
烘焙预处理对小球藻热解特性与产物分布的影响
2022-08-17 06:38谢东明
可再生能源 2022年8期
谢东明, 钟 典, 李 俊, 曾 阔, 彭 静
(1.广西壮族自治区特种设备检验研究院, 广西 南宁 530200; 2.华中科技大学 能源与动力工程学院, 煤燃烧国家重点实验室, 湖北 武汉 430074; 3.华中科技大学 中欧清洁与可再生能源学院, 湖北 武汉 430074)
0 引言
藻类作为第三代生物质,因其具有生长速度快、光合效率高、不占用耕地、生物油产率高、固碳能力强等优点,近些年受到了人们的重视[1]~[4]。微藻中含氮量为5.2%~16.3%[2],[4],[5], 其热解生物油中包括酰胺类、胺类、吡咯类、喹啉类、吡啶类、吲哚类等含氮物质。 早先的研究集中在将含氮物质从中裂解出去,生成烃类、氨等物质作为运输燃料。 近年来,这些含氮物质可作为一些精细化学品的重要前驱体而受到了研究者的广泛关注[4],[5]。如何通过更优的处理方式实现藻类生物质热解高值化利用成为了新的挑战。
烘焙(200~300 ℃)是生物质常见的预处理方式,可有效地提高生物质原料的能量密度,改善其机械性能, 并能在一定程度上解决生物质,特别是藻类生物质含水率高、热值低、不易储存等问题,降低运输、处理、储存以及作为能源利用的成本[6]。 烘焙预处理对生物质热解也具有深远的影响。 研究表明,随烘焙温度的升高,热解生物油的品质有所提高,水分含量最多减少了42.8%,稠环化合物及酚类产物的含量有所增加,热值约升高34%,但是生物油的产率大为降低,焦炭的含量增加,其化学组成逐渐向煤过渡[7]。 此外,烘焙时间越长,烘焙温度越高,热解时的交联反应和碳化缩合反应就越多, 使热解过程中水分和CO2析出减少, 且水的析出温度由120 ℃提升至200℃[8]。这表明,需要对整个生产过程进行综合考虑,确保生物油含氧含水量高的缺点被合理补偿。 文献[9]指出,在较高烘焙温度下,木质素的侧链断裂,导致具有侧链的酚类产量减少,不含有侧链的酚类产量增加。 宇妍[10]探究了不同脱水方式与烘焙联合预处理对生物质燃料特性的影响, 经过联合预处理进一步提升了生物质的能源化利用品质。 藻类生物质与木质纤维素类生物质成分差异较大, 烘焙预处理对其热解特性与产物分布的影响是否类似, 以及对含氮产物生成的影响仍有待研究。
本文在不同烘焙预处理条件下对小球藻热解特性的影响规律进行了研究。 通过在不同升温速率下的热重实验,探究其在40~900 ℃下的热重反应特性, 采用分布式活化能模型(DAEM)计算烘焙后小球藻的热解活化能,揭示烘焙预处理对其热解动力学的影响;在升温速率为10 ℃/min 的条件下采用热重-红外联用仪分析烘焙后小球藻热解气体中含氮化合物的变化规律;采用Py-GC/MS 在不同热解温度(500 ℃和900 ℃) 下探究烘焙预处理对其热解生物油的影响规律。
1 实验原料与方法
1.1 实验原料
