
云南干热河谷区设施葡萄与蔬菜间作的土壤培肥效果
2022-08-15 02:52张永慧李贝贝徐权杰吴流通常换换苏友波
中国土壤与肥料 2022年6期
张永慧,李贝贝,徐权杰,吴流通,常换换,苏友波*
(1.云南农业大学资源与环境学院,云南 昆明 650210;2.康乐县农业农村局,云南 康乐 631500)
全球陆地生态系统中干热河谷地区占三分之一,被认为是全世界环境最恶劣的地区之一,生态环境极端脆弱,极端高温频发、降雨少、蒸发量大、土壤贫瘠、易水土流失和植被退化[1],严重影响干热河谷地区土地可持续利用和粮食安全,威胁人类健康;我国干热河谷地区主要在云南省、台湾省、海南省和四川省,其中在云南地区主要分布于元江、怒江、金沙江和澜沧江流域的河谷地带,占据重要的生态区位。刘光华[2]通过元谋干热河谷地区罗望子人工林改善土壤物理性状,水土流失减轻;周扬[3]研究发现巨菌草可作为攀西干热河谷地区植被恢复的先锋植物,对土壤理化性质具有积极显著的效应;何白云[4]研究表明不同土地利用方式土壤细菌群落结构主要受土壤水分、土壤容重、粉粒、C/N的影响。云南省宾川县地处云岭横断山脉边缘、金沙江南岸、云贵高原西南部,为典型的干热河谷地区。研究设施葡萄间作蔬菜对土壤理化性状的影响,对干热河谷地区葡萄产业的发展和当地生态修复具有科学指导意义。
云南宾川县位于我国西南边陲,气候条件优越,其设施葡萄种植面积达1.08万hm2(2019年),产值达38.29亿元[5],种植设施葡萄成为推动区域经济发展和脱贫攻坚的重要手段。葡萄(Vitis viniferaL.)是葡萄科葡萄属木质藤本植物,在世界各地广泛种植,我国葡萄总产量据世界首位[6]。当前葡萄的种植存在结构单一、土地利用率低、大水大肥灌溉、养分过剩等问题,导致土壤养分失衡、土壤环境恶化、土壤板结、耕地质量下降[7],严重制约葡萄产业的发展。间作是常见的经济林经营方法,在恢复生态平衡、提高土地利用率,协调农林经济矛盾和增加经济效益方面成效显著[8],在果树行间种植大豆[9]、小麦[10]、决明子[11],可达到充分利用自然资源及长短期收益互补的目的,且充分利用自然资源和促进资源协同最大化。有研究表明,葡萄果园内生草可保持水土、改善土壤理化性质、增加有机质等[12-13];利用葡萄采收期后,在行间种植草莓[14]、冬马铃薯[15]、甘蓝[16]可以提高土地利用率,增加经济收入。然而设施葡萄间作蔬菜对土壤理化性质、土壤酶活性及微生物多样性有何影响尚需进一步探究。
本研究通过连续两年的田间小区试验,研究了葡萄间作不同蔬菜对根区土壤理化性质和微生物群落功能多样性以及酶活性的影响,以评估不同间作蔬菜品种对根区土壤肥力的影响,以期为葡萄种植地区农林间作的系统科学调控提供依据。
1 材料与方法
1.1 研究区域概况
研究区位于云南省干热河谷区宾川县,属亚热带低纬度高原季风气候区,年均太阳辐射量6479 MJ·m-2,年平均气温21.9℃,全年无霜期,年均日照时数2719.4 h。年均降水量559.4 mm,为云南省年降水量最低的地方。该区土壤主要为砂壤土,土层深厚,特别适宜葡萄种植。
试验在宾川县太和村(25°53′43.98″N,100°36′13.25″E)进行,海拔1464.9 m,选择6年生大棚葡萄种植园为试验对象。葡萄品种为红提,间作蔬菜品种选择适宜宾川县种植的7种蔬菜(萝卜、青笋、蚕豆、冰菜、蒜、辣椒、白菜),间作系统内蔬菜与葡萄树干基部距离50 cm,间作处理为葡萄//萝卜(GR)、葡萄//青笋(GG)、葡萄//蚕豆(GB)、葡萄//冰菜(GI)、葡萄//蒜(GGa)、葡萄//白菜(GC),葡萄单作区域作为对照(CK),小区周围设保护行,各区田间管理措施相同,小区面积为100 m2,每种处理设置3个重复,共设24个小区。葡萄“Y”字形整型,株行距为1 m×3 m。土壤的碱解氮85.5 mg·kg-1、有效磷17.7 mg·kg-1、速效钾130.0 mg·kg-1、有机质18.8 g·kg-1、pH值6.13。
