
三江源国家公园植被净初级生产力变化趋势及影响因素
2022-08-15 07:15王军邦张秀娟芦光新王春雨张志军李英年
生态学报 2022年14期
左 婵,王军邦,张秀娟,芦光新,叶 辉,王春雨,张志军,李英年
1 长江大学园艺园林学院,荆州 434000 2 中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室,生态系统大数据与模拟中心,北京 100101 3 青海大学农牧学院,西宁 810000 4 九江学院旅游与地理学院,九江 332005 5 中科院西北高原生物研究所,西宁 810000 6 青海省生态环境监测中心,西宁 810000
青藏高原被誉为世界“第三极”,其特殊的海拔、地形地貌导致生态系统结构简单且脆弱,对全球气候和环境变化的响应十分敏感和强烈[1]。三江源地区位于青藏高原腹地,属于典型的生态系统脆弱区,是我国生物多样性保护和生态环境建设重点区域,是维系区域生态安全的重要屏障[2—3]。由于全球气候变暖和过度放牧等人类活动加剧的双重影响[4],三江源地区草地退化改变着植被格局、水碳平衡和生产力,进而可能影响区域生态安全和社会经济可持续发展[5]。随着2000年省级三江源自然保护区的建立[6],2005年三江源生态保护和建设工程的实施[7],2016年成为中国首批国家公园试点[8],2021年宣布正式成为国家公园,进一步提升了生态保护力度和体制创新。而开展该区域高寒植被净初级生产力变化及其对气候变化和人类活动的响应研究,对于理解高寒植被生产力变化机制,有效实施生态保护和促进区域可持续发展,将提供理论基础和科学决策依据。
植被净初级生产力(Net Primary Production,NPP)是植被在单位面积上所积累的有机物质的量,是植物光合作用所同化的总初级生产力减去植物自养呼吸后的剩余部分[9—10],是地球生命系统的物质和能量基础,是生态系统其他服务功能形成和维持的基础。对三江源区生产力变化已经开展了大量研究,主要集中在植被生产力估算模型和遥感数据的应用与发展[3,11—12]、全球气候变化背景下植被生产力变化[13—16]以及气候和人类活动影响的分析[17—21]等方面,针对植被变化的原因存在三种解释:(1)气候变化[22—24];(2)过度放牧等人类活动影响[25];(3)生态保护与修复等恢复措施的综合效应[26—27]。高寒植被存在较大的空间异质性,且受气候变化等的复杂综合影响而导致厘定气候变化和人类活动影响存在较大不确定性,另外也缺乏基于长时间序列数据的研究,导致生产力变化的主要驱动机制始终存在较大争议。
生态遥感耦合模型通过整合生态系统过程模型和遥感参数模型分别在生理生态模拟和空间异质性量化方面各自的优势,能够实现空间明晰地模拟和估算陆地生态系统生产力过程变量[28]。作为生态遥感耦合模型之一,GLOPEM-CEVSA耦合模型基于陆地生态系统碳循环过程及其生理生态学理论,实现了碳周转、碳固定、碳分配等碳循环过程的模拟[28—30],研究表明该模型在估算NPP中精度更高[3],在三江源地区生产力估算及草畜平衡分析等方面得到了广泛的应用[31]。
因此,本文应用GLOPEM-CEVSA模型模拟1981—2018年三江源地区植被潜在和现实净初级生产力,分析近40年植被生产力时空格局及影响因素,揭示在全球气候变化背景下人类活动的影响,以期为区域生态监测与评价和国家公园保护与发展管理决策提供理论依据和方法参考。
1 研究区概况
三江源国家公园位于青海省南部三江源地区,地理范围是33°09′—36°47′N,89°50′—95°18′E,海拔为3500—4800 m,是长江、黄河、澜沧江3条重要河流的水源发源地和我国重要的淡水补给地。园区范围包括长江源、黄河源以及澜沧江源园区,总面积12.31万km2。三江源地处青藏高原腹地,长江源园区和澜沧江源园区以冰川、高山和高平原丘陵地貌为主,黄河源园区大部分为高平原,如图1。
