
保加利亚乳杆菌噬菌体CC34 裂解基因的克隆表达及功能鉴定
2022-08-17 07:00董右铭
农产品加工 2022年13期
卢 野,董右铭
(四平市产品质量检验院,吉林 四平 136000)
噬菌体(bacteriophage) 是感染细菌的一种病毒,噬菌体分布极广,凡是有细菌的场所,就可能有相应噬菌体的存在[1]。由于其可以在宿主体内快速复制及繁殖的特性,目前被广泛用于抑制多种食源性致病菌的感染[2-3]。但在酸奶发酵过程中所需要的乳酸菌也逃脱不了噬菌体的感染,造成乳酸菌产酸量降低产生的品质问题,从而导致生产方的经济损失[4-6]。酸奶目前销售量不断上升,人们对酸奶饮品的青睐也日益提高,但噬菌体的感染使酸奶在发酵过程中风味变差,品质下降甚至产酸终止等现象一直困扰着酸奶业。目前,关于乳酸菌噬菌体的分离,酸奶发酵中噬菌体污染的预防已有很多报道,但对乳酸菌噬菌体的功能基因开发、鉴定却研究很少[7-8]。试验对酸奶中分离到的保加利亚乳杆菌噬菌体CC34 的裂解基因进行克隆、表达及功能的初步鉴定,为酸奶工业噬菌体污染的预防奠定基础[9-10]。
1 材料与方法
1.1 材料
1.1.1 菌株、毒株与质粒
大肠杆菌 DH5α(TaKaRa)、pET-28b(+)(Novagen)、保加利亚乳杆菌(Lactobacillus bulgaricus)、金黄色葡萄球菌(Staphylococcus aureus)、大肠杆菌(Escherichia coli)、噬菌体CC34,实验室提供。
1.1.2 试剂
凝胶回收试剂盒、质粒提取试剂盒、基因组DNA 提取试剂盒、各种限制性内切酶、T4 连接酶、Taq 酶,大连宝生物公司 (TaKaRa) 提供;DNA 纯化试剂盒,长春库美生物科技有限公司提供;Gold erView染料(SBS)、中范围蛋白质标准分子量(MBI),长春翊博生物科技有限公司提供;TAE 电泳缓冲液,Tris 缓冲液 (pH 值 7.2)、PBS 缓冲液 (pH 值 7.2)、Tris - 甘氨酸电极缓冲液,由实验室事先配制。
1.2 试验方法
1.2.1 裂解基因e 的PCR 扩增
根据噬菌体CC34 片段测序结果,对e 基因开放阅读框设计一对特异性引物(ATAGCCATGGCAGGCATTCGTTG,下划线为Nco Ⅰ酶切位点) 及(ATG-GGTCGACGGTAAGCCTGGAA,下划线为SalⅠ酶切位点),由大连TaKaRa 公司合成。以噬菌体基因组DNA 为模板,扩增条件为94 ℃,3 min;94 ℃,45 s;57 ℃,45 s;72 ℃,90 s;总共循环 30 次;72 ℃延伸10 min。扩增结束后取2 μL PCR 产物通过琼脂糖凝胶电泳检测条带大小,在紫外照射下回收目的片段,根据TaKaRa 凝胶回收试剂盒说明书对e 基因的PCR 扩增产物回收纯化,将产物放置在-20 ℃下保存。
1.2.2 表达载体pET- 28b(+)- e 的构建及鉴定
扩大培养含有pET-28b(+)质粒的大肠杆菌,按照TaKaRa 质粒小提试剂盒提取pET-28b(+)质粒,与1.2.1 得到的胶回收产物分别用NcoⅠ和SalⅠ2种限制性内切酶进行双酶切,酶切产物经电泳检测后纯化回收2种产物,以质粒∶DNA = 1∶10 的比例用T4 连接酶连接过夜,将连接产物转化至大肠杆菌DH5α 感受态细胞,过夜培养后挑取疑似阳性菌落通过双酶切进行鉴定是否转化成功。
1.2.3 pET- 28b(+)- e 的 IPTG 诱导表达
将转化成功的pET-28b(+)-e 的大肠杆菌BL21接种至5 mL LB 液体培养基(含25 μg/mL 卡那霉素)中,于37 ℃下以转速200 r/min 振荡培养过夜;以1%的接种量接种至50 mL LB 液体培养基(含25 μg/mL卡那霉素) 中,培养温度及转速与上述步骤相同,OD600达 0.4~0.6; 加 入 500 μL IPTG 至 终 浓 度 为1 mmol/L 诱导,继续以相同的培养温度及转速培养,每间隔1 h 取诱导后的菌液1 mL;将菌液离心后分别收集上清液和沉淀物,沉淀物用50 μL PBS 重悬沉淀,于-20 ℃下保存待用。
1.2.4 pET- 28b(+)- e 表达产物 SDS- PAGE分析
细菌沉淀加入2×SDS 样品缓冲液50 μL,充分混匀后,放置沸水中煮5 min,配制12.5%的SDSPAGE分离胶,5%浓缩胶,取10 μL 样品电泳,电泳结束后用银染法显色。
1.2.5 pET- 28b(+)- e 细菌的溶菌试验
将1.2.3 步骤中诱导表达的细菌沉淀分别用TBS和PBS 重悬,冰浴超声 10 s,间隔 10 s,30~50 循环,直至样品变清亮;超声完成后,于4 ℃下以转速12 000 r/min 离心20 min,转移上清进行溶菌试验。取100 μL 生长到对数期的指示菌(保加利亚乳杆菌、金黄色葡萄球菌、大肠杆菌)分别与3 mL LB半固体培养基混匀,均匀铺在有下层LB 固体培养基上。将无菌的牛津杯放在铺有指示菌的上层琼脂上,依次加入 100 μL 的 pET-28b(+)-e,pET-28b(+)(空白对照) 的超声上清,于30 ℃下培养过夜,第2 天观察溶菌圈。
2 结果与分析
2.1 裂解基因e 的PCR 扩增结果
以噬菌体基因组为模板,可以扩增得到一条大小在490 bp 左右的特异性条带,与目的片段e 基因的大小相符。
裂解基因e 的PCR 扩增结果见图1。