1.2 样品采集
时间为2019~2020年,每年9月8日播种,11月8日采样,所有蔬菜同时播种,连续种植两年。每10 d灌溉1次,种植蔬菜后无施肥。每个小区随机选取3棵具有代表性的植株,采集蔬菜根际土壤0~20和20~40 cm深处土壤样品,采用“S”形取样法采集剖面土壤样品,土壤剖面按0~20和20~40 cm自下而上分层采取原状土,每个小区3点混合成1个土样,装入干净的自封袋中,将土壤样品鲜土现场过2 mm筛后装入无菌自封袋和保鲜盒内(团聚体土样),冷藏带回实验室4℃保存。土壤样品鲜土现场过2 mm筛后装入无菌自封袋内,冷藏带回实验室4℃保存。
1.3 测定方法
1.3.1 土壤团聚体
土壤团聚体分级采用湿筛法,筛分收集各级土粒称重,把筛分的土壤样品按比例配成 50.00 g,自上而下的放入孔径为 2、1、0.5、0.25、0.106 mm的套筛中,然后在土壤团聚体分析仪上以 30 r·min-1的速度筛分 10 min(振幅 4 cm)。湿筛结束后将留在各级筛孔上的团聚体洗入铝盒中,50℃烘干后称量,计算水稳性团聚体的组成。
1.3.2 土壤养分
土壤理化性状均参考《土壤农化分析》中的方法测定。有效磷采用Olsen法、速效钾采用醋酸铵浸提火焰光度法、碱解氮采用碱解扩散法测定。
1.3.3 土壤酶活性
脲酶活性采用靛酚蓝比色法、蔗糖酶活性采用3,5-二硝基水杨酸比色法、过氧化氢酶活性采用高锰酸钾滴定法测定。
1.3.4 土壤微生物群落功能多样性
称取 10.0 g 新鲜土壤放入盛有 90 mL 无菌0.85%NaCl 溶液的三角瓶中,在摇床上180 r·min-1振荡30 min,于超净工作台中静置 30 min 后,将悬浮液稀释100倍,然后接种于 Biolog ECO 板中,每孔接种量为150 μL。接种完毕,将Biolog ECO 板置于 25℃恒温避光培养24、 48、72、96、120、144 h后,用Biolog Reader 分析仪读取590 nm处的光密度值。
1.3.5 数据处理
使用Excel 2007和PASW Statistics 18进行数据分析,使用Origin Pro 8.0作图。采用144 h的数据进行主成分分析及多样性指数比较。采用单因素方差分析和Duncan 最小显著性差异法比较处理间的差异。
(1)土壤团聚体的平均质量直径(MWD)和几何平均直径(GMD)的计算方法如下:
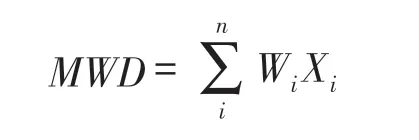
式中,Wi第i粒级团聚体质量百分数,Xi为相邻两级团聚体的平均粒径。

式中,Wi为土壤不同粒级团聚体的重量(g);lnxi为土壤粒级平均直径的自然对数。
(2)平均颜色变化率(AWCD)、Shannon指数、Mclntosh指数、Simpson指数(D)的计算方法, Simpson指数直接用D表示。 Gini系数(G)的计算参照Bronwyn D.Harch,计算公式如下所示:
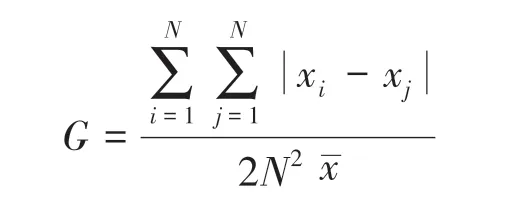
式中,xi、xj指每一种碳源的光密度值(1~31),N指碳源总数(31),指平均光密度值。
