
二脒那秦内源性激活血管紧张素转化酶2对细胞脂质沉积的抑制效应及其机制
2022-08-26 09:00曹西月付亚丽谢娜娜张亚峰张源淑
畜牧兽医学报 2022年8期
曹西月,付亚丽,谢娜娜,张亚峰,张源淑
(南京农业大学动物医学院/农业部动物生理生化重点开放实验室,南京 210095)
脂肪组织是机体储存多余脂质的主要场所,同时也是机体的一个重要的内分泌器官,可影响机体糖脂代谢、胰岛素敏感性等,参与机体多种生理病理过程。脂质在脂肪组织的过度沉积会直接引起肥胖,也大大增加了患高血脂、高血压、糖尿病等代谢相关疾病的风险,已成为严重影响公共卫生健康的主要社会问题之一。在畜牧业中,动物脂肪的过度沉积不仅降低了饲料利用率,直接影响动物产品的品质,而且会影响动物的繁殖能力。因此,寻找改善动物机体脂质过度沉积的药物或调节剂成为畜牧工作者共同关心的问题。
肾素-血管紧张素系统(renin-angiotensin system,RAS)在调节机体体液和电解质平衡以及心血管稳态方面至关重要,主要包括ACE/Ang Ⅱ/AT1R经典轴和ACE2/Ang1-7/Mas调节轴两条调节通路,后者在一定程度上拮抗前者。近年来,越来越多的研究表明,RAS两条调节通路参与了体内的糖脂代谢、胰岛素抵抗及糖尿病并发症等多种过程。并证实经典轴上的关键肽分子Ang II的过度活跃,会加重不同器官和系统的代谢紊乱,引起胰岛素抵抗,而ACE2/Ang1-7/Mas轴的激活则能够促进啮齿动物能量消耗,抑制Ang II过多导致的肥胖或脂肪沉积。RAS系统中的主要成员血管紧张素转化酶2(Angiotensin converting enzyme 2, ACE2)可以通过将Ang II水解成Ang 1-7减少脂肪组织炎症,改善葡萄糖稳态,被认为是治疗肥胖或脂肪沉积的潜在靶点。
二脒那秦(diminazene aceturate,DIZE)是一种常见的兽用抗寄生虫药物,2011年发现其可内源性激活ACE2,给小鼠口服可降低体重,减少白色脂肪组织沉积,并改善小鼠的代谢状况。随后有研究发现 DIZE可通过激活ACE2改善糖尿病鼠的心脏功能障碍和视网膜病变,提示DIZE作为ACE2的内源性激活剂。显示出巨大的研究或开发潜力。但目前对DIZE的应用研究多局限于糖尿病肾病、心血管疾病方面,其在脂肪组织脂肪沉积及代谢紊乱中的作用及机制的研究尚未见报道。
本试验以小鼠前脂肪细胞(3T3-L1)为对象,通过诱导分化获得脂肪细胞,确定DIZE与脂肪沉积及ACE2的相互关系,再用siACE2处理细胞抑制ACE2的表达,探讨DIZE介导ACE2的内源性激活对3T3-L1前脂肪细胞脂质沉积的影响,并从脂质合成及葡萄糖摄取和氧化代谢方面探讨其相关机制,为从ACE2的角度改善脂肪沉积的开发研究提供理论依据。
1 材料与方法
1.1 细胞株
3T3-L1小鼠前脂肪细胞由南京医科大学基础医学院生化研究室惠赠。
1.2 试剂与仪器