本文所用的小球藻(干燥基)使用前先在105℃烘箱中干燥24 h,再手工磨碎,并用30 目筛网筛分后得到粒径小于0.613 mm 的干燥原料,其工业分析 (GB/T 28731—2012)、 元素分析(Vario EL cube,德国元素分析系统公司)以及高位热值(微电脑热量计系统,WZR-1T-CII & IKAC2000)结果如表1 所示。

表1 小球藻原料性质Table 1 Proximate and ultimate analysis of Chlorella
1.2 实验仪器与设备
烘焙预处理实验装置如图1 所示。
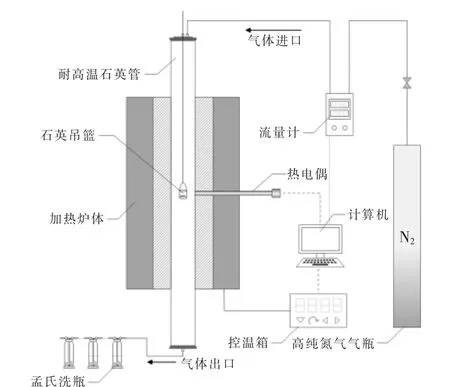
图1 烘焙实验装置示意图Fig.1 Schematic diagram of the experimental setup
称取2~5 g 干燥好的小球藻粉末, 装入石英吊篮中;将石英吊篮固定在耐高温石英管顶部,通入氮气 (纯度99.999 9%,245 mL/min)30 min,对石英管进行排空; 将管式炉预热至所需温度(200,225,250,275,300,325 ℃, 温度间隔25 ℃)并保持恒温;待炉体温度达到所需温度后,降低载气流量至170 mL/min,同时将装入样品的石英吊篮推入反应器中部, 分别保持相应的停留时间(30,60,90 min)。 在实验过程中,采用三段式洗涤去除尾气中的污染物和焦油, 之后排放至室外(250 mL 孟氏洗瓶, 前两个装入异丙醇和玻璃珠的混合物,第3 个装入变色硅胶)。实验结束后,取出吊篮进行称重,获得烘焙前后的质量变化。每次实验至少重复3 次,且相对误差不超过5%,取3次的平均值计算质量产率、 相对能量密度和能量产率。

式中:YM为生物质烘焙后的质量产率,%;WM为生物质烘焙后的相对能量密度,%;YE为生物质烘焙后能量产率,%;MT为烘焙后收集的生物质质量,g;M0为烘焙前实验称取的生物质质量,g;HHV0,HHVT分别为烘焙前、 后生物质的高位热值,kJ/kg。
对不同条件下烘焙预处理的产物与原样进行热重实验。 升温速率设置为5,10,20 ℃/min,在氩气氛围下从40 ℃升温至900 ℃,同时采用同步热分析-红外光谱联用仪(STA449F3Jupiter,德国耐驰)对挥发分进行红外光谱分析。 采用裂解器-气相色谱质谱联用仪(CDS5200,美国CDS-Agilent)对相同几组样品进行热解实验, 热解温度设置为500 ℃和900 ℃,升温速率为2 000 ℃/s,保持30 s。气相色谱-质谱仪(GCMS)在线检测样品的挥发分组成,采用氩气气氛,色谱柱以30 ℃/min程序升温至300 ℃,分流比为1∶50,质谱检测的核质比为35~600 m/z。
2 结果与讨论
2.1 烘焙对小球藻热解动力学的影响
2.1.1 小球藻烘焙前后的热解失重特性