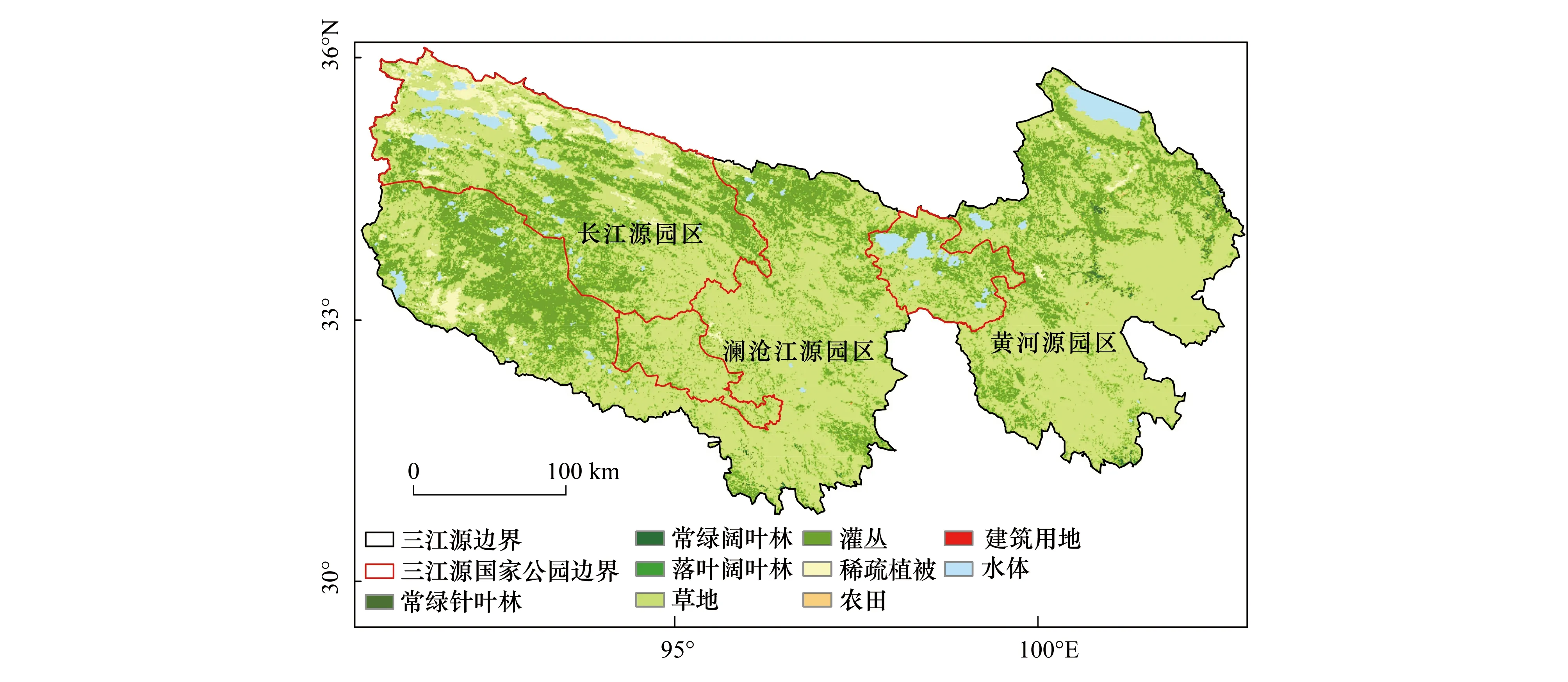
图1 三江源区地理位置及主要土地覆被类型
2 材料与方法
2.1 数据来源及处理
2.1.1气象数据
用于模型输入的1981—2018年气象数据,是利用全国753个和周边国家345个气象台站观测数据经空间内插的栅格数据[32],空间分辨率为1 km,时间分辨率为8天,包括最高、最低气温、降水量、风速、日照时数和相对湿度。空间插值采用由澳大利亚国立大学基于光滑薄板样条算法的插值软件ANUSPLIN[33],研究表明ANUSPLINE插值的数据分别能解释94%和77%的气温和降水空间变异,明显优于其他方法[34]。插值中主要考虑了经纬度和海拔高度对各气候要素的影响,其中海拔数据采用了90 m空间分辨率SRTM(Shuttle Radar Topography Mission)的数字高程模型(DEM)数据[35],经重采样为1 km空间分辨率作为插值辅助数据。
2.1.2基于卫星遥感参数反演的FPAR数据
基于卫星遥感参数反演的FPAR,是经GIMMIS3g和MODIS数据融合得到的1981—2018年1 km空间分辨率每8天时间步长的数据产品。GIMMIS3g数据提供了1981—2015年每半月1/12°空间分辨率归一化植被指数数据(NDVI3g)[36];最新MODIS的FPAR产品,提供了2003年以来每8天1 km空间分辨率数据产品(MCD15A2.C006)。为了提高遥感数据的时空连续性和可用性,以2003—2015年两产品重叠期间数据,基于人工神经网络算法重构了1981—2002年每8天1 km空间分辨率FPAR数据,而2003—2018年每8天1 km空间分辨率数据直接采用了MCD15A2产品,最终得到了1981—2018年每8天1 km空间分辨率数据产品,从中裁切出三江源地区,用于本研究。
2.1.3植被覆盖类型数据
本文采用的植被覆盖类型图是基于如下三种数据源综合而成:A.刘勇洪等[37]以MODIS数据分类结果,该分类利用2001—2002年1 km的MODIS NDVI时间序列数据和多波段反射率光谱数据对中国区域进行了土地覆盖分类;B.基于30 m分辨率Landsat遥感数据的青海地区2000年土地覆盖图(三江源项目组数据);C.基于30 m分辨率Landsat遥感数据的三江源地区2004年土地覆盖图(三江源项目组数据)。本研究数据融合中土地覆盖大类(森林、灌丛、草地、农田、水体、建设用地)以C为主,在细化森林类型的时候采用了A数据源,并在ArcGIS中采用众数聚合法进行空间重采样为1 km空间分辨率,用于模型输入。植被覆盖类型中生态系统类型包括常绿针叶林、常绿阔叶林、落叶针叶林、落叶阔叶林、灌丛、草地、稀疏植被、农田、建设用地和水域。
2.1.4土壤质地数据
土壤质地数据来自刘明亮全国土壤质地数据[38],用于土壤水分参数的计算。该数据是以中国资源环境数据库1∶400万土壤类型图做空间插值,选取了第二次土壤普查共516个剖面点,对每一个一级类别所覆盖的土壤剖面点计算粒级机构的平均值,作为此土壤类型的粒级构成。本研究在ArcGIS中对数据经邻近值法重采样处理得到与气象数据的空间分辨率相匹配的1 km数据,并裁剪出三江源地区数据用于模型输入。
2.1.5模型验证数据
采用本省曲麻莱县生态气象站(95°48′00″E,34°07′00″N,4175.0 m)和门源县海北站生态气象站(100°51′35″E,36°57′33″N,3140.0 m)长期监测牧草产量数据用于模型结果验证。牧草产量监测取样,是在气象站周围10 km范围内平坦开阔且面积为100 m×100 m的草地做为采样区,在采样区内随机选取5个样方(1 m×1 m),从5至8月每月测量并详细记录样方内牧草的高度和覆盖度,并以收获法测定鲜重,以牧草生长期内(5—8月)最大值作为该年度牧草产量;采样区仅在冬季进行放牧,春、夏、秋季进行封育[39]。