DIZE购自美国SIGMA公司;3-异丁基-1-甲基黄嘌呤(3-Isobutyl-1-methylxanthine,IBMX)和地塞米松(Dexamethasone,DEX)购自上海陶术生物科技有限公司;重组人胰岛素购自南京凯默尔生物科技中心;Opti-MEM培养基购自美国Gibco公司;Lipofectamine 3000 购自赛默飞世尔科技有限公司;无毒快速油红O染液、葡萄糖和三酰甘油定量测定试剂盒均购自南京建成生物工程研究所;4% 多聚甲醛购自南京寿德实验器材有限公司;RIPA蛋白裂解液购自南京碧云天生物技术有限公司;PVDF膜购自美国Millipore公司;ECL化学发光检测试剂盒购自上海天能科技有限公司等。
兔源β-actin抗体(货号AC026)、辣根过氧化物酶标记的山羊抗兔IgG二抗(货号AS014)均购自武汉爱博泰克生物科技有限公司;兔源柠檬酸合酶抗体(CS,货号GB111883)和兔源脂肪酸合酶抗体(FAS,货号GB11546)购自武汉赛维尔生物科技有限公司;兔源葡萄糖转运蛋白 4 型抗体(GLUT4,货号BS3680)和ACE2抗体(货号BS66001)均购自南京巴傲得生物科技有限公司;兔源乙酰 CoA 羧化酶抗体(ACC,货号D155300)和固醇调节元件结合蛋白-1c 抗体(SREBP1c,货号NB100)购自生工生物工程(上海)股份有限公司。
Tecan Spark多功能酶标仪(瑞士,Tecan公司);POWER-PAC300电泳仪(美国,Bio-Rad公司);POWER-PAC HC转印仪(美国,Bio-Rad公司);Tanon-3900全自动化学发光图像分析系统(上海天能科技有限公司)等。
1.3 细胞培养与处理
1.3.1 细胞诱导分化 参照张许等方法。3T3-L1前脂肪细胞接种于6孔板内,常规DMEM高糖培养基,10% 胎牛血清,37 ℃、5% CO条件下培养,待细胞生长至完全融合,加入分化诱导剂 Ⅰ(10% FBS, 0.5 mmol·LIBMX,10 μg·mL胰岛素和 2.0 mmol·LDEX 的诱导分化剂),继续培养,待80% 左右的细胞内出现脂滴后,换分化诱导剂 Ⅱ(10% FBS,10 μg·mL胰岛素),48 h后弃诱导剂 Ⅱ,改用常规培养液,直至95% 以上细胞出现脂滴。
1.3.2 DIZE处理及细胞活力检测 首先用不同浓度DIZE(0、6.25、12.5、25、50和100 μmol·L)处理分化成功的3T3-L1脂肪细胞24、48、72 h,常规MTT法测定细胞活力,于490 nm处测定吸光度。筛选DIZE作用合适浓度和时间。按如下公式计算:
细胞活力值(%)=(样品孔A490-空白孔A490)/(对照孔A490-空白孔A490)×100。
确定DIZE作用浓度为12.5和25 μmol·L,处理时间为48 h,用于后续试验。