选取烘焙时间为60 min,烘焙温度为225,250,275 ℃时的烘焙产物及小球藻原样4 个样品进行热重动力学测试, 得到生物质随温度的变化关系。 为了更好地对热重数据进行分析,将DTG曲线中的最低点定义为峰值失重率, 所对应的温度称为峰值失重温度。
图2 为小球藻原样及3 个烘焙样品的TG 与DTG 曲线。 由图2 可以看出,在不同升温速率下,焦炭产率、峰值失重率都发生了变化,且开始失重的温度随升温速率的升高而逐渐升高。 对比不同烘焙条件下的DTG 曲线可以看出:随着烘焙温度的上升, 样品在所有升温速率下的主失重峰(300~400 ℃)均出现滞后现象,对应的温度升高(未烘焙样品:278~312 ℃;225 ℃烘焙后:302~327℃;250 ℃烘焙后:325~352 ℃;275 ℃烘焙后:356~383 ℃);样品在主失重峰的失重率也随烘焙温度的上升而减小, 而对应的400~500 ℃处肩峰的失重率则会上升。 研究指出,微藻三组分的热重曲线会出现2~3 个峰, 在300~400 ℃的主峰主要是由于糖类和蛋白质的热降解脱出挥发分所致,400~500 ℃的肩峰是由于脂类的热降解所致[2],[3]。从DTG 曲线还可以看出, 随着烘焙温度的升高,主峰与肩峰的间距变小,由于脂类热解区间较高,在烘焙过程中基本不发生变化, 而较易降解的糖类和蛋白质在该阶段部分发生脱挥发分反应[11],烘焙使小球藻的均一性得到提高。

图2 小球藻在不同升温速率下TG 和DTG 曲线Fig.2 TG and DTG curves of Chlorella at different heating rates
不同热解条件后的焦炭产率计算式分别为

式中:φrel为相对焦炭产率,%;φabs为绝对焦炭产率,%;Mresidual为焦炭质量,g。
不同样品在不同升温速率下的小球藻焦炭产率如图3 所示。从图可以看出,样品最终焦炭产率受到烘焙条件和升温速率变化的影响, 且烘焙条件的影响占主要地位。 以烘焙后的样品质量计算相对焦炭产率, 烘焙处理明显提高了相对焦炭产率,且随着温度升高而升高,以升温速率为10℃/min 为例, 当烘焙温度从225 ℃提高至275 ℃时,小球藻相对焦炭产率由15.76%增加至35.91%[图3(a)]。 提高升温速率会使原始样品与经过烘焙处理后样品的相对焦炭产率差异加大。
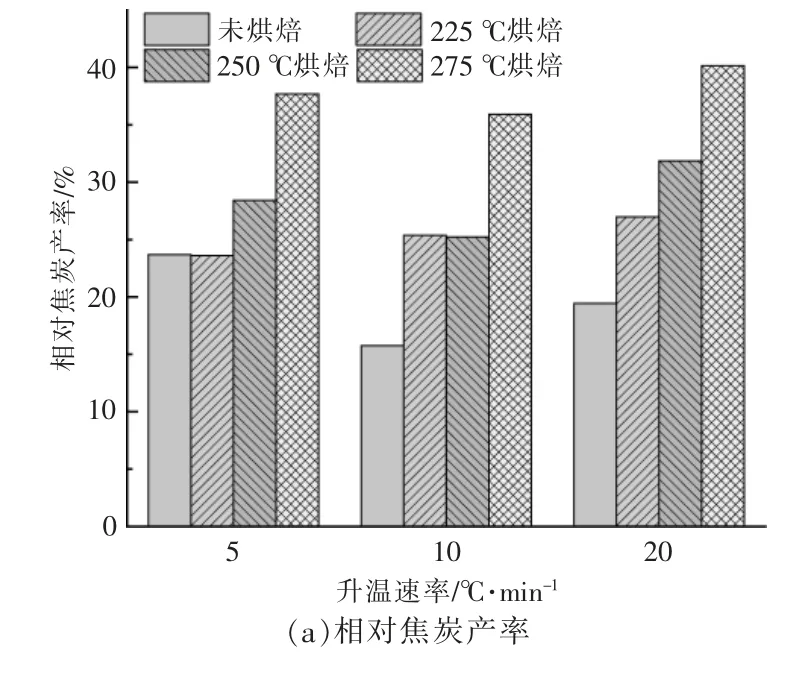
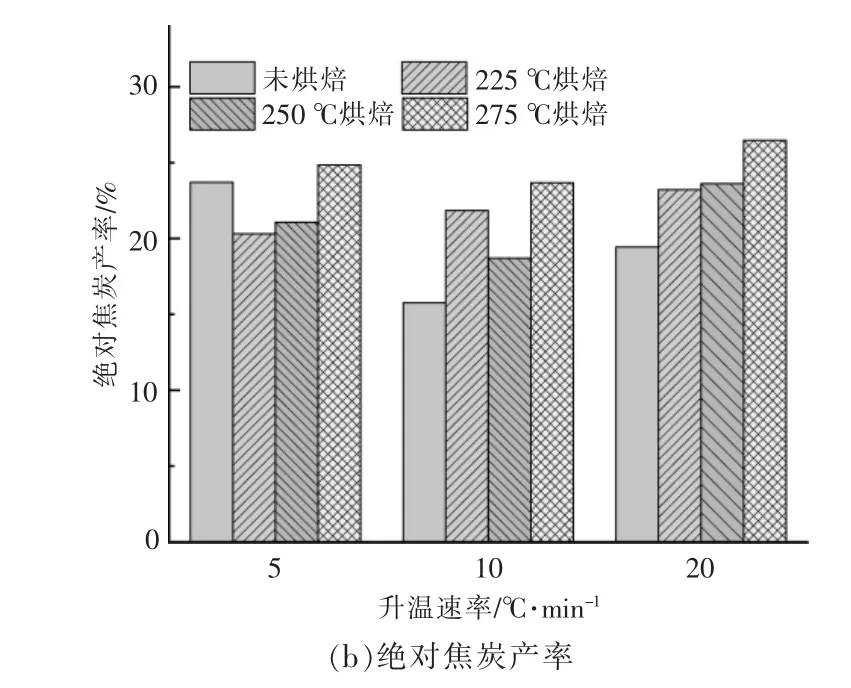
图3 在不同升温速率下小球藻烘焙前后的焦炭产率Fig.3 Residual rates of Chlorella before and after torrifying at different heating rates
