
云南高原湖泊水葱克隆生长与有性繁殖特征及影响因子
2022-08-15 07:57:28刘振亚牛孟莹肖德荣
广西植物 2022年7期
赵 飘, 刘振亚, 王 娜, 牛孟莹, 艾 静, 肖德荣, 王 行*
( 1. 西南林业大学 云南省高原湿地保护修复与生态服务重点实验室, 昆明 650224; 2. 西南林业大学 国家高原湿地研究中心/湿地学院, 昆明 650224; 3. 西南林业大学 生命科学学院, 昆明 650224; 4. 云南省农业科学院甘蔗研究所, 云南 开远 661600; 5. 温州大学 生命与环境科学学院, 浙江 温州 325035; 6. 温州大学 城镇水污染生态治理技术 国家地方联合工程研究中心, 浙江 温州 325035 )
生长与繁殖是植物实现种群维持与扩散的重要途径(王志保等,2018a),易受环境的变化而发生改变,进而对生态系统的结构和功能产生影响(Sherry et al., 2007)。植物生长与繁殖对未来气候变化的响应受到广泛关注(Roa-Fuentes et al., 2012;王志保等,2018b)。高原湖泊湖滨带植物对气候变化高度敏感(刘振亚等,2017),因此研究湖滨带植物生长与繁殖对外界环境因子的响应规律,是科学评估未来气候变化对高原湖泊有何影响和如何影响的基础。
地形(经纬度、海拔)、气候(降水、温度)和土壤养分等环境因子均对植物的生长和繁殖产生影响(Gong et al., 2019;Zhang et al., 2019)。海拔是影响植物生长和繁殖的主导因子(Körner, 2007),其中植物物候(Walker et al., 2014)、叶片功能性状(Jiang & Ma, 2015)与海拔密切相关。随着海拔的升高,植物的生物量增加(李亚芳,2015)、高度降低(Liu et al., 2016)、个体变小(Méndez & Traveset, 2003),而对繁殖的投入增加(Fabbro & Körner, 2004)。除海拔因素外,植物株高与纬度呈驼峰关系,中纬度最高(Liu et al., 2016),而随着纬度的增加,植物的株高降低(Moles et al., 2009)。地理空间的变化,使植物生境有所差异,有研究表明,温度增加对植物克隆生长或促进(Li et al., 2014)、或抑制(Kreyling et al., 2008)、或无显著影响(Bloor et al., 2010),对其有性繁殖具有明显的促进作用(王志保等,2018a;Xiao et al., 2019)。降水增加导致植物叶片气孔关闭、光合速率降低,植物克隆生长受到抑制(Körner, 2007),降水增加也能促进植物克隆生长(Roa-Fuentes et al., 2012)。增加土壤养分可促进植物个体的生长(Nasto et al., 2019),增加植物种子产量(王亚等,2021)。一般情况下,植物生长及繁殖在大尺度上受气候因子的影响(Svenning & Sandel, 2013),而在山地等区域尺度上则受海拔梯度的影响(唐志尧和方精云,2004)。在低海拔地区,土壤养分对植物生长与繁殖的影响更为显著,而在高海拔地区则是温度更为显著(Sang, 2009)。可见,研究植物生长和繁殖策略对环境变化的响应是生态学研究的热点,然而当前研究结果尚存在较多的不确定性。
云南省地形极为复杂多样,其西北部为横断山脉、东部为云贵高原,最大垂直高差达6 663.6 m,地形、气候及土壤等环境因子的空间差异明显,具有独特的地质和植被及立体气候特征(杨岚和李恒,2010)。湖泊作为云南高原生态屏障的重要组成,具有典型的“面山-湖滨-湖盆”的生态特征,在生物多样性和特有性保育等方面发挥着不可替代的作用,且对环境变化高度敏感(Xiao et al., 2019)。挺水植物是云南高原湖泊湖滨带的重要组成,是高原湖泊生态结构和功能的基础。水葱()是纳帕海、泸沽湖、拉市海、滇池、杞麓湖及异龙湖等6个高原湖泊湖滨带的共有种(杨岚和李恒,2010),其克隆生长和有性繁殖是其维持种群空间分布和扩散的重要途径。因此,研究水葱对不同地理空间环境因子的响应,能很好地诠释湖滨带挺水植物生长与繁殖对未来气候变化的适应策略。
本研究利用云南的立体地形及其独特气候,选择6个不同区域的高原湖泊,以其湖滨带共有分布的挺水植物水葱为研究切入点,研究水葱克隆生长(密度、株高、基径、地上生物量)以及有性繁殖(结实率、穗生物量、穗生物量投资比、单位面积种子产量、单位面积活性种子数)等在不同区域的差异特征,分析环境因子(地理位置、气候、土壤和水等环境因子)与水葱生长和繁殖指标的相关关系,拟解决以下科学问题:(1)高原湖泊挺水植物水葱生长和繁殖特征在地理空间分布是否存在地理分布差异;(2)其生长和繁殖特征是否存在经纬度、海拔地带性分布规律;(3)由于地理空间的差异而发生的环境变化对植物克隆生长与有性繁殖造成什么样的影响,其影响程度如何。以期为掌握高原湖泊对未来气候变化的响应机制奠定基础。
1 材料与方法
1.1 研究区概况
选择纳帕海、泸沽湖、拉市海、滇池、杞麓湖、异龙湖等6个湖泊为研究地。纳帕海、泸沽湖、拉市海位于滇西北高原,是滇西北高原生物多样性和特有性保护的热点区域,其中纳帕海和拉市海为国际重要湿地。滇池、杞麓湖、异龙湖位于滇中高原,既是滇中城市群经济社会可持续发展的生态屏障,也是关注水生态安全和生物多样性保护的重点区域。6个湖泊所处地形差异大,地理位置为99.66°—102.77° E、23.67°—27.85° N,海拔在1 412 ~ 3 260 m之间,跨经度、纬度分别为3.11°和4.18°,海拔高差达1 848 m。所处地理位置的差异,使6个湖泊的气候、水、土壤等环境因子存在一定差异(表1)。