2 结果与分析
2.1 间作模式对根际土壤酶的影响
由图1可知,葡萄间作蔬菜对土壤酶活性的影响较大。整体上,0~20 cm 土层的土壤酶活性高于20~40 cm土层,说明浅层土壤酶活性高于深层土壤酶活性。0~20 cm土层中,GR、GI、GC土壤蔗糖酶活性显著高于其他4种间作处理(P<0.05),与CK相比分别增加了83.8%、81.7%、79.1%。土壤脲酶活性GG、GGa显著高于其他5种间作处理,与CK相比脲酶活性最大增加了74.9%,最小增加了约9.6%。过氧化氢酶活性由大到小依次为:GI>GB、GP>GGa>GR、GG、GC>CK,与CK相比,过氧化氢酶活性最大增加了105%,最小增加了38%。在20~40 cm 土层,土壤蔗糖酶活性由大到小依次为:GC>GI、GR>GP、GG>GGa、GB>CK,行间间作叶菜类蔗糖酶活性增加,增幅最大为186.7%,最小为30.6%。土壤脲酶活性由大到小依次为:GG>GR、GB、GP>GGa、GC>GI>CK,脲酶活性增加,增幅最大为131.2%,最小为26.1%。土壤过氧化氢酶活性由大到小依次为:GI>GB、GGa、GP>GR、GG>GC>CK。

图1 葡萄与蔬菜间作对葡萄根际土壤酶活性的影响
2.2 间作模式对根际微生物多样性的影响
2.2.1 代谢活性
Eco微生态板31种碳源平均颜色变化率(AWCD)的大小可反映微生物群落代谢活性的强弱。各处理的AWCD值在24~72 h培养时间段内缓慢波动上升,72 h之后快速升高。较单作而言,GC、GB、GI、GR根际微生物群落整体代谢活性增强,而GC、GGa、GG根际微生物群落整体代谢活性减弱,尤其在72~192 h 之间(图2),GP的AWCD增长最快。总体上,不同处理下对碳源的利用随时间的推移而不断升高,且存在“S”形的变化趋势。结果表明葡萄园不同的间作模式,对土壤微生物碳代谢能力存在差异,不同间作模式下,根区微生物多样性不同,对碳源的利用率不同;其中GP的微生物群落代谢活性最强,GG的微生物群落代谢活性最弱。

图2 葡萄与蔬菜间作根际土壤微生物AWCD随培养时间的变化
2.2.2 多样性指数
如表1所示,葡萄单作处理的Simpson指数低于间作模式(除GGa外),间作根区土壤的碳源代谢功能中优势度优于单作(P<0.05)。 葡萄单作Shannon指数显著高于GR(P<0.05),而GP、GB、GI、GGa、GC间作的Shannon指数显著高于葡萄单 作(P<0.05),分 别 提 高2.6%、2.1%、3.3%、4.3%、2.7%,说明间作的物种丰富度优于葡萄单作(P<0.05)。Mclntosh指数总体为升高趋势,与单作相比,GR、GP、GG、GB、GI、GGa、GC间作处理分别显著升高244%、215%、452%、300%、214%、495%、347%(P<0.05);Gini系数呈现升高的趋势,说明不同蔬菜与葡萄间作根际微生物群落的功能多样性存在差异,微生物群落不同碳源间的代谢能力差异显著,其中GG模式下碳源的代谢能力最强,而GC碳源代谢能力最小。总体来说,葡萄间作模式优于单作,而从微生物多样性指数的数据而言,最佳的间作模式为GP、GB,即葡萄间作辣椒和葡萄间作蚕豆处理。

表1 葡萄与蔬菜间作根际土壤微生物群落功能多样性指数
2.3 间作模式对根区土壤养分的影响