若考虑烘焙过程中脱出的挥发分, 即以烘焙前原始样品的质量计算绝对焦炭产率,如图3(b)所示。此时随着烘焙温度的升高,绝对焦炭产率依旧呈现上升趋势, 说明烘焙温度升高会促进最后热解焦的生成。 在升温速率为5 ℃/min 时,225 ℃和250 ℃烘焙的小球藻的绝对焦炭产率甚至低于未烘焙原料,说明225~250 ℃烘焙处理后,小球藻在低升温速率(5 ℃/min)下热解更有利于挥发分的析出。
2.1.2 小球藻烘焙前后的热解动力学
热解活化能是动力学的重要参数, 生物质热解是众多官能团在热能作用下一系列复杂连续的反应。由多条热重曲线得出的分布式活化能,计算结果较准确且随反应的加深而变化, 避免了单一表观活化能生物质复杂反应的局限性[2],[12]~[14]。 本文采用活化能分布模型(DAEM)研究小球藻热反应过程,探讨热解活化能与转化率的关系[2],[12]~[14]。

式中:V/V*为热解转化率,%;T 为温度,K;A 为指前因子,s-1;R 为气体常数,R=8.314 J/(mol·K);E为活化能,kJ/mol;w 为τ 时刻样品质量,g;w0为样品初始质量,g;w∞为样品热解后最终剩余量,g。
对热重实验中升温速率为5,10,20 ℃/min 的热重数据进行计算整理后,得到小球藻的Arrhenius谱图及热解活化能随转化率的变化关系, 分别如图4,5 所示。 从图4 可以看出,当转化率为0.05~0.80 时, 拟合曲线的相关系数R2整体上均大于0.98 000,说明拟合效果较好,计算得到的活化能也较为可靠。从图5 可以看出,烘焙预处理总体上会提升小球藻的热解活化能, 但不同反应阶段的影响不尽相同。 当转化率低于0.2 时,仅225 ℃烘焙后的样品初始阶段活化能稍微大于原样品,其余烘焙温度样品的活化能相比原样差别不大;当转化率为0.2~0.55 时, 不同烘焙温度下小球藻的热解活化能与原样相比均明显增大, 但烘焙温度对活化能的增加影响不大。 此阶段大致与图2 中最大失重峰对应,即蛋白质与糖类的分解阶段,说明烘焙处理使得糖类和蛋白质经历了预热解过程, 生成了一些更难以分解的产物; 当转化率为0.55~0.75时,不同烘焙温度下样品的热解活化能出现差异,这一阶段中225 ℃烘焙的活化能最高,但随烘焙温度升高活化能反而逐渐降低, 与原始样品接近。此阶段大致与图2 中肩峰对应,即脂质的分解阶段, 说明虽然烘焙处理会使脂质变为更难分解的产物,但提高烘焙温度反而会逐渐消除这一影响,使热解反应更容易进行。 在最后阶段,烘焙处理的样品活化能均出现激增, 以275 ℃烘焙样品为代表,此阶段对应了焦炭的生成,说明烘焙处理后焦炭产物增加,难以再进行分解。这也对应了图3 中的结论。
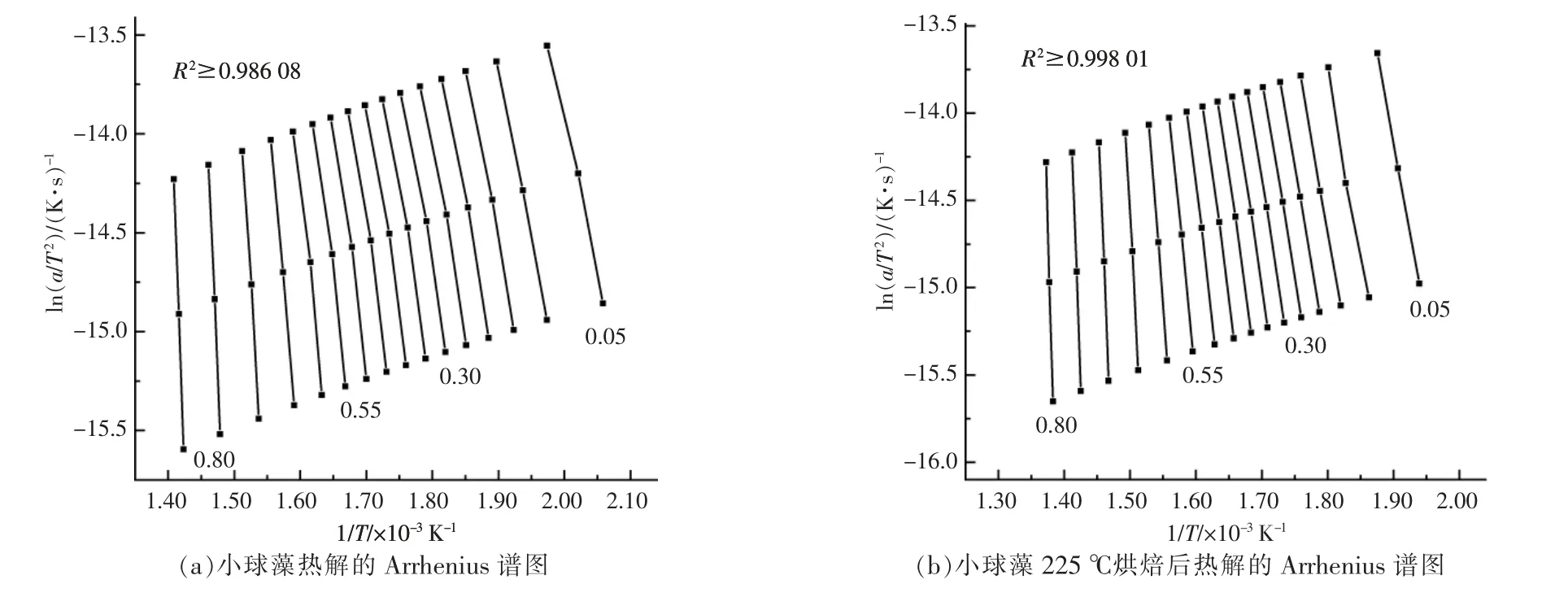

图4 小球藻热解动力学计算Fig.4 Calculation of pyrolysis kinetics of Chlorella vulgaris
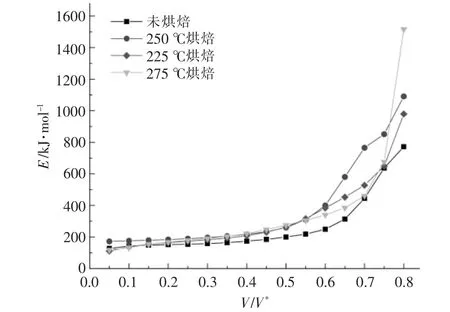
