差异。但目前仍然缺乏对加工过程黄精多糖的结构动态变化规律的系统解析,例如糖苷键断裂的方式,多糖链组成及高级溶液构象等。
4 黄精多糖活性及其在加工中的变化
多糖的结构与功能有密切关系,黄精多糖在加工过程中结构的变化会影响其活性,如表2 所示。
4.1 抗氧化活性
研究发现,三类的生黄精多糖均具有抗氧化活性[45,52,57],但与通用的阳性对照维生素C 相比,黄精多糖抗氧化活性较低[53,62]。鸡头黄精蒸制后,其多糖体外抗氧化活性随着蒸制次数的增加呈现增加的趋势[32,54],尤其在第四蒸后显著增加[54]。“九蒸九制”过程中的鸡头黄精多糖的抗氧化活性与其单糖组成相关,即鸡头黄精多糖的ABTS 自由基清除及羟自由基清除率与甘露糖摩尔质量占比成反比,与阿拉伯糖摩尔质量占比成正比,ABTS 自由基清除率与葡萄糖摩尔质量占比成正比[54]。不同干燥方法可改变黄精多糖的抗氧化活性,微波真空干燥法获得的黄精的抗氧化活性显著高于热风干燥[28]。经过不同处理(发酵、蒸、酒制)后的水提液体,发酵和蒸制后的样品的DPPH 自由基清除率较生黄精样品均提高,发酵和酒制后的样品铁离子还原能力显著提高[38]。由于该研究采用水提液,而水提液中的其他物质也可能引起抗氧化活性的变化。另外,体外细胞研究表明黄精多糖能通过增强细胞内抗氧化活性,提高HT22 细胞的存活率,改善线粒体功能,从而保护细胞因H2O2引起的氧化损伤,其作用机制与激活SIRT1/AMPK/PGC-1α信号通路有关[63]。
4.2 降血糖
三类生黄精多糖均具有降血糖功效。市售鸡头黄精多糖不仅可降低STZ 诱导糖尿病大鼠的血糖,而且可限制视网膜病理性血管生成[64]。通过下调信号通路(Bax、EGF、p38、VEGF 和TGF-β)及上调Bcl-2 来抑制视网膜组织中细胞凋亡,进而减缓糖尿病引起的视网膜受损[64]。滇黄精多糖可以降低二型糖尿病大鼠的空腹血糖,调节肠道菌群,促进短链脂肪酸产生[65]。鸡头黄精多糖在体外模拟酵解过程中能够显著提高来源于二型糖尿病小鼠的肠道菌群的丰富度和多样性[66]。多花黄精多糖能够提高一型糖尿病大鼠生存率、降低空腹血糖,具有肝保护作用(降低炎症因子)[67]。同时,多花黄精多糖能促进GLP-1生成,而GLP-1 与胰岛功能、饮食控制、血糖稳态、炎症及心血管保护作用有关[39-40]。王艳芳[68]研究发现,高压蒸制结合酒制后的滇黄精多糖也具有降低高脂小鼠血糖、空腹糖耐量、血清胰岛素及胰岛素抵抗的作用。总之,加工前后的黄精多糖均具有降血糖作用,其降血糖机制主要通过调节肠道菌群及改善胰岛素功能。但是,目前尚缺乏生、熟黄精多糖之间的降血糖效果及机制差异上比较的研究。
4.3 改善脂肪代谢异常
生鸡头黄精多糖可通过AMPK 通路,调节脂肪代谢及炎症反应从而改善高脂诱导的小鼠肥胖[69]。滇黄精多糖也可改善脂肪代谢异常[68,70-72]。生品及酒制滇黄精多糖均能通过改善线粒体功能而缓解高脂饮食诱导的非酒精性脂肪肝,其机制可能与增强线粒体的抗氧化应激、能量代谢、脂肪酸β-氧化以及抑制肝细胞凋亡有关。但酒制的滇黄精多糖在降低血清与肝脏脂质,提高线粒体能量代谢及抑制线粒体介导的肝细胞凋亡方面效果优于生滇黄精多糖,而生滇黄精多糖抗氧化应激效果优于加工后的滇黄精多糖的效果[71]。因而,生、熟黄精的多糖具有改善脂肪代谢异常的作用,二者在降血脂的机制上各有优势。