基于涡度相关理论的通量观测数据,可以拆分出总初级生产力,可间接验证模型估算的净初级生产力。鉴于只能获得中国通量观测网络共享的位于三江源区东缘的海北灌丛2003—2010年通量观测数据,因此,通过该通量观测数据,间接评价模型对NPP的估算性能。
2.2 人类活动影响模拟
参照许端阳等[40],人类活动对植被变化的影响,以气候潜在植被生产力与卫星遥感的现实植被生长力之差估算:
NPPHA= NPPRS-NPPCL
(1)
式中,NPPHA为人类活动导致的净初级生产力;NPPRS是以卫星遥感的FPARRS进行模拟得到的现实生产力;NPPCL是以气候驱动模拟的FPARCL计算的潜在生产力,可分别表示为:
NPPRS= PAR×FPARRS×ε*×σ-Ra
(2)
NPPCL= PAR×FPARCL×ε*×σ-Ra
(3)
式中,PAR是光合有效辐射;ε是基于GPP概念的植被光能利用率;Ra为植被自养呼吸;σ为基于气孔导度类比模型的环境胁迫因子,在理想环境条件下,植被存在潜在最大光能利用率(ε*),现实光能利用率则受环境因子胁迫(σ),ε*和σ的计算详见文献[41—43]。
FPARRS直接采用卫星遥感反演的FPAR数据,FPARCL采用下式计算[44]:
FPARCL=1-e-kLAI
(4)
LAI=LAIMIN+σ×(LAIMAX-LAIMIN)
(5)
式中,k为消光系数,取值0.5,LAIMIN和LAIMAX是逐像元的最小和最大叶面积指数;σ的解释见式(3);GLOPEM-CEVSA模型的详细介绍见文献[41—43]。
2.3 研究方法
2.3.1人类活动影响分析
三江源地区人类活动影响程度,以人类活动影响的生产力NPPHA与潜在生产力NPPCL之比HC量化;比值小于0区域人类活动呈负面影响,比值大于0区域人类活动呈正面影响,且值越大影响程度越大。
(7)
以现实和潜在净初级生产力的年际变化趋势之差,量化人类活动对净初级生产力年际变化趋势产生的影响,即:
SlopeHA= SlopeRS-SlopeCL
(8)
式中,若SlopeHA大于0,表明人类活动影响为促进作用;若小于0,人类活动影响则为负面影响。以实际人类活动因素对净初级生产力的变化幅度产生的影响SlopeHA与气候驱动的潜在净初级生产力(NPPCL)的年际变化SlopeCL比值的绝对值(│SlopeHA/SlopeCL│)判断NPP趋势变化的主导因子,当绝对值大于1则表示人类活动影响为NPP趋势变化的主导因子,小于1则表示气候变化为主导因子。
2.3.2气候因子影响分析
采用多元线性回归方法分析影响净初级生产力变化的气候因素,回归方程可表示为:
Y=A0+A1X1+A2X2+A3X3
(9)
式中,Y是净初级生产力时间序列,X1、X2、X3分别为年累计降水量、年平均日最高气温和年平均日最低气温时间序列;A0、A1、A2、A3为回归系数,当自变量经标准化处理后,通过回归系数可比较自变量对因变量的相对贡献大小。
3 结果与分析
3.1 模型验证
采用曲麻莱和海北站牧气站长期监测牧草产量数据,对模型估算的实际净初级生产力进行验证时,模型估算的现实NPP与牧草产量显著线性相关(P<0.01),如图2所示;由复相关系数可知,模型能够分别解释两地牧草产量年际变化的59%和67%,全部牧草产量年际变化的77%;值得说明的是,由于牧草产量数据实质上仅仅是地上生产力,因此,二者的回归系数,即斜率并不会等于1。如图2所示,基于通量观测数据的验证结果表明,模拟与观测间显著线性相关(P<0.01),模型能够解释通量塔观测GPP季节变化的89%,说明模型对高寒草地生产力具有较高的估算精度。