如图3所示,总体上,葡萄间作蔬菜土壤速效养分高于葡萄单作,各处理内土壤速效养分0~20 cm土层高于20~40 cm土层。0~20 cm土层中,GR、GG、GI、GGa、GP的碱解氮含量分别较CK显著提高63.91%、58.87%、54.17%、54.41%、60.63%(P<0.05);GI、GP的有效磷含量分别较CK显著提高46.23%、53.00% (P<0.05);GG、GB的速效钾含量分别较CK显著提高46.23%、53.00% (P<0.05);GGa、GG、GI的土壤有机质含量显著高于其他处理。20~40 cm土层中,GG、GP和GC的碱解氮含量分别较CK显著提高36.02%、51.51%和37.85%(P<0.05);GI、GP的有效磷含量分别较CK显著提 高94.14%、100.75% (P<0.05);GG、CB、GI的速效钾含量分别较CK显著提高69.51%、6.02%、60.00%(P<0.05)。GGa的土壤有机质含量显著高于其他处理。根据试验处理土壤碱解氮含量的变化规律,GR、GG、GGa、GP、GI优于其他间作及葡萄单作,而在有效磷的变化规律中,GI、GP间作优于其他组合,在速效钾的变化规律中,GG、GB、GI、GC优于其他间作或单作模式;土壤有机质大部分间作大于单作,浅层土壤有机质含量高于深层土壤。
2.4 间作模式对土壤团聚体的影响
从表2可知,间作使土壤的团聚体发生改变,不同的间作模式对团粒结构的改变有差异。总体而言,葡萄与蔬菜间作,可以增加团聚体的团聚度,提高抗蚀性,且稳定性越高。而7种间作模式中,从水稳性团聚体分级来看,0~20 cm 土层土壤的大部分团聚体为大团粒(>0.25 mm),占比平均高达67%,粗大团聚体(>5 mm)平均占比为12%,细大团聚体(2~5 mm)平均占比为18%。20~40 cm土层土壤大部分为大团粒结构,占比高达62%,粗大团聚体(>5 mm)平均占比为12%,细大团聚体(2~5 mm)平均占比为20%。其中,0~20 cm土层GB、GI处理土壤的MWD、GMD均高于其他模式,其团聚体稳定性较好。20~40 cm土层GR、GB、GC处理土壤的MWD、GMD均高于其他间作模式,其团聚体稳定性较好。
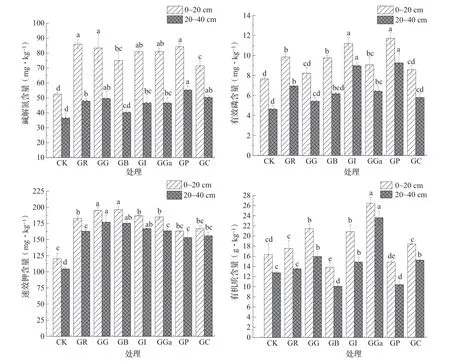
图3 葡萄与蔬菜蔬菜间作对根际土壤养分的影响

表2 葡萄与蔬菜间作对根际土壤团聚体的影响
2.5 化学指标和生物指标对土壤肥力影响的关联性分析
土壤养分与土壤微生物特性相关性表明(表3),土壤蔗糖酶、脲酶、过氧化氢酶活性及微生物多样性Mclntosh指数与土壤碱解氮、速效钾含量存在极显著或显著正相关关系,Gini系数与其存在负相关关系,但没有达到显著差异。土壤过氧化氢酶活性、Mclntosh指数与有效磷含量达到极显著正相关,土壤蔗糖酶活性、Gini系数与其存在正相关关系但未达到显著水平,脲酶活性与其存在负相关关系亦未达显著水平。土壤蔗糖酶、脲酶、过氧化氢酶活性与土壤有机质含量存在正相关关系,其中脲酶活性与其相关性极显著,其Mclntosh指数和Gini系数与其相关性不显著。

表3 土壤养分含量和微生物特性间的相关性
3 讨论
3.1 葡萄与蔬菜间作对土壤速效养分的影响
目前,国内外关于设施葡萄间作蔬菜的研究较少,主要是研究葡萄园生草对园区遮阴、水分的影响[17]。本研究对比了7种蔬菜与葡萄间作对土壤理化性质的影响,结果表明,在干热河谷地区不一定所有的间作模式都有利于土壤健康和可持续耕作。本研究发现,间作土壤理化性质优于单作;间作蔬菜可以使秋冬季葡萄园区土壤速效养分氮、磷、钾含量增加,最明显的特征就是表层土壤中的速效养分含量变化显著高于深层,间作制度有利于提高养分的有效性。