图5 不同烘焙温度下小球藻的热解活化能随转化率的变化关系Fig.5 Changes of activation energy of Chlorella vulgaris at different torrification temperatures
2.2 烘焙对小球藻热解气体产物的影响
根据Lambert-Beer 定律[15],各挥发分气体的红外光谱的吸光度与气体的浓度呈线性关系,特征吸收峰越高,说明该气体的相对含量越高。 图6为小球藻原样及烘焙预处理后热重-红外联用仪所得到的红外三维立体图。由图可以看出,各个样品热解挥发分的红外谱图中均可见几个显著的吸收峰, 分别在3 200~3 400 cm-1,2 950 cm-1,2 340 cm-1,1 754 cm-1,1 500~1 600 cm-1,900~1 100 cm-1,669 cm-1。 其中大部分吸收峰随反应时间变化,在热解40 min 左右均出现最大值, 对应温度为280~300 ℃, 结合各组样品DTG 曲线可以发现,该温度区间为最大失重峰出现的温度区间。
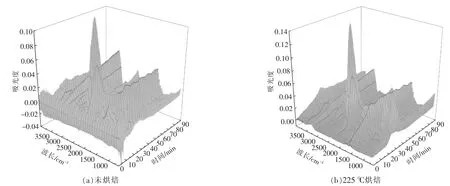
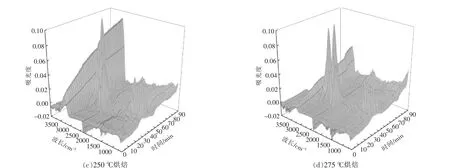
图6 小球藻烘焙预处理后在10 ℃/min 下热解时气体的FTIR 分析Fig.6 FTIR analysis of pyrolytic volatile fraction of Chlorella vulgaris after pretreatment at 10 ℃/min
图7 展示了上述4 种样品在失重速率最大处的挥发分鉴定:①3 200~3 400 cm-1为羟基的吸收峰,对应挥发分中结合水的生成;②2 950 cm-1为C-H 键的伸缩振动峰,对应挥发分中的轻质烷烃类物质的生成;③2 340 cm-1与669 cm-1对应CO2吸收峰,标志热解脱-CO 以及脱-COOH 反应;④1 754 cm-1对应有机组分中的C=O 结构, 主要存在于脂质热解产物与有机酸产物中;⑤1 500~1 600 cm-1主要是芳烃中的C=C 键的吸收峰,对应产物中芳烃类物质的生成;⑥900~1 100 cm-1附近的峰由含氧官能团的吸收峰组成, 如有机组分中C-O 和无机组分中的P-O-P,Si-O,对应醇类、酚类以及无机硅磷酸盐[16]。
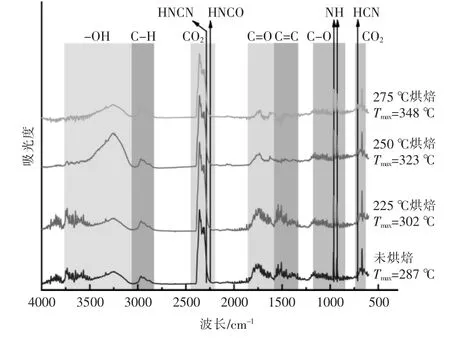
图7 小球藻烘焙预处理前后在失重速率最大处的挥发分鉴定Fig.7 Identification of volatile fraction at the maximum rate of weight loss of Chlorella before and after torrification