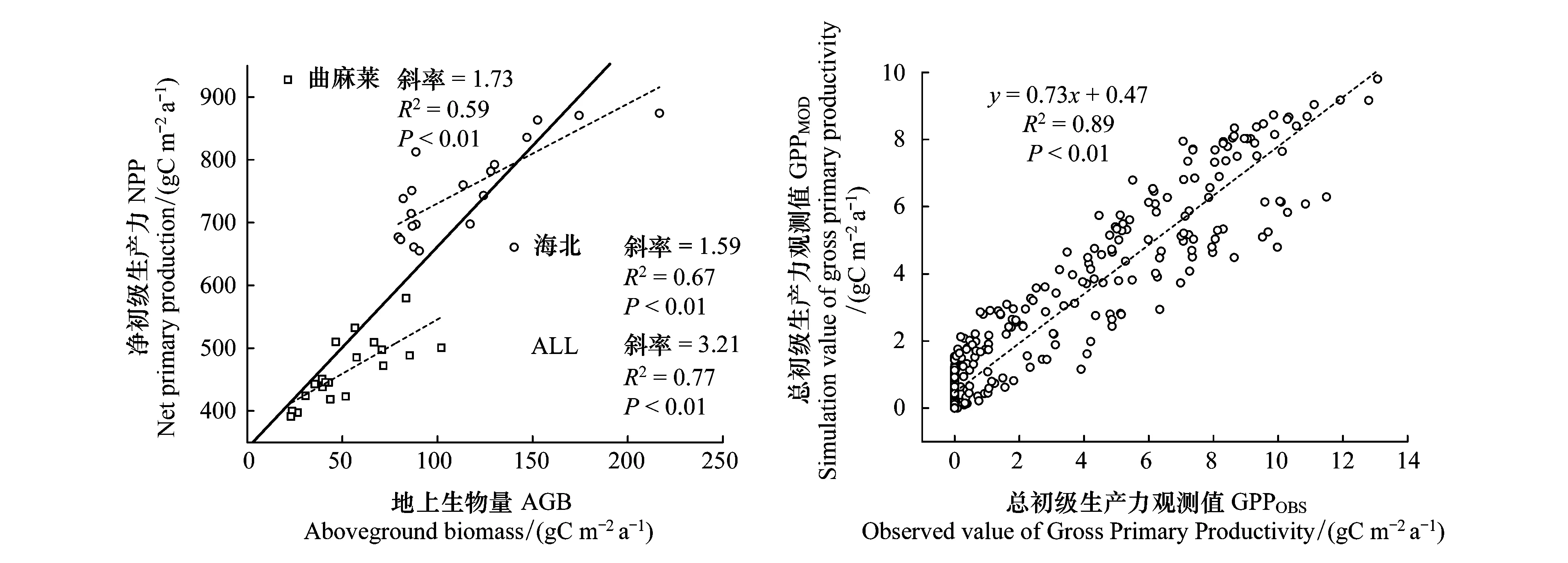
图2 净初级生产力模拟值与站点监测地上生物量数据以及海北通量站模型模拟总初级生产力值与观测数据的对比
3.2 净初级生产力空间格局
三江源地区NPP存在明显的从东南向西北逐渐递减的空间分布格局,如图3所示。近40年,三江源全区现实NPP(NPPRS)多年均值为(309.70±225.77)g C m-2a-1,占潜在NPP(NPPCL)的61.65%,如图4所示。其中,黄河源、长江源和澜沧江源NPPRS分别为(249.88±122.65)g C m-2a-1、(140.18±123.72)g C m-2a-1和(330.55±146.71)g C m-2a-1,分别占各园区NPPCL的64.98%、62.85%和61.65%;则NPPHA分别为(-133.85±173.59)g C m-2a-1、(-82.36±155.02)g C m-2a-1和(-205.23±189.48)g C m-2a-1。
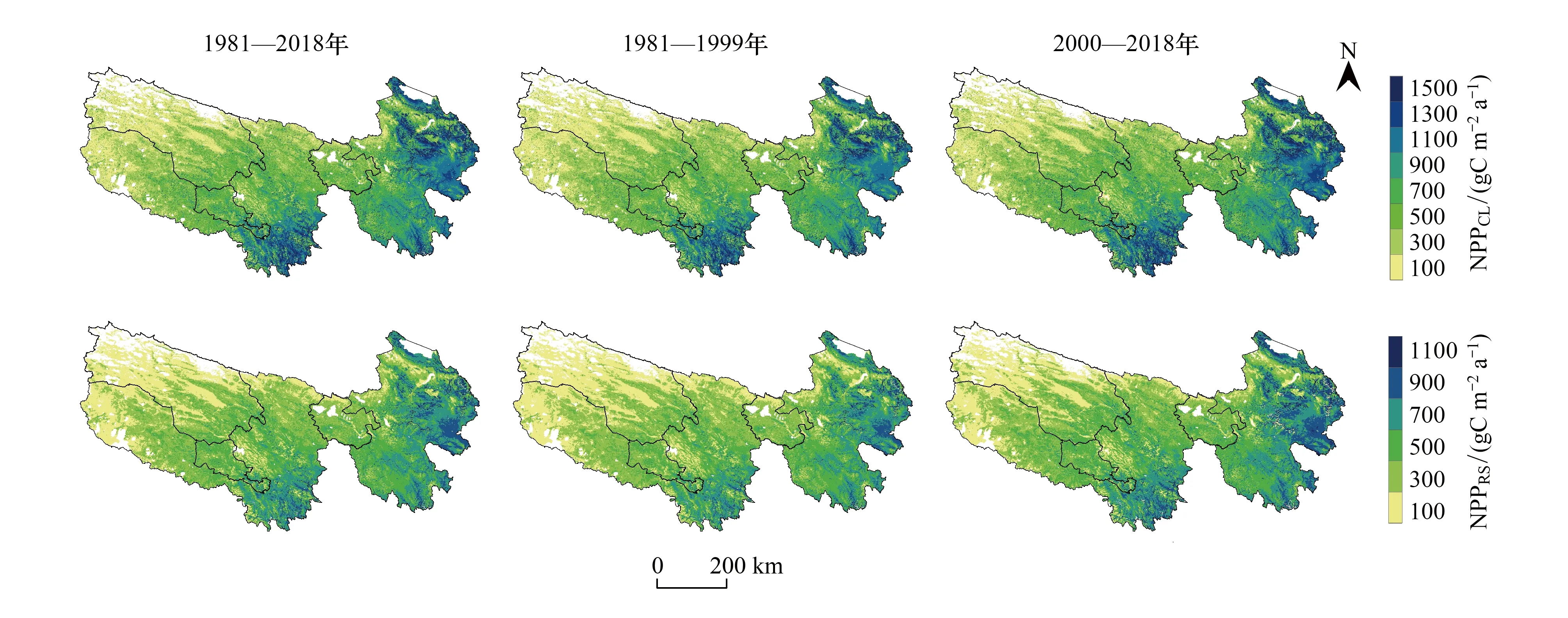
图3 三江源地区1981—2018年及2000年前后多年平均NPPCL和NPPRS的空间分布格局
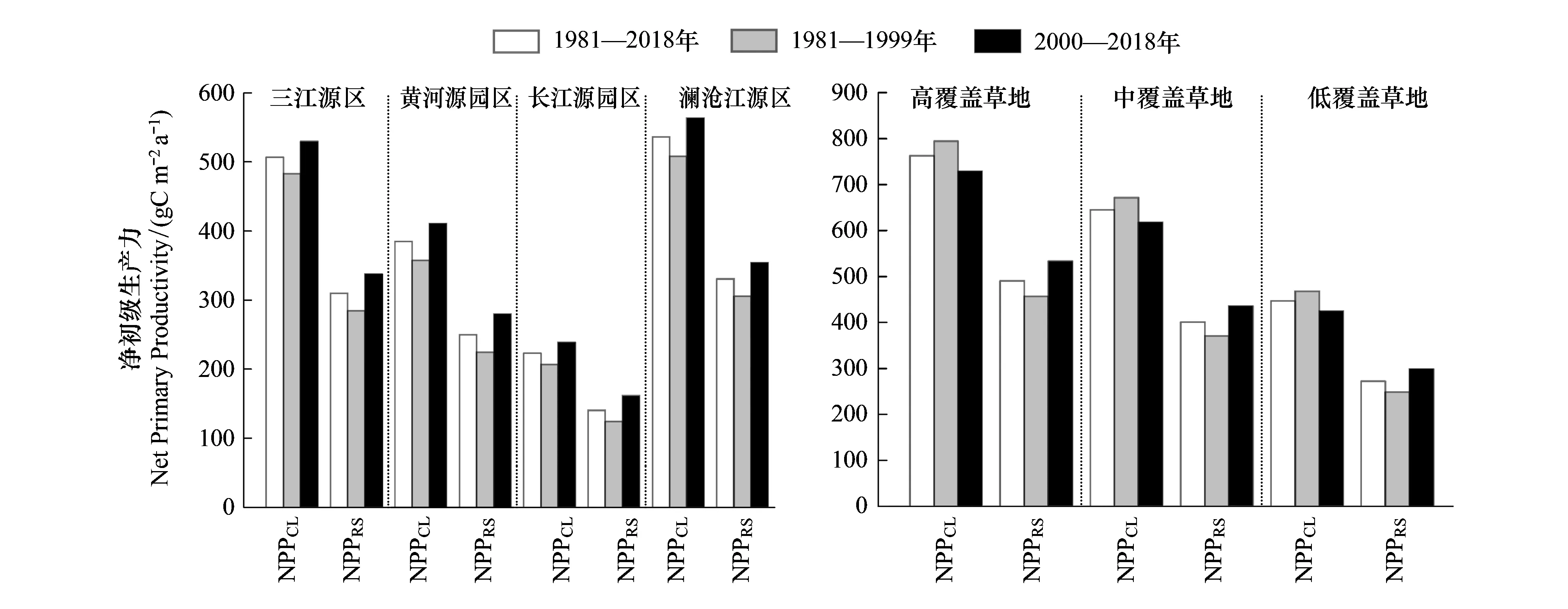
图4 三江源全区及不同园区净初级生产力区域多年平均值及不同草地类型净初级生产力区域多年平均值
三江源地区分布面积最广的草地,如图4,NPPRS多年均值为(336.74±218.02)g C m-2a-1;根据2010年全国LUCC数据所给出的高、中、低覆盖草地,其NPPRS多年均值分别为(489.97±231.05)g C m-2a-1、(399.85±213.10)g C m-2a-1和(271.96±191.04)g C m-2a-1,分别占对应覆盖度草地NPPCL的64.27%、62.03%和60.93%;则NPPHA分别为(-270.51±213.38)g C m-2a-1、(-244.49±221.47)g C m-2a-1和(-173.76±211.63)g C m-2a-1。
3.3 净初级生产力年际变化趋势
1981—2018年三江源全区NPPRS和NPPCL整体均呈显著增加趋势,NPPRS以每年2.00 g C m-2a-1的速率显著增加,大于仅气候驱动下的增加速率(1.74 g C m-2a-2),如图5和表1所示。2000年后NPPRS增速是NPPCL的2倍,而2000年前,前者仅为后者的59.16%。较之2000年前,2000年后NPPRS的增速是2000年前的1.5倍,而NPPCL增速则仅为前期的40.08%。结果明显表明现实生产力增加趋势在显著加强。
2000年前后年际趋势的空间分布存在较大差异,如图5所示。2000年前NPPCL在三江源东南部呈显著下降趋势,东部呈显著增加趋势,2000年后呈下降趋势区域则为三江源东部一带及澜沧江园区东部等,而黄河源南部一带为增加趋势。NPPRS近20年三江源东北部环青海湖一带呈显著增加趋势,而下降区域由三江源中部转移至东部、长江源园区东部和澜沧江源园区大部;下降趋势区域相较气候驱动下呈明显减缓状态。
近40年NPPRS增速最大的为黄河源园区(2.38 g C m-2a-1),最小为长江源园区(1.20 g C m-2a-1),黄河源、长江源和澜沧江源园区的NPPRS增长速率占各园区NPPCL增长速率分别为89.13%、90.23%和77.43%,如图6和表1所示。2000年前后相比,黄河源园区NPPCL变化不明显,而近20年NPPRS增长速率增加了4.8倍,变化最大;长江源园区近20年NPPRS是2000年前的2.32倍,变化趋势显著增强;澜沧江源园区变化趋势受气候影响较大,NPPCL增速下降超一倍,而NPPRS在2000年后仅增速12%。