1.3.3 siACE2转染干扰细胞ACE2表达 参照本实验室刘颖建立的方法。以3×10个·孔的密度将细胞接种于6孔板中,诱导分化至倒数第3天;用200 μL Opti-MEM稀释适量siRNA(siRNA序列见表1),混匀,室温静置5 min;用200 μL Opti-MEM加7.5 μL Lipofectamine 3000,混匀,室温5 min; 将上述两步的溶液混合,室温孵育20 min;无血清培养基漂洗细胞,然后在每孔中加入常规培养液1.5 mL;每孔加入上述混合溶液400 μL;将细胞置于培养箱中孵育6 h,继续培养48 h。
1.3.4 试验分组及处理 将诱导分化成功的3T3-L1脂肪细胞进行分组:1)对照组(control group),细胞培养基为常规培养液。2)DIZE试验组(12.5和25 μmol·LDIZE group),其中,一组为分化诱导剂 Ⅱ +12.5 μmol·LDIZE溶液,一组为分化诱导剂 Ⅱ +25 μmol·LDIZE溶液。3)siACE2 组(siACE2 group),即siACE2 干扰处理诱导分化成功的3T3-L1脂肪细胞。4)siACE2+DIZE组(siACE2+DIZE group),即siACE2 干扰预处理诱导分化成功的3T3-L1脂肪细胞,加25 μmol·LDIZE。以上分组处理完毕后,37 ℃、5% CO继续培养48 h。

表1 siRNA片段序列
1.4 指标测定
1.4.1 细胞上清中三酰甘油和葡萄糖含量测定 三酰甘油采用GPO-PAP法,操作按试剂盒说明书方法进行。510 nm处空白孔调零,测各孔OD值,按如下公式计算含量:
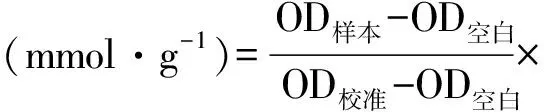
葡萄糖测定采用葡萄糖氧化酶法,按试剂盒说明书方法进行。空白孔调零,505 nm测各孔OD值,按如下公式计算含量:
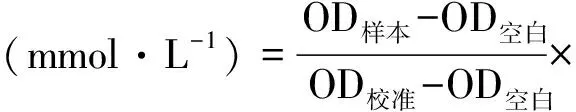
葡萄糖吸收量(mmol·L)=培养液葡萄糖(mmol·L)-处理组葡萄糖(mmol·L)
1.4.2 细胞油红O染色 脂滴染色操作参照无毒快速油红O染液试剂盒说明书。在6孔板中处理细胞,4% 多聚甲醛固定,油红O染液,洗涤。复染液染色,洗涤,镜下观察并拍照。弃PBS,100% 异丙醇回收油红,测每孔OD值,进行定量分析。
1.4.3 细胞ACE2、FAS、ACC、SREBP-1c及GLUT4和CS的蛋白检测 1)蛋白裂解液制备:RIPA裂解(含1% PMSF)获得总蛋白提取物。BCA法测定蛋白浓度,统一蛋白浓度,-20 ℃储存备用。2)SDS-PAGE:采用10% 分离胶和5% 浓缩胶的聚丙烯酰胺凝胶,蛋白上样量为30 μg,蛋白Marker 2 μL;电泳条件:浓缩胶80 V,分离胶110 V。3)转印:湿法转印,选用PVDF膜,90 V,转膜90 min。 4)杂交和显色:转膜结束后,取出PVDF膜,5% 脱脂奶粉封闭2 h,TBST洗膜。