4.4 增强免疫力
生多花黄精多糖[3]、生、熟鸡头黄精多糖[47,49,73]在体内、体外实验中具有显著的提高免疫力的活性,但滇黄精的多糖研究较少。鸡头黄精(生品)的多糖可以通过多条通路(MR 及TLR4[49]、NF-κB/MAPK[47])刺激RAW264.7 细胞的免疫活性,也可通过CR3 及TRL2 通路刺激NK 细胞活性[49]。另外,黄精(湖北)多糖可以提高环磷酰胺免疫抑制小鼠模型的脾脏指数、胸腺指数、促进溶血素形成、提高腹腔巨噬细胞吞噬功能[74]。黄精(延边)多糖可以促进小鼠脾淋巴细胞产生IL-2 及IFN-γ;促进RAW264.7 巨噬细胞增殖、产生IL-6(白细胞介素-6,Interleukin-6)及TNF-α(肿瘤坏死因子-α,Tumor necrosis factor-α)、NO(一氧化氮,Nitric oxide)、提高iNOS mRNA 水平,从而提高免疫活性[75]。
加工可改变黄精多糖的免疫活性。研究表明,基于RAW264.7 细胞模型及动物模型,酒制鸡头黄精多糖的免疫刺激活性优于生黄精多糖[48]。另外有研究表明,鸡头黄精(生品)多糖及鸡头黄精(酒制)多糖均具有免疫调节作用,分子量<50 kDa 的组分是多糖的有效部位,且酒制后的黄精多糖抑制炎症反应作用加强[27]。因此,虽然研究已报道酒制后黄精多糖具有比生黄精多糖更高的免疫刺激能力[48],但是由于酒制黄精加入了黄酒,黄酒的作用值得单独探究。不同炮制方式(四制及九制)黄精水提物对气阴两虚大鼠模型具有不同优势,四制黄精水提物调节血脂代谢改善体重方面最佳,而九制黄精水提物在改善肝功能,提高免疫球蛋白效果较为突出[23]。然而该研究采用水提物中可能还有其他的活性组分,需进一步明晰其多糖的实际贡献。综上,黄精中核心的多糖的免疫刺激活性在加工中的变化值得进一步明晰。
4.5 抗疲劳
三类生黄精多糖在动物模型上均具有显著的抗疲劳效果,包括延长小鼠负重游泳时间、降低运动后的血清尿素氮、提高肝糖原[20,76-80]。其中,多花黄精多糖还能显著降低负重游泳后的血乳酸含量[76]。但是,三种黄精(生品)多糖在抗疲劳效果无显著差异。多花黄精的中性多糖组分PCP-1 的抗疲劳机制是通过osteocalcin 信号通路[43]。通过超高压(293 MPa)提取的鸡头黄精(生品)多糖通过提高机体代谢与抗氧化能力,减少腓肠肌线粒体应激损伤实现抗疲劳功效[81]。
通过不同炮制方法(纹制、“九蒸九晒”、酒制)获得的黄精粗多糖能够提高小鼠负重游泳时间[82-84]。生黄精粗多糖比炮制后的样品能显著提高SOD 及降低MDA[82]。但也有研究表明多花黄精炮制品中多糖优于生黄精中多糖,包括抗疲劳效果及组织中SOD 活力的提高,MDA 含量的降低,从而抗氧化程度增加[84]。但该研究采用水煎液体,未对多糖进行进一步纯化分离,水提液中存在较多的其他物质,例如,多酚等可能对结果产生干扰。有研究表明,古法炮制后的黄精具有良好的抗疲劳效果[20],但该研究未与生黄精进行比较,因此炮制方法对于黄精多糖的影响尚未明确。虽然研究表明生、熟黄精多糖在同等剂量下均具有抗疲劳效果[82-83]。但是,由于加工降低黄精多糖含量,多糖的降低对于其实际抗疲劳效果可能产生不利影响。