对比烘焙处理前后以及不同烘焙温度下的样品热解挥发分的红外谱图(图7)可以发现,相比未烘焙的原样,烘焙后样品热解产生的水变少,含氧官能团如羧酸、醛、酮等的析出减少,芳烃类物质减少,醇、醚以及烃类则没有明显变化。此外,随着烘焙温度的升高,CO2的含量经历了一个先增加后减少的过程, 在烘焙温度为225 ℃时达到峰值,但总的强度仍高于未烘焙原样。
小球藻热解产物中的4 种主要含氮官能团(NH,HCN,HNCO 和HNCN) 随热解温度的变化如图8 所示。总体来看,烘焙处理使小球藻热解含氮物质增加, 但同时烘焙温度的升高也会降低后续热解含氮产物的生成。 图8(a),(b)分别为不同烘焙温度样品的NH3和HCN 的生成强度, 可以看到,烘焙处理对其强度影响规律相似,生成的含氮物质强度由高到低依次为225 ℃烘焙>250 ℃烘焙>275 ℃烘焙>未烘焙。 这是由于烘焙温度升高,部分含氮的产物在烘焙阶段就提前释放,导致热解阶段的含氮产物析出减少;图8(c),(d)中,HCNO 和H2NCN 的含量由高到低依次为225 ℃烘焙>275 ℃烘焙>250 ℃烘焙>未烘焙, 其中烘焙时间均为60 min。 但这两种相对复杂的含氮物质的析出强度并不和烘焙温度呈线性相关, 而是在250 ℃出现极小值。 这可能是由于生成过程会受到脂质与糖类烘焙产物的影响, 进而影响了在后续热解过程中交互脱氮的过程[17]。
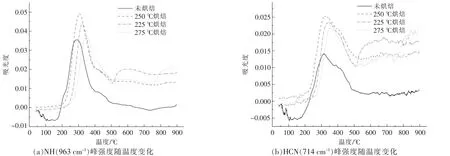

图8 小球藻烘焙前后含氮物质随温度的变化关系Fig.8 Relationship between nitrogenous substances and temperature before and after torrification of Chlorella vulgaris
2.3 烘焙对小球藻热解生物油组分的影响
为了便于分析, 根据原样和烘焙样品在不同热解温度下总离子流色谱图峰面积的占比, 选取了数十种主要热解产物。 将生成产物分为轻质挥发分、杂环化合物、脂肪族含氧化合物、其他含氧化合物及其他产物5 大类, 各类产物的占比如图9 所示。

图9 小球藻及烘焙样品在不同温度下的热解产物分布Fig.9 Distribution of pyrolysis products of Chlorella and torrified samples at different temperatures
由图9 可以看到, 热解温度为900 ℃下的热解产物中,轻质挥发分、杂环化合物、含氧化合物所占比例均低于500 ℃下的热解产物(烘焙温度为250 ℃时的轻质挥发分除外),然而其他产物的占比大幅度增加,导致最终生物油的产率增大。这主要是由于500 ℃下原先检测不到的重质生物油在热解温度提升至900 ℃的高温环境下发生裂解,生成了更多以轻质芳烃和不饱和烃类物质(碳原子个数小于6)为主的轻质挥发分。