分别加入一抗(β-actin兔源抗体1∶10 000稀释;ACE2、FAS、ACC、SREBP-1c、GLUT4及CS兔源抗体1∶1 000稀释),4 ℃孵育过夜。TBST洗膜,加入山羊抗兔IgG二抗(1∶10 000稀释),室温孵育,最后洗膜。ECL发光(A液∶B液=1∶1),凝胶成像系统曝光并拍照。5)数据处理:Image J软件检测各蛋白条带灰度值,以β-actin作内参,目的条带与其相比得到目的蛋白的相对表达量,然后对数值进行归一化处理。
1.5 数据统计与分析
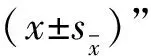
2 结 果
2.1 细胞转分化结果
由图1可见,至诱导分化的第14天,95% 细胞出现脂滴,说明前脂肪细胞诱导成功,可用于后续的研究。
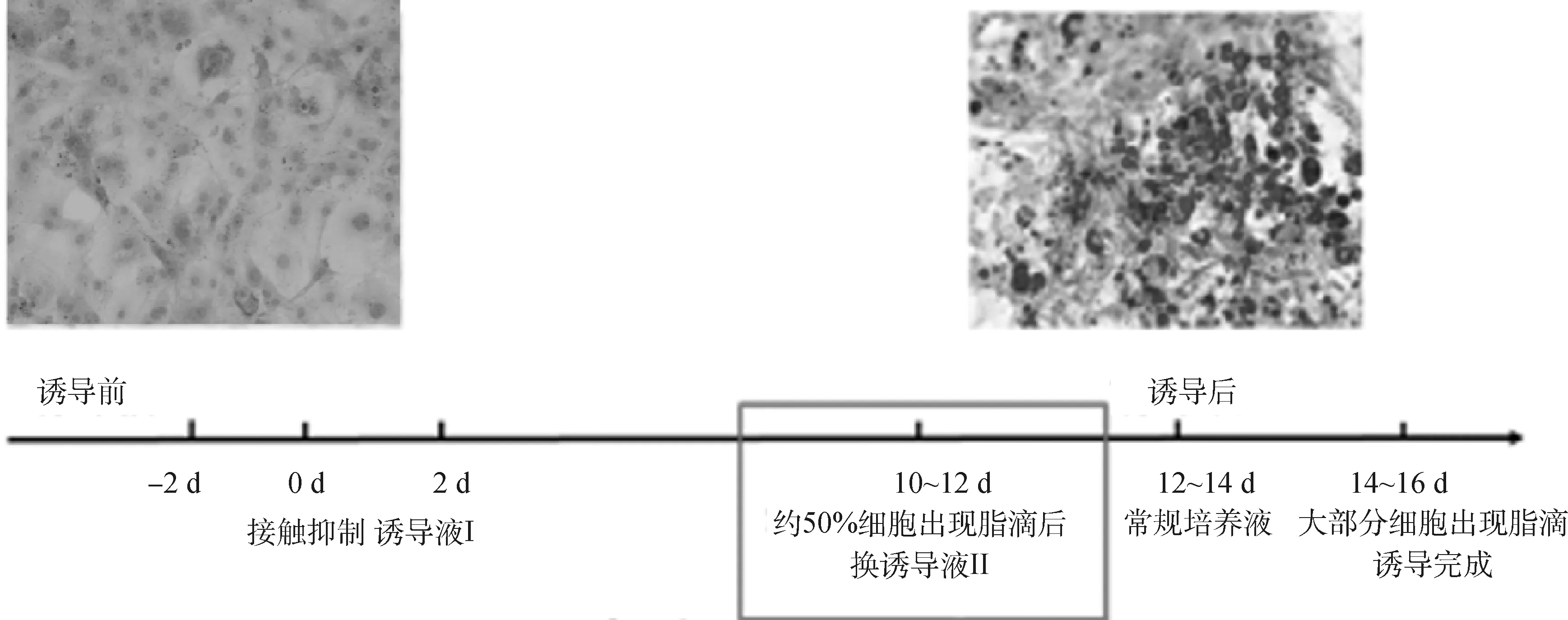
图1 诱导分化成功的3T3-L1脂肪细胞Fig.1 3T3-L1 adipocytes successfully induced and differentiated
2.2 DIZE对细胞活力的影响
MTT法检测经DIZE处理后的细胞活力,结果由图2所示,0~12.5 μmol·LDIZE处理24、48和72 h。对细胞活力无明显抑制作用,浓度达25 μmol·L时,细胞活力有所下降,至100 μmol·L时,细胞活力显著下降(<0.05)。同时结果显示,50 μmol·L以上的DIZE浓度处理72 h均可显著抑制细胞活力(<0.05)。因此,在后续试验中,确定DIZE作用浓度为12.5和25 μmol·L, 处理时间为48 h。
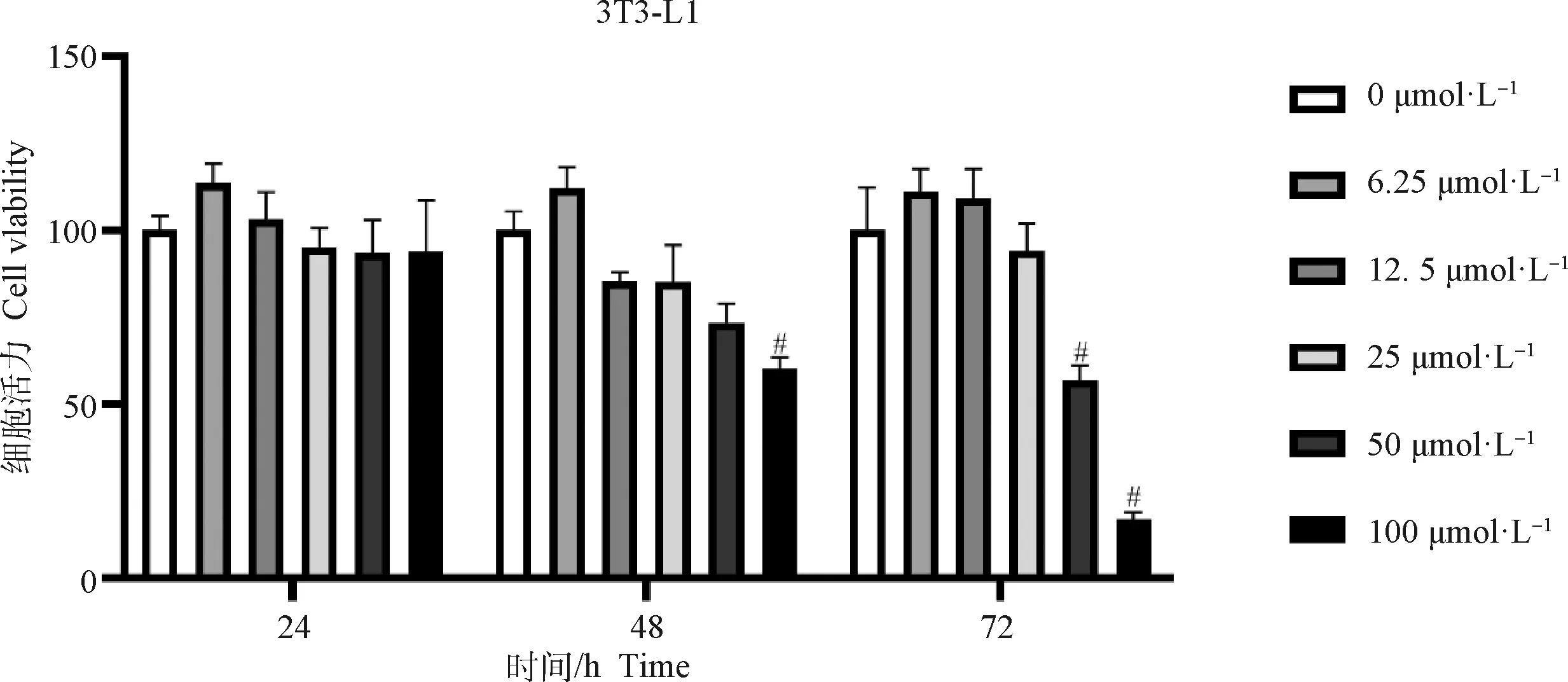
与对照组相比,#.P<0.05Compared with CON, #. P<0.05图2 DIZE对细胞活力的影响(n=6)Fig.2 The effect of DIZE on cell viability (n=6)
2.3 DIZE对3T3-L1脂肪细胞内脂滴合成的影响
2.3.1 DIZE对3T3-L1脂肪细胞上清三酰甘油和葡萄糖含量的影响 结果见表2。与对照组相比,不同浓度DIZE处理48 h后,均可极显著降低细胞上清中三酰甘油含量(<0.01),siACE2组显著降低(<0.05),siACE2+DIZE组极显著降低(<0.01)。 siACE2组较25 μmol·LDIZE组极显著升高(<0.01),但仍低于对照组;siACE2+DIZE组较25 μmol·LDIZE组无显著差异。即DIZE处理能够促进脂肪动员,降低3T3-L1脂肪细胞上清三酰甘油的含量。
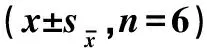
表2 DIZE对细胞三酰甘油和葡萄糖含量的影响