与郭雄飞[18]关于生物炭对间作体系中刨花润楠土壤0~15 cm土层养分高于15~30 cm土层的研究结果一致,导致该结果的一个原因是间作促进葡萄根区与土壤养分循环有关的土壤微生物增多;另一个原因是间作使葡萄根区肥料释放出的养分,会促进蔬菜生长[19]。但本研究发现,间作土壤速效钾含量高,土壤速效钾含量浅层土壤高于深层土壤;而土壤钾含量高可以促进光合作用,提高CO2的同化率,调节根系生长和结实率等,促进植物抵抗胁迫[20-21]。这充分说明秋冬季节在设施葡萄园葡萄行间间作生长周期短的蔬菜类作物,有利于土壤保水保肥,而速效钾含量高,也有利于葡萄的生长和提高抗病抗逆性。
3.2 葡萄与蔬菜间作对土壤酶活性的影响
土壤酶主要来源于土壤微生物、土壤动植物[22-23],能催化土壤中的物质转化和能量流动,是植物生长和肥料利用的重要指标之一。土壤蔗糖酶参与碳循环,促进土壤易溶性营养物质转化[24];脲酶促进土壤氮转化,分解有机氮[25];过氧化氢酶促进土壤过氧化氢分解,以防止毒害作用。本文研究发现,葡萄与蔬菜间作,其土壤蔗糖酶、脲酶、过氧化氢酶活性均升高,其中GC、GR、GI的蔗糖酶活性最高,GGa的脲酶活性最高,GI的过氧化氢酶活性最高。司鹏等[26]研究发现,葡萄//草土壤酶活性高,深层土壤酶活性较低;这与张艳等[27]研究关于甘薯//玉米间作相比甘薯连作提高了蔬菜根际土壤酶活性的结果一致。
3.3 葡萄与蔬菜间作对土壤微生物的影响
土壤微生物群落结构和组成是对土地利用、耕地管理、植被、土壤养分、pH值变化的敏感指标[28]。本研究中葡萄间作不同蔬菜,在土壤有机质的变化中葡萄与大蒜间作效果最好。与前人研究关于大蒜与番茄间作的结果可以缓解连作障碍,促进微生物活性研究结果相同;可能是葡萄和大蒜间作使大蒜中大蒜素等分泌物增加[29],也与Castellano-Hinojosa等[30]研究的间作促进果树养分、微生物活性的结果相似。原因是葡萄//白菜间作相互作用,不同凋落物和分泌物刺激根区土壤,抑制土壤微生物群落活性,有机碳利用和转化效率;相比之下,辣椒、蚕豆分别与葡萄间作,其凋落物、根系分泌物等物质降低了微生物碳源转化和土壤有机碳的利用。这意味着在葡萄园利用间作不同的蔬菜,对根区微生物群落及有机质活性较好的模式是葡萄//蒜、葡萄//白菜、葡萄//青笋、葡萄//冰菜;与单作相比,间作改变了土壤微生物群落环境,这与马琨等[31]关于间作栽培对连作马铃薯根际土壤微生物群落影响的结果相似。这一结果可能是因为间作增加了植物种植种类,使土壤根区微生物的多样性和根系分泌物不同,促进了土壤微生物群落和优势种群的发展。另一方面,季节差异和深根植物与浅层植物间作,导致了空间根系分布差异大。
4 结论
间作改善了葡萄和蔬菜土壤微生物群落的多样性,提高了土壤脲酶、蔗糖酶、过氧化氢酶活性,与土壤有机碳、碱解氮、有效磷和速效钾含量的变化有关。因此,通过间作蔬菜可改变葡萄果园土壤理化性质,促进土壤有益微生物参与土壤养分循环,对土壤质量产生积极影响。
猜你喜欢
中国种业(2022年9期)2022-10-13
东南大学学报(自然科学版)(2022年4期)2022-08-12
北方果树(2022年1期)2022-01-21
养生阅刊(2021年5期)2021-06-03
安徽农学通报(2020年13期)2020-07-24
农民致富之友(2020年10期)2020-05-11
农民致富之友(2020年4期)2020-04-03
江苏农业科学(2019年7期)2019-09-17
中老年健康(2016年12期)2017-01-18
新农业(2016年11期)2016-12-27