图1 裂解基因e 的PCR 扩增结果
2.2 表达载体pET-28b(+)-e 的双酶切鉴定结果
用 Nco Ⅰ和 Sal Ⅰ对构建好的 pET-28b(+)-e 载体进行双酶切,得到5.3 kD 和490 bp 2 条片段(图2),分别对应pET-28b(+)和e 基因的大小,与预期结果一致。
双酶切鉴定结果见图2。
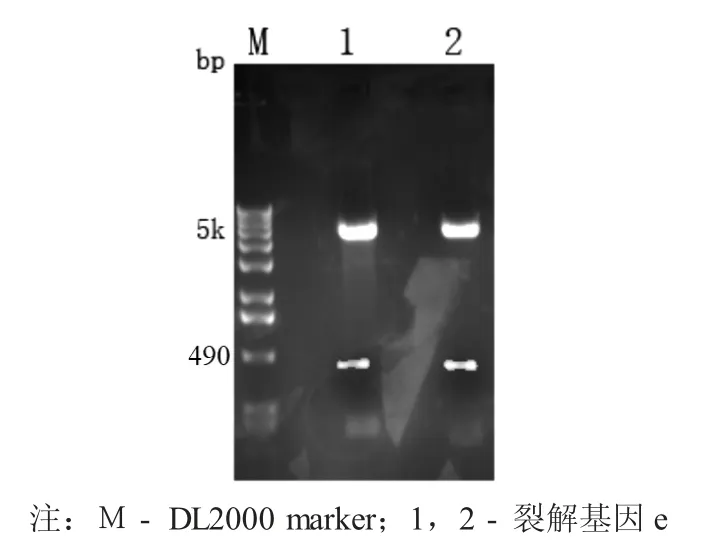
图2 双酶切鉴定结果
2.3 pET-28b(+)-e 表达产物 SDS-PAGE分析
pET-28b(+)-e 的细菌沉淀总蛋白及表达上清总蛋白在18 kD 附近都有一明显的蛋白表达带(黑箭头所指),分子量大小与预期相符。pET-28b(+)-e在诱导表达后的第2,第3 小时的表达量最大,之后减少。与之相反的是pET-28-e 的表达上清的目的蛋白在诱导表达的后期(4,5,6 h) 却增加(图4)。出现这种情况的原因是pET-28-e 在诱导表达时会出现溶菌,表达宿主的大量死亡,使本来胞内表达的e蛋白释放到培养液里。
pET-28b(+)-e 细菌沉淀总蛋白的 SDS-PAGE见图3,pET-28b(+)-e 表达上清总蛋白的SDS-PAGE见图4。

图3 pET-28b(+)-e 细菌沉淀总蛋白的SDS-PAGE

图4 pET-28b(+)-e 表达上清总蛋白的SDS-PAGE
2.4 pET-28b(+)-e 蛋白粗提液的溶菌试验结果
pET-28-e 细菌沉淀超声所得到的粗提液对保加利亚乳杆菌、金黄色葡萄球菌、大肠杆菌都有不同程度的溶菌作用,而且其在Tris 缓冲液的活性比PBS 缓冲液的活性强。
pET-28b(+)-e 蛋白粗提液对不同细菌的溶菌试验结果见表1。

表1 pET-28b(+)-e 蛋白粗提液对不同细菌的溶菌试验结果
3 结论
成功构建了酸奶中乳酸菌噬菌体CC34 的裂解基因 e 的表达载体:pET-28b(+)-e。pET-28b(+)-e的胞内表达,使e 的表达量有极大的提高,根据溶菌试验的证明,pET-28b(+)-e 表达的e 对多种细菌都有溶菌作用,e 的表达量大,但溶菌活性表现还不是很强,可能是pET-28b(+)-e 表达有相当一部分形成没有活性的包涵体,只有少部分以可溶性蛋白表达的原因。可通过降低诱导表达的培养温度来增加可以溶性蛋白的表达量,以增加pET-28b(+)-e 的溶菌活性[11-13]。pET-28b(+)-e 的蛋白粗提液表现出对TBS 的偏好性,在PBS 中,溶菌活性被完全抑制了。
很多噬菌体进入宿主机体的关键步骤就是通过溶菌酶打开细菌细胞壁肽聚糖的多种共价键,使肽聚糖降解,造成细菌的死亡,这种能力在人类病原菌治疗、食品、植物病害防治等方面有着很大的开发利用价值[14-15]。所研究的裂解基因e 和溶菌酶基因在理化性质上有很多相似的地方,所做的研究也为进一步了解噬菌体病毒包装机制奠定了基础。
猜你喜欢
科技视界(2022年10期)2022-05-20
中国兽医学报(2022年2期)2022-03-04
植物保护(2021年4期)2021-11-12
科学24小时(2020年4期)2020-05-14
保健与生活(2020年6期)2020-03-20
昆明医科大学报(2019年2期)2019-09-10
恋爱婚姻家庭·养生版(2019年1期)2019-01-25
中国卫生产业(2018年12期)2018-05-14