与对照组相比,不同浓度DIZE处理48 h后葡萄糖含量均显示上调,25 μmol·LDIZE组差异显著(<0.05)。siACE2组和siACE2+DIZE组较对照组和25 μmol·LDIZE组均显著下调(<0.05)。
2.3.2 DIZE对3T3-L1脂肪细胞内脂滴沉积的影响 油红O染色可以衡量出脂肪细胞内的脂肪蓄积程度,细胞内积累的脂质越多,油红染色结果越深。同时,利用吸光度检测对油红O染色结果进行定量分析。结果见图3。与对照组相比,不同浓度DIZE处理48 h后,细胞内脂滴显著减少(<0.05,图3B和3C),siACE2组和siACE2+DIZE组的脂滴蓄积较25 μmol·LDIZE组显著增加(<0.05,图3D和3E)。
A. 正常对照组; B. 12.5 μmol·L-1 DIZE组; C. 25 μmol·L-1 DIZE组; D: siACE2组; E. siACE2+DIZE组; F. 油红染色定量分析;图中数值均以“mean±Sem”表示。与对照组相比,#.P<0.05;与25 μmol·L-1DIZE组相比,*.P<0.05A. CON group; B. 12.5 μmol·L-1 DIZE group; C. 25 μmol·L-1 DIZE group; D. siACE2 group; E. siACE2+DIZE group; F. Quantitative analysis of oil red staining; Values represent the “means±Sem”. Compared with CON, #. P<0.05; Compared with 25 μmol·L-1 DIZE group,*. P<0.05图3 DIZE对3T3-L1脂肪细胞内脂滴合成的影响Fig.3 Effects of DIZE and siACE2 on lipid droplet accumulation in 3T3-L1 cells
2.3.3 DIZE对细胞内ACE2蛋白表达的影响 由图4可见,与对照组相比,不同浓度DIZE处理48 h均可上调3T3-L1脂肪细胞中ACE2的表达水平。25 μmol·LDIZE组较对照组差异显著(<0.05), siACE2组和siACE2+DIZE组较对照组和DIZE组显著下调(<0.05)。
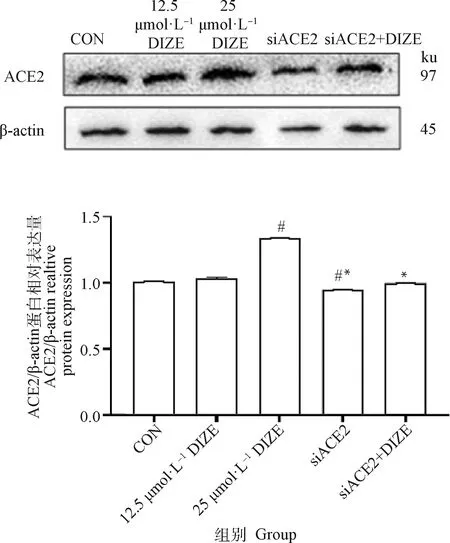
与对照组相比,#.P<0.05;与25 μmol·L-1DIZE组相比,*.P<0.05Compared with CON, #. P<0.05; Compared with 25 μmol·L-1 DIZE group, *. P<0.05图4 DIZE对3T3-L1脂肪细胞内ACE2蛋白表达的影响(n=6)Fig.4 Effect of DIZE on the expression of ACE2 protein in 3T3-L1 cells (n=6)