4.6 抑菌活性
采用分级提取的生多花黄精获得的不同多糖均具有抑菌活性[85-88]。加工对于黄精的抑菌活性有一定的改变[16]。例如,高压2 h 获得熟黄精多糖对大肠杆菌、金黄色葡萄球菌的抑制效果较好,生黄精多糖则对枯草芽孢杆菌的抑制作用相对较强[16]。
4.7 其他活性
除上所述,黄精多糖还拥有多种生理活性,其中鸡头黄精多糖的研究较多。鸡头黄精多糖,通过增强免疫力(TLR4-MAPK/NF-κB 信号通路)从而抗肺肿瘤[73]。多花黄精粗多糖能够体外抑制S180癌细胞及人乳腺癌细胞MCF-7,在体内具有抑制S180肉瘤的作用,其机制也是通过增强免疫力[89]。鸡头黄精多糖还可以提高D-半乳糖诱导衰老大鼠的学习和记忆能力,改善其肾脏组织的病变,通过Klotho-FGF23 内分泌轴,改善抗氧化,平衡钙、磷代谢[90]。黄精多糖还具有提高阿尔兹海默病症模型的学习记忆能力[91-92]。鸡头黄精多糖能够通过抑制NGAL 或者KIM-1 mRNA的表达,抑制p38MAPK/ATF2 信号通路从而抑制TNF-α、IL-1β及IL-6 进而修复庆大霉素诱导的急性肾损伤[93]。鸡头黄精多糖对椎间盘退变大鼠髓核细胞凋亡、炎症和氧化应激的保护作用[94]。鸡头黄精多糖还可以通过促进VEGF、骨钙素表达抑制Col1a1、ACP5、CTSK 蛋白表达,促进大鼠胫骨骨折愈合[95]。
综上,目前生、熟黄精多糖在不同的生物活性上各有优势。生黄精的活性研究较为充分,需要加强对熟黄精多糖的活性及机制研究,这对于黄精功能食品的开发利用尤为重要。进一步,系统比较加工对黄精多糖的生物活性的影响对于黄精加工的精准控制具有重要作用。
5 展望
综上,基于文献检索,本文对黄精常用的加工方式进行综述,分析目前加工存在的问题。进一步阐述加工方式对黄精中多糖含量、结构及活性的影响。已有研究已明确黄精在加工过程中发生多糖含量显著下降,并且黄精多糖结构降解同时发生聚集,但引起多糖含量及结构变化的关键控制点尚不明确。现有研究发现加工可一定程度改变黄精多糖的生物活性,因此不可忽视加工对于黄精多糖构效关系的影响,但目前尚未开展深入研究。因此,解析加工过程中黄精多糖含量及多糖结构和活性的变化可帮助控制过度加工引起的黄精多糖含量的下降等不良影响。这些拓展研究将为黄精食品生产中精准控制及开发利用提供重要理论基础和科学依据,对黄精产业规范化加工及其健康持续发展具有重要意义。
猜你喜欢
小猕猴学习画刊·下半月(2023年10期)2023-11-04 10:08:36
课外生活·趣知识(2023年8期)2023-10-08 18:09:15
Digital Chinese Medicine(2020年3期)2020-11-03 03:12:12
云南农业科技(2020年5期)2020-10-14 07:24:50
中成药(2018年12期)2018-12-29 12:25:50
中成药(2018年9期)2018-10-09 07:18:58
中成药(2018年9期)2018-10-09 07:18:38
西夏研究(2017年4期)2017-08-17 08:53:12
西藏科技(2016年5期)2016-09-26 12:16:40
药学与临床研究(2015年4期)2015-06-05 11:35:51