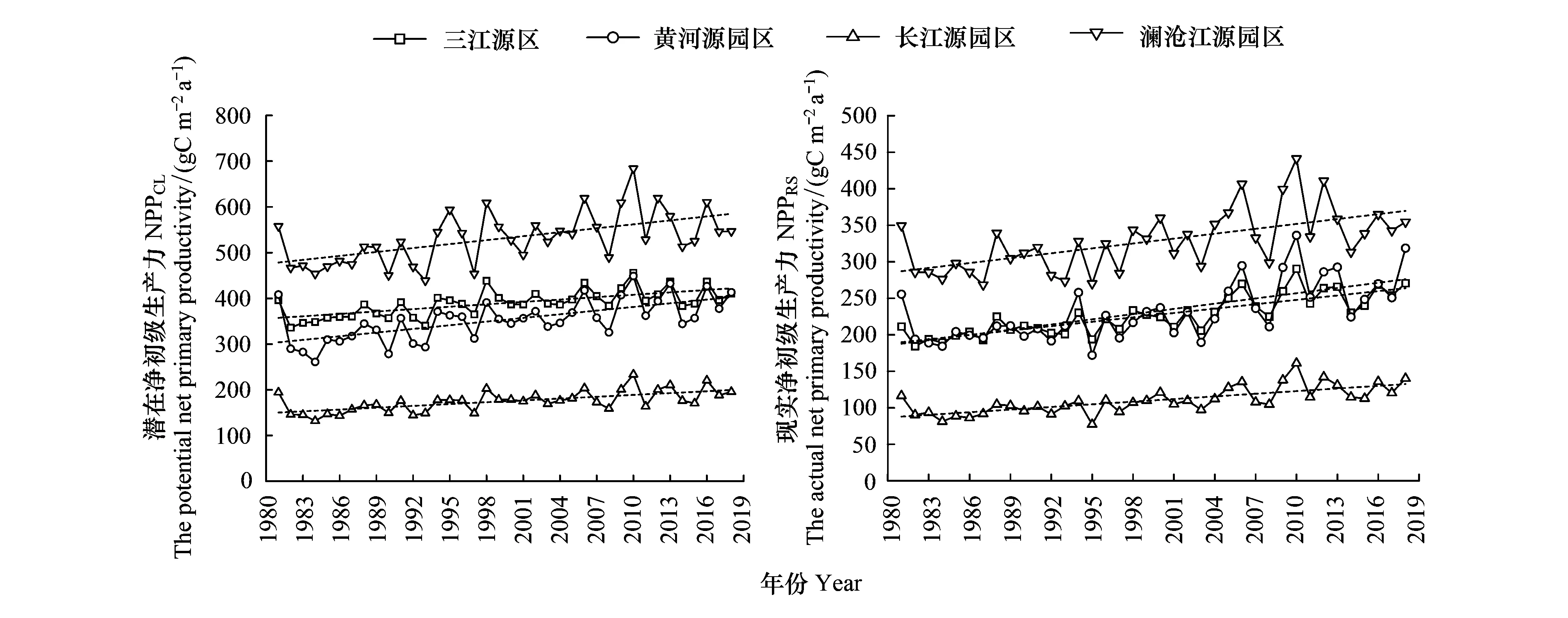
图6 1981—2018年三江源全区及不同园区植被净初级生产力年际变化趋势

表1 1981—2018年三江源全区及不同园区净初级生产力前后19年变化趋势
植被类型中,近40年草地NPPRS和NPPCL均呈显著增加趋势,现实生产力增速(2.10 g C m-2a-1,R2=0.67,P<0.01)较潜在生产力增速(1.84 g C m-2a-1,R2=0.43,P<0.01)高14%,如图7所示。其中,高、中、低覆盖度草地NPPRS在2000年后变化趋势是2000年前的1.30、1.48和1.52倍;NPPCL增加趋势均呈趋缓变化,其中高、中、低覆盖度草地变化趋势2000年后较2000年前分别下降了41.69%、56.91%和64.75%。
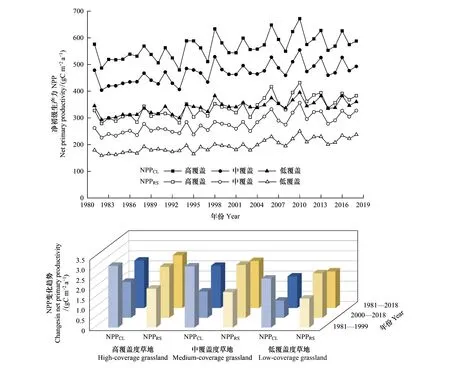
图7 不同草地覆盖类型净初级生产力年际变化趋势对比
3.4 气候变化影响
以1981—2018年年累计降水、年平均日最高气温和年平均日最低气温为自变量,以生产力为因变量的多元线性回归方程,可分别解释三江源区NPPCL和NPPRS年际变化的51%和73%,可分别解释黄河源、长江源和澜沧江源园区NPPCL的48%、52%和42%,NPPRS的58%、69%和50%,如表2所示。
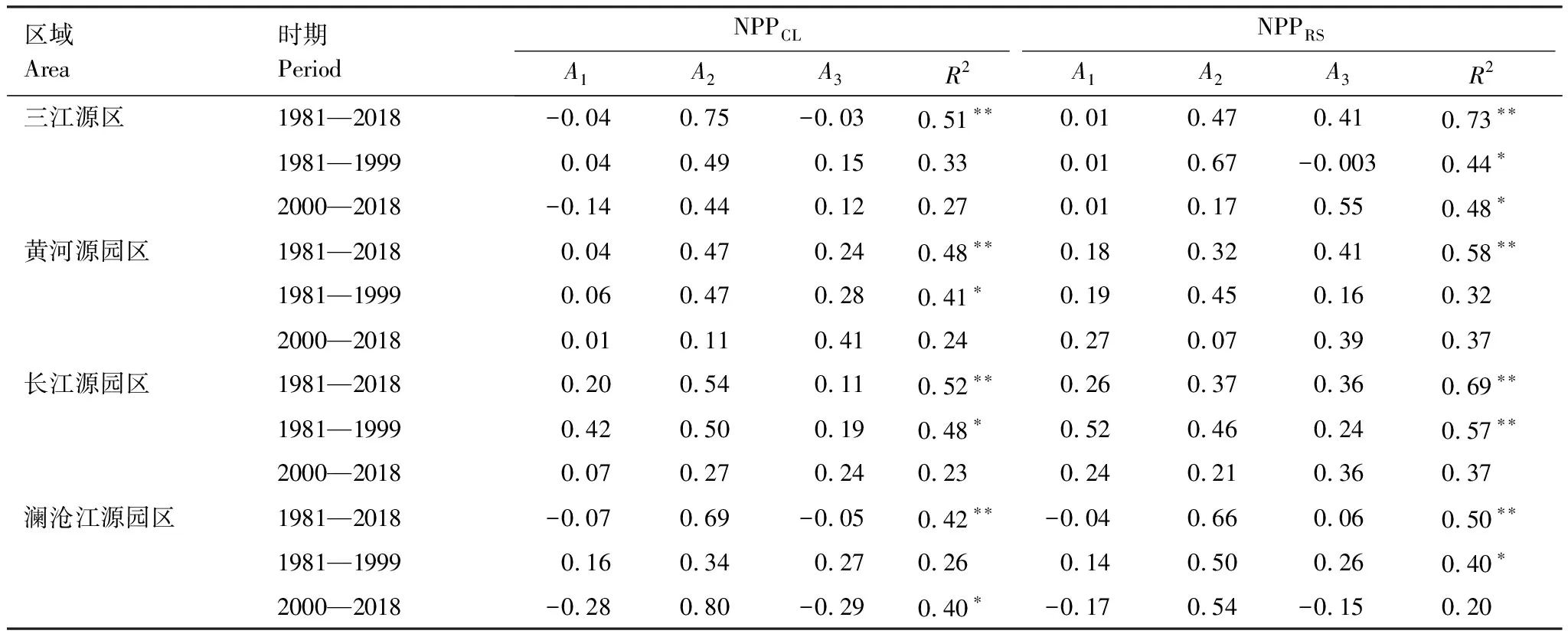
表2 三江源全区及不同园区标准化的净初级生产力与气候因子间多元线性回归分析
近40年三江源地区NPPRS主要受气温的影响,根据标准化回归系数,最高气温(0.47)和最低气温(0.41)的贡献分别是降水(0.01)贡献的65.6倍和57.6倍;NPPCL主要受最高气温影响(0.75),最低气温影响较小(-0.03),二者的贡献分别是降水贡献(-0.04)的18.4倍和0.7倍。就三个园区而言,NPPRS在黄河源园区最高和最低气温的贡献是降水贡献的1.8倍和2.3倍,表明受最低气温影响较大;长江源园区对应贡献比都是1.4倍,表明受最高和最低气温共同影响;澜沧江源园区对应贡献比为15.6倍和1.4倍,受最高气温作用较大。NPPCL在三个园区均受最高气温影响最大,与降水的贡献比分别为11.6倍(黄河源)、2.7倍(长江源)和9.5倍(澜沧江源),而最低气温与降水的贡献比依次分别为5.9倍、0.6倍和0.7倍。
三江源区NPPRS尽管在近40年期间最高气温是其主控气候因子,但在2000年前后的主导气候因子出现不同。2000年前最高气温和最低气温分别与降水的贡献比为66.5倍和0.3倍,最高气温是主导因子;而2000年后贡献比分别为15.8倍和52.3倍,最低气温成为了主导气候因子。在不同园区间相比,2000年前最高气温与降水的贡献比为黄河源2.3倍,长江源0.9倍和澜沧江源3.4倍,而2000年分别降低至0.3倍、0.9倍和3.2倍;相反最低气温的贡献比,黄河源从0.8倍增加到了1.4倍,长江源从0.5倍增加到了1.5倍。而澜沧江源区变化较为特别,整个时段内气温对生产力年际变化为正影响,但在2000年后最高气温成为负影响,与降水的贡献比,从2000年前的3.4倍变为了2000年后的-3.2倍;然而最高气温的绝对影响仍然最高。园区NPPCL受最高气温的显著影响,2000年以后NPP受气候变化影响不显著。
3.5 人类活动影响
基于现实生产力NPPRS与气候潜在生产力NPPCL之差的人类活动影响的生产力(NPPHA),近40年三江源全区平均为-196.95 g Cm-2a-1,是气候潜在生产力NPPCL的38.9%。如图8所示,人类影响总体为负,但空间分布存在较大差异,东北部的海南州和中南部的囊谦地区为较大的负影响;北部黄河源及西北部可可西里和唐古拉地区为正影响,正影响区域面积仅占全区植被总面积的13.84%。