小球藻的热解产物中含氮产物较多, 含氮物质主要包含在杂环类物质中, 这是由于藻类的构成与木质纤维素类生物质不同,蛋白质、脂类、糖类是其主要成分[11],而蛋白质中的氨基酸侧链是N 元素的主要来源。 对于在500 ℃下的热解产物而言,杂环类化合物主要由吲哚、吡咯类、喹啉类、吡啶类、肽类和吡喃类组成,脂肪族含氧化合物主要由乙酸、羟基丙酮、长链脂肪酸及其衍生物长链脂肪酰胺类组成, 其他含氧化合物主要由苯酚类组成,此外还有部分C-O-N=C 结构及酯类结构,其他产物主要由胺类、甲苯类、腈类(C≡N 键)等组成。在900 ℃下的热解产物,不同之处在于其他产物中的种类增加,主要由苯、甲苯、萘、茚等芳香类产物以及二烯类、丙烯腈等组成。
当小球藻在500 ℃下热解时,由图9 可以看出:随着烘焙温度的升高,小球藻热解产物中的杂环化合物逐渐增加,只有烘焙温度为225 ℃时的产率低于未经烘焙预处理小球藻原样;含氧化合物先增加后减少,在烘焙温度为250 ℃时达到最高; 其他产物先减少后增加, 在烘焙温度为250 ℃时产率达到最低, 且均低于未经烘焙预处理原样。 当小球藻在900 ℃下热解时,杂环化合物占比在烘焙温度为250 ℃时达到最低,略高于未经过烘焙预处理的原样产率;含氧化合物和其他产物随烘焙温度的升高先增加后减少,在烘焙温度为250 ℃时达到最高,二者的不同之处在于含氧化合物的所占比例变化不大,而其他产物远远高出未烘焙原样的产率。 这说明,高温下重质生物油的热裂解过程几乎不会产生含氧化合物,以轻质芳烃和烯烃类为主, 经过烘焙预处理后,更容易发生重质生物油的裂解,最佳烘焙温度为250 ℃。
3 结论
本文通过TG-FTIR 与Py-GC/MS 方法,探究了烘焙预处理和不同烘焙温度对小球藻热解动力学及其产物的影响,得到以下结论。
①烘焙预处理的峰值失重温度随烘焙温度的升高而升高,同时两个主要失重峰的差距减小,提高了原料组分的均一性; 提升烘焙温度能降低部分活化能, 但烘焙处理后的小球藻总体热解反应活化能仍会增加。
②烘焙预处理促进了小球藻热解脱羧过程,产生更多CO2; 同时会使小球藻在热解过程中含氮产物的生成显著增加, 提升烘焙温度能够抑制这一趋势。
③小球藻的热解生物油中含氮物质较多,以吲哚、吡咯类、喹啉类为主,随着烘焙温度的升高,生物油中的杂环化合物比例增加。 当小球藻在900 ℃下发生热解时,以生成轻质芳烃和烯烃类产物为主,几乎不产生含氧化合物,且烘焙预处理更容易使生物油发生裂解,最佳烘焙温度为250 ℃。
猜你喜欢
矿山安全信息(2022年4期)2022-11-25
矿山安全信息(2022年8期)2022-11-25
南方农业(2022年12期)2022-11-23
科学技术创新(2022年28期)2022-10-21
金属世界(2022年4期)2022-07-29
可再生能源(2022年5期)2022-06-09
海洋湖沼通报(2022年2期)2022-05-08
水产养殖(2022年1期)2022-03-03
今日农药(2017年10期)2017-11-14
山东工业技术(2017年12期)2017-07-06