2.3.4 DIZE对3T3-L1细胞内FAS、ACC和SREBP1c蛋白表达的影响 结果见图5。由图5可以看出,与对照组相比,不同浓度DIZE处理48 h可下调3T3-L1脂肪细胞中的FAS表达水平,但无统计学差异。siACE2组的FAS蛋白表达较对照组和DIZE组显著下调(<0.05),siACE2+DIZE组的FAS蛋白表达较siACE2组有所下降,但仍高于DIZE组(<0.05)。
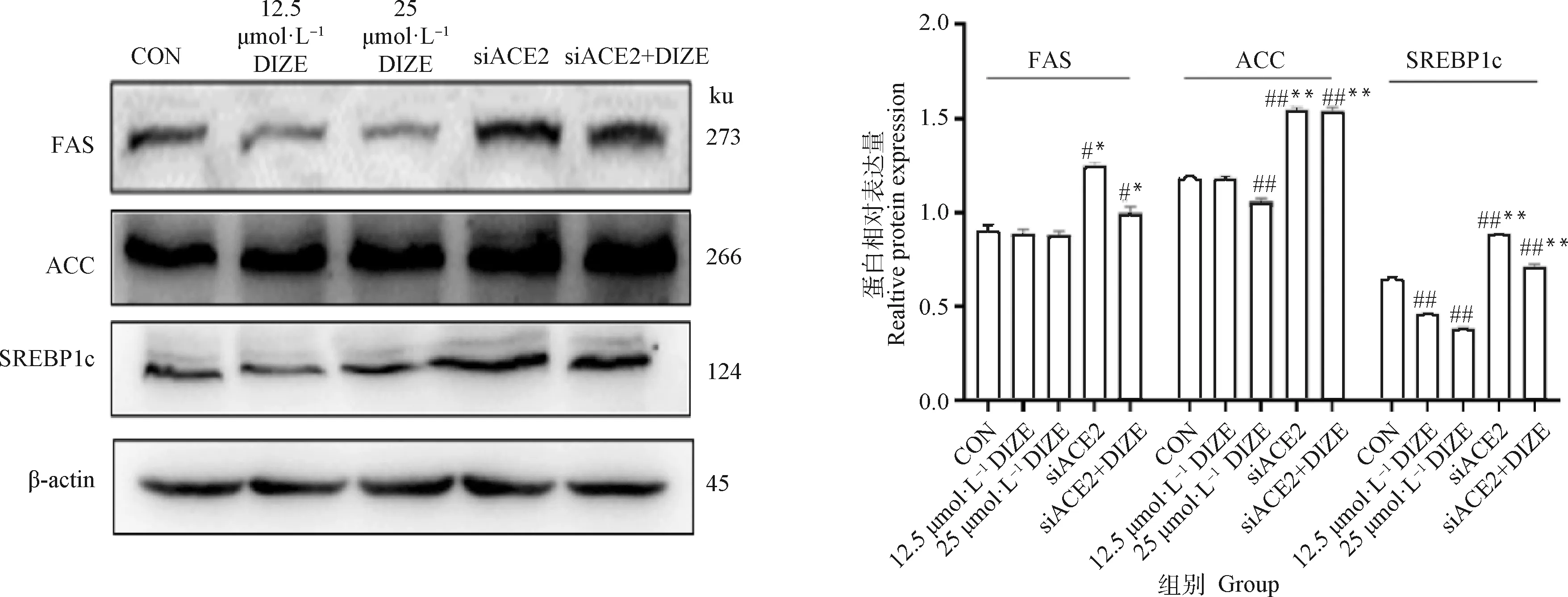
与对照组相比,#. 差异显著(P<0.05),##. 差异极显著(P<0.01);与25 μmol·L-1 DIZE组相比,*. 差异显著(P<0.05),**. 差异极显著(P<0.01)Compared with CON, #. P<0.05; ##. P<0.01; Compared with 25 μmol·L-1 DIZE group, *. P<0.05, **. P<0.01图5 DIZE对3T3-L1脂肪细胞内FAS、ACC和SREBP1c蛋白表达的影响(n=6)Fig.5 Effect of DIZE on FAS, ACC and SREBP1c protein expression in 3T3-L1 cells (n=6)
与对照组相比,不同浓度DIZE处理48 h可下调3T3-L1脂肪细胞中的ACC表达水平,25 μmol·LDIZE组ACC表达显著降低(<0.05)。siACE2组较对照组和DIZE组极显著上调(<0.01),siACE2+DIZE组较siACE2组有所下降,但仍高于DIZE组(<0.01)。
与对照组相比,不同浓度DIZE处理48 h可显著下调3T3-L1脂肪细胞中的SREBP1c表达水平(<0.01)。siACE2组较对照组和DIZE组极显著上调(<0.01),siACE2+DIZE组较siACE2组有所下降,但仍极显著高于DIZE组(<0.01)。
2.4 DIZE对3T3-L1脂肪细胞葡萄糖摄取和氧化的影响
葡萄糖转运蛋白GLUT4的表达结果见图6 A。与对照组相比,25 μmol·LDIZE组表达显著上调(<0.05),siACE2组和siACE2+DIZE组的GLUT4蛋白表达较对照组和25 μmol·LDIZE组均显著下调(<0.05)。即,DIZE处理可促进3T3-L1脂肪细胞的葡萄糖摄取。
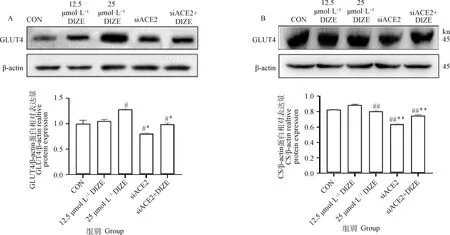