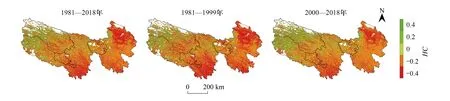
图8 三江源地区人类活动对净初级生产力的影响程度的空间格局分布
2000年前后NPPHA分别为-198.36 g C m-2a-1和-192.04 g C m-2a-1,分别占潜在生产力NPPCL的41.1%和36.3%,人类活动影响降低了4.8%。对于黄河源、长江源和澜沧江源三个园区,人类活动影响分别降低了5.2%、7.5%和2.8%,其中,以长江源的降低幅度最大。
三江源地区NPPHA的变化趋势如图9所示,近40年,全区NPPHA变化趋势为0.26 g C m-2a-1,是潜在NPPCL趋势的14.75%,即人类活动对全区NPP为正影响,对趋势的贡献为14.75%。其中,长江、澜沧江和黄河源园区NPPHA的变化趋势分别为-0.13 g C m-2a-1、-0.65 g C m-2a-1和-0.29 g C m-2a-1,分别占NPPCL趋势的-9.59%、-22.42%和-10.72%,表明在三个园区人类活动为负影响,以长江源园区受人类活动负影响最小,澜沧江园区负影响最大。

图9 三江源地区人类活动对净初级生产力变化趋势的影响
2000年前后NPPHA的变化趋势分别为-1.07 g C m-2a-1和1.27 g C m-2a-1,即2000年后NPPHA年际变化转变为趋势增加,人类活动对NPP变化趋势的影响由负逐渐转变正向促进作用。趋势斜率为负的区域面积占比从2000年前的79.12%,降低到2000年后的56.34%。其中显著为负的地区,2000年前主要分布在三江源东北部和中部、长江源区东南部以及澜沧江园区及周边大片区域,2000年后主要分布在长江源园区东部、澜沧江源园区大部以及黄河源园区南部。NPP趋势变化中人类活动占主导的面积占比由18.73%明显增加至38.76%,正向贡献的范围逐渐在扩大。
2000年后NPP趋势变化中人类活动在长江和黄河源园区为正向贡献,而在澜沧江源为负影响。具体地,长江、澜沧江和黄河源园区的NPPHA变化趋势分别为0.06 g C m-2a-1、-0.83 g C m-2a-1和0.76 g C m-2a-1,是NPPCL变化趋势的5.03%、-44.25%和27.45%。而在2000年前人类活动对三个园区NPP趋势均为负影响,NPPHA趋势是NPPCL趋势的-64.29%、-76.36%和-76.61%。后期人类活动影响增强,较前期增加了69.32%、32.11%和104.055%,黄河源园区人类活动对NPP变化趋势的影响变化最显著,而澜沧江园区人类活动对NPP变化趋势呈长期负影响,但影响逐渐减弱。
4 讨论
4.1 模型不确定
作为典型生态脆弱区的三江源,其植被生产力状况、变化及驱动机制研究,一直倍受诸多关注,不同模型、方法和输入数据间存在差异,而导致结果不确定性较大。对该地区以往研究文献数据与本研究对比表明,郭佩佩等[16]采用Thornthwaite Memorial模型估计三江源全区NPP为570.35 g C m-2a-1,蔡雨恋等[45]对比研究了Miami模型和周广胜模型模拟结果,发现2004—2008年NPP多年均值分别为486.90 g C m-2a-1和302.45 g C m-2a-1,而本研究的模拟的气候潜在NPP为506.65 g C m-2a-1,在前人采用气候模型估算的数值范围内。由于诸如Thornthwaite Memorial等气候模型,建立在有限观测数据的经验统计模型基础上,同时所考虑的生态过程相对有限,而导致结果存在较大不确定性。而本研究所发展的模型,其中气候模块类比气孔导度构建了叶面积指数模拟模型,进而以Beer定律估算FPAR,考虑了土壤水分过程及气温对植被的胁迫作用,而应该更为合理。
本研究估算近40年三江源全区现实NPP(NPPRS)多年均值为309.70 g C m-2a-1,高于刘凤等[17]、蔡雨恋等[45]、张颖等[46]和沃笑等[47]采用CASA遥感模型估算结果,而显著低于贺倩等[15]采用CASA模型研究2010年和2015年三江源植被NPP均值397.43 g C m-2a-1和345.44 g C m-2a-1。由此可见相同模型研究结果存在显著差异,模型参数和输入数据的不确定性同样影响着模型模拟结果的精度。本研究的估算值与王军邦等[31]采用相同模型模拟的1988—2004年三江源地区多年平均NPP值相比高出近一倍,这可能与研究时段及采用的数据不同有关,并且本研究所采用的是经参数优化的模型,也对模型输入FPAR产品进行了改进,估算值应该更加符合三江源地区植被生产力状况。
但目前的模型参数优化,仍然建立在数量有限的通量站点观测基础上,这些站点难以代表面积较广空间异质性较高的相同植被类型内部的差异,因此,今后需要进一步收集地面观测数据,利用多尺度卫星遥感数据,构建地面基准数据,然后继续逐像元的模型参数优化,才有可能进一步降低模型的不确定性。本研究利用同一个模型框架和同一套输入数据,估算潜在和现实净初级生产力,一定程度上可消除由模型结构和输入数据不同而带来的不确定性,因此,研究结果更具可信。而基于像元尺度的模型参数优化,应该能进一步提高模型估算精度,因此需在今后研究中予以关注[48]。
4.2 植被变化的主导因子
三江源地区植被净初级生产力变化的主导气候因子还存在着极大的争议。部分研究,如孙庆龄等[49]、李辉霞等[50]、Xu等[51]、才吉[52]和彭凯峰等[53],均认为降水对三江源地区净初级生产力的影响大于气温,NPP的变化趋势主要与水分条件有关。王军邦等[32]、蔡雨恋等[45]、张妹婷等[54]和许洁等[55]研究,则认为气温的变化才是影响三江源地区NPP变化的主要气候因素,本研究结果与后一观点一致。近40年三江源地区NPP主要受气温的影响,其中潜在NPP的主导因素是最高气温,现实NPP主要受最低气温和最高气温的共同影响,与降水的相关性不高或呈限制作用,这也与周秉荣等[39]结论一致。
本文结果表明不同时段存在不同的气温影响机制。在全球变暖的背景下,高海拔的寒冷地带变缓效应更加明显,三江源地区气候整体上呈暖湿化趋势,降水和温度对植被生产力起着至关重要的作用[14,56—57]。以往大部分研究认为三江源的气候变化越来越适宜于牧草的生长[14,58],而本研究认为,尽管从近40年的时间尺度上三江源气候变化表现出有利于植被生产,但进入21世纪,气候变化趋势相较20世纪下降明显,气候变化对植被生产力促进作用逐渐降低,与张颖等[57]研究结论一致。李猛等[20]研究三江源草地ANPP与气候因子之间的关系,认为气温的升高将有利于草地植被的生产,而未来降水的变化将会因温度条件的差异而产生不同的效应,从而可能导致三江源草地植被生产变化在区域上的差异。Raich等[59]和Wang等[60]认为在高海拔地区,低温更容易形成水分或养分胁迫,限制植物生产,当温度达到一定阈值,胁迫作用会逐步缓解,植物生产力也会提高。结合表1和表2,三江源地区总体上随气温增高而植被生产力增加,但存在区域差异,比较明显的是长江源前期降水的影响较大,而后期影响减小;澜沧江后期出现受降水和最低温度的抑制。
值得深思的是,气候变暖在促进植被生长的同时也将加速冰川融化,从长远而言,有可能导致该地区供水不足,进而限制植被生长,导致生态环境进一步恶化。另外,气候变化不仅直接影响植被生存,也影响植被群落结构变化,如引起物种的生态位分布改变[61],但目前研究中尚未考虑气候变化对植物群落的影响。因此,今后研究中不仅需考虑植被功能和群落对气候变化的响应,也需考虑这些变化对气候的反馈作用,以更全面理解全球气候变化背景下生态系统的变化。