A. GLUT4蛋白表达变化;B. CS蛋白表达变化。与对照组相比,#.P<0.05,##. P<0.01;与25 μmol·L-1 DIZE组相比,*. P<0.05,**. P<0.01A. GLUT4 protein expression changes; B. Changes in CS protein expression. Compared with CON, #. P<0.05; ##. P<0.01; Compared with 25 μmol·L-1 DIZE group, *. P<0.05, **. P<0.01图6 DIZE对3T3-L1脂肪细胞葡萄糖摄取和氧化的影响(n=6)Fig.6 Effects of DIZE on glucose uptake and oxidation in 3T3-L1 adipocytes (n=6)
柠檬酸合酶CS的表达结果见图6 B。与对照组相比,12.5 μmol·LDIZE组表达有上调,但无统计学差异。25 μmol·LDIZE组表达水平极显著下调(<0.01)。siACE2组较对照组和25 μmol·LDIZE组均极显著下调(<0.01),siACE2+DIZE组较siACE2组有所升高,但仍极显著高于对照组(<0.01)。
3 讨 论
已有的活体研究数据显示,DIZE处理能够促进心肌细胞、肾、视网膜、肌肉组织还有脑组织中的ACE2表达,降低小鼠体重、血清胆固醇和三酰甘油的水平,但是与脂肪组织中ACE2的作用存在矛盾。De Macedo等研究显示,DIZE可以促进正常饮食小鼠附睾脂肪组织中ACE2 mRNA表达,而Bruce等则发现DIZE仅促进高脂饮食大鼠血清ACE2表达,对腹膜后脂肪组织ACE2 mRNA水平无明显作用。体外研究方面,DIZE可促进人视网膜色素上皮细胞和小鼠肺部Ⅱ型上皮细胞系(MLE-12)中的ACE2表达,但对人肝星状细胞系(LX-2)和小鼠枯否细胞系(KUP5)中的ACE2活性无显著促进作用。本研究在体外应用分化的3T3-L1前脂肪细胞进一步验证。首先,通过细胞活力检测筛选,确定了12.5、25 μmol·L2个给药浓度以及作用时间48 h。然后,用该药物处理时间、浓度及siACE2分别作用于分化后期的3T3-L1细胞,测定了细胞上清三酰甘油和葡萄糖含量,并对脂滴蓄积情况进行油红染色观察分析,以及检测细胞内ACE2的表达。DIZE处理使细胞上清三酰甘油含量减少,葡萄糖含量升高,提示DIZE处理可促进细胞脂肪动员及葡萄糖摄取。细胞油红染色结果显示,细胞脂滴显著减少,该浓度DIZE可促进细胞内 ACE2 表达。本试验还采用了siRNA 干扰技术下调 ACE2,进一步研究 DIZE 的作用。参照本实验室刘颖已建立的方法,利用 siRNA 转染技术,使3T3-L1细胞ACE2基因转录后表达沉默,ACE2基因表达降低,成功建立了ACE2基因表达降低或沉默细胞系,可用于验证DIZE内源性激活ACE2对细胞脂质沉积的抑制效应及其机制。结果显示,DIZE对ACE2的激活作用可被 siACE2 的预处理所消除,细胞油红染色结果也与其一致。该结果提示,DIZE通过激活 ACE2 表达而抑制脂质积累。