4.3 生态保护工程对植被变化的贡献
三江源地区自2000年设立自然保护区,2005年实施生态保护与建设工程,学者们对此进行了大量的生态工程成效评估。邵全琴等[62—63]研究指出生态保护工程缺乏长期效应,人类活动在2010年后积极效应下降,认为环境治理的速度跟不上生态恶化的速度,仍存在着诸多问题。李作伟等[64]研究1982—2013年气候条件对植被生产力的平均贡献率为87%,而人类活动的贡献率为13%,认为进入21世纪以来人类活动正面影响较为明显,在一定程度上加快了其变化速率;张颖等[57]研究认为人类活动对草地植被的负面影响力有明显减弱。本研究也表明人类活动对三江源地区植被的负面影响力在程度和范围上逐渐减弱,促进了植被生产力的增加。
三江源地区受气候变化及过度放牧等人类活动的共同影响,草地退化、土壤沙化等一系列问题的产生及加剧导致植被生产力的增加趋势变缓,而2000年以后生态保护与建设工程、草地禁牧减畜管理、人工补播草种、鼠害治理等生态恢复措施的实行促使植被压力减弱,生态恢复效果显著,生产力呈显著的增长趋势[65]。然而,王军邦等[66]采用CEVSA-RS模型模拟2000—2017年以来全国陆地生态系统总初级生产力,也发现青藏高原人类活动影响整体上仍呈负面作用,对草地影响严重。尽管生态保护工程在一定程度上恢复了植被覆盖度,促进了部分地区的生产力变化趋势,但并未平衡过度放牧所引起的植被生产力下降等问题,部分地区出现效应反弹现象,人类活动影响可能有所加剧[67]。
生态保护和建设工程等人类活动并未根本性遏制草地退化问题[68],本研究表明2000年以后生态保护与建设工程、禁牧管理、人工草地种植等生态恢复措施的实行促进生产力增长加速,这些生态措施有所减缓了人类活动的负面影响。然而,在气候占主导作用的这一生态脆弱区,未来仍需进一步加大生态环境保护和建设力度,完善生态保护体系,实现区域生态系统的健康稳定。
5 结论
研究利用GLOPEM-CEVSA耦合模型,分别以气象数据和气象及遥感反演的FPAR数据作为输入,实现了1981—2018年三江地区仅气候驱动的潜在植被净初级生产力(NPPCL)和气候遥感共同驱动的现实植被净初级生产力(NPPRS)的模拟估算。分析结果表明,全区植被现实NPP多年均值为309.70 g C m-2a-1,是植被潜在NPP的61.65%;其中,黄河源、长江源和澜沧江源园区分别为64.98%、62.85%和61.65%,澜沧江源园区受人类活动影响程度最大。近40年受气温变化的主导性影响,全区NPP整体呈上升趋势;而人类活动影响呈促进生产力增加趋势,但作用不显著。2000年前后人类活动对NPP的负影响程度和范围都在降低,2000年以后生态保护与建设工程、禁牧管理、人工草地种植等生态恢复措施的实行促使生产力变化趋势的增长。今后需进一步加大生态环境保护和建设力度,持续开展生态保护与建设工程;同时在研究中,不仅需考虑植被功能和群落对气候变化的响应,也需考虑植被变化对气候的反馈作用,以更全面理解全球气候变化背景下生态系统的变化。
猜你喜欢
青海草业(2022年2期)2022-07-23
小哥白尼(野生动物)(2022年3期)2022-06-16
长江科学院院报(2022年5期)2022-05-19
北京大学学报(自然科学版)(2022年2期)2022-04-08
现代艺术(2022年1期)2022-02-07
民族音乐(2018年1期)2018-04-18
红土地(2016年3期)2017-01-15
环球市场信息导报(2015年15期)2015-12-03
海峡姐妹(2015年9期)2015-02-27
公务员文萃(2013年5期)2013-03-11