已知在脂肪组织中,先是乙酰 CoA 羧化酶在脂肪酸合成酶系(主要是乙酰 CoA 羧化酶(ACC)和脂肪酸合成酶(FAS))的作用下从头合成脂肪酸,然后脂肪酸酯化形成甘油骨架,由二酰甘油酰基转移酶(DGAT)介导合成三酰甘油(TG)。固醇调节元件结合蛋白-1c(SREBP-1c)是脂肪组织中SREBPs的主要表达形式,其在细胞核中与固醇反应元件结合,可诱导几乎所有参与TG和FA合成的必需基因的转录,能够直接启动 FAS和ACC转录翻译,从而促进TG合成。De Macedo等研究显示,口服DIZE可降低正常饮食小鼠ACC、FAS的基因转录,改善小鼠的代谢状况,减少脂肪沉积。本研究检测了FAS、ACC和SREBP1c蛋白表达的变化,结果显示,DIZE处理后,细胞内FAS、ACC和SREBP1c蛋白表达下降,且DIZE对FAS、ACC和SREBP1c蛋白的抑制作用可被siACE2消除,证实DIZE确实通过激活ACE2表达而抑制脂质积累,结果与文献[15]一致。提示,DIZE处理可通过激活ACE2表达来下调脂肪合成关键酶的蛋白表达,抑制细胞内的脂质蓄积。
有研究显示,激活 ACE2/Ang1-7/Mas 轴能够促进基础水平和胰岛素刺激下的葡萄糖转运蛋白的表达,降低肝糖原合成,缓解胰岛素抵抗。Takeda等研究发现,ACE2 敲除的正常饮食小鼠骨骼肌GLUT4表达显著降低,Ang1-7可以促进GLUT4在ACE2敲除小鼠中的表达,这一效应可被Mas受体阻滞剂A779 阻断。在脂肪组织方面,Mas受体缺失也导致胰岛素依赖的葡萄糖转运降低,GLUT4 表达降低。本试验结果显示,DIZE干预后GLUT4表达增加,与上述的研究结果一致,即DIZE处理脂肪组织葡萄糖转运增加。
本试验还对葡萄糖在细胞内的代谢去路作了进一步探讨。发现DIZE处理后,随着葡萄糖转运蛋白的表达上调,三羧酸循环关键酶柠檬酸合酶(CS)的表达也随之上调,即糖的有氧氧化(三羧酸循环)激活,糖分解代谢加强。结合DIZE抑制成脂因子表达的结果,说明DIZE干预促进摄取的葡萄糖主要进入了糖的有氧氧化途径,而不是进入成脂途径,间接抑制了脂肪酸的从头合成。
4 结 论
DIZE可以通过激活ACE2减少细胞脂质沉积,提示ACE2或DIZE改善脂质沉积的研究和开发潜力,也为从ACE2的角度对日益增长的肥胖和代谢综合征的治疗提供新的思路。
猜你喜欢
家庭科学·新健康(2020年6期)2020-07-06
科学导报(2019年45期)2019-09-23
保健与生活(2019年23期)2019-09-10
恋爱婚姻家庭·养生版(2018年6期)2018-08-01
分析化学(2018年1期)2018-01-18
中学科技(2017年11期)2017-12-26
科学中国人(2016年9期)2016-11-04
中学生数理化·八年级物理人教版(2015年10期)2016-01-04
健康博览(2015年3期)2015-07-03
科学大众(中学)(2009年8期)2009-09-06