1.2 样品采集
2020年8—10月,分别在纳帕海、泸沽湖、拉市海、滇池、杞麓湖、异龙湖湖滨带水葱典型分布区域,设置3 ~ 6个1 m × 1 m样方,其中纳帕海6个、泸沽湖4个、拉市海3个、滇池3个、杞麓湖3个、异龙湖4个,共计23个。野外测定样方内水葱总株数、结穗数等指标,并在样方内选取10株成熟结穗植株,测定植株高度,用精度为0.01 mm的游标卡尺测量基径。将结穗植株剪下带回实验室,测定穗的形态、生物量和种子活性参数。采用收割法,将样方内25 cm × 25 cm地上植株带回实验室,测定水葱地上生物量等参数。用GPS分别记录植物样方的经纬度和海拔。用定深泥炭钻(荷兰Eijkelkamp)采集每个样方内表层(0 ~ 10 cm)土壤样品,装入自封袋带回实验室,以测定土壤理化指标。用500 mL塑料瓶采集水样带回实验室,测定有关理化指标。
1.3 室内分析
1.3.1 地上生物量及有性繁殖指标的测定 采用烘干称重法,将刈割带回的植株样品放置65 ℃的烘箱中烘至恒重,测定地上生物量。首先,在室内用精度为0.1 cm的直尺测定每个样方带回的10株水葱的穗长,记录每穗小穗数、种子数等指标,计算水葱的结实率(结实株数/总株数×100%)、种子产量(每平方米种子数=单株平均种子数×每平方米结实株数)。然后,将分株装袋植株自然风干后,分别测定穗生物量和植株生物量,计算有性繁殖生物量投资比,即穗生物量投资比=穗生物量/植株生物量×100%。最后,将每个样方种子混匀,每个样方分别选取水葱种子200粒,3个重复,通过物理方法剥去种壳,用解剖刀沿种子侧面将胚纵切开,放置在1%四唑磷酸缓冲液中,在(30±1)℃条件下染色24 h,在10倍放大镜下分别观察其着色情况及种子活性。胚主要构造呈鲜红色,或胚根尖端2/3不染色而其他部分正常染色的种子为有生活力种子。在此基础上,计算单位面积活性种子数量,即活性种子数=种子产量×(着色种子数/染色种子数×100%)。
1.3.2 土壤和水体环境指标的测定 土壤有机碳含量(soil organic carbon,SOC)采用酸洗法测定(唐伟祥等,2018),土壤全氮(soil total phosphorus,STN)、全磷(soil total phosphorus,STP)采用HSO-HO消煮法测定。水样全氮(water total nitrogen,WTN)、全磷(water total nitrogen,WTP)采用连续流动分析仪(德国 SEAL Analytical AA3)测定。通过全球网格数据(precision: 0.16°×0.16°;http://www.paleo.bris.ac.uk/),分别获取各采样点的年均温(mean annual temperature,MAT)、最暖月均温(mean temperature of the warmest month,WMT)、最冷月均温(mean temperature of the coldest month,CMT)、年均降水(mean annual precipitation,MAP)等气候参数。各湖泊湖滨带土壤、水体采样点环境参数见表1。
1.4 数据处理
利用SPSS 19.0软件,对不同区域水葱密度、株高、基径、地上生物量、结实率、穗生物量及其投资比、单位面积种子产量、单位面积活性种子数量等生长繁殖指标进行单因素方差分析(One-Way ANVOA),差异显著性水平设定为= 0.05。通过Pearson相关性分析法,研究水葱生长和繁殖指标与气候、土壤、水文因子的相关性。在此基础上,通过Stepwise回归分析方法,进一步筛选影响水葱生长与繁殖特征的关键因子。使用R 4.01软件中Agricolae软件包对影响水葱生长与繁殖的关键因子进行路径分析,研究关键影响因子对水葱生长和繁殖的作用路径和贡献率。
2 结果与分析
2.1 生长与繁殖的空间差异
由表2可知,除地上生物量外,水葱生长与繁殖参数在不同地理空间上存在显著差异(<0.05)。其中,密度在纳帕海最高(853±99 plant·m),滇池最低(256±43 plant·m),在不同采样点间存在显著差异(<0.05);株高在异龙湖最高(209±28 cm),纳帕海最低(128±5 cm),在不同采样点间存在极显著差异(<0.001);基径在异龙湖最高(13.3±0.5 mm),拉市海最低(6.4±1.4 mm),在不同采样点间存在极显著差异(<0.001);结实率在异龙湖最高(71%±6%),在纳帕海最低(35%±6%),在不同采样点间存在极显著差异(<0.01);穗生物量在异龙湖最高(457±390 g·m),在拉市海最低(46±17 g·m),在不同采样点间存在显著差异(<0.05);穗生物量投资比在异龙湖最高(13.9%±3.9%),在纳帕海最低(2.2%±1.3%),在不同采样点间存在极显著差异(<0.001);单位面积种子产量在泸沽湖最高(18.0×10±6.6×10grain·m),在滇池最低(3.8×10±0.6×10grain·m),在不同采样点间存在显著差异(<0.05);单位面积活性种子数量在泸沽湖最高 (16.0×10±7.1×10grain·m),在滇池最低(2.7×10±0.3×10grain·m),存在极显著差异(<0.01)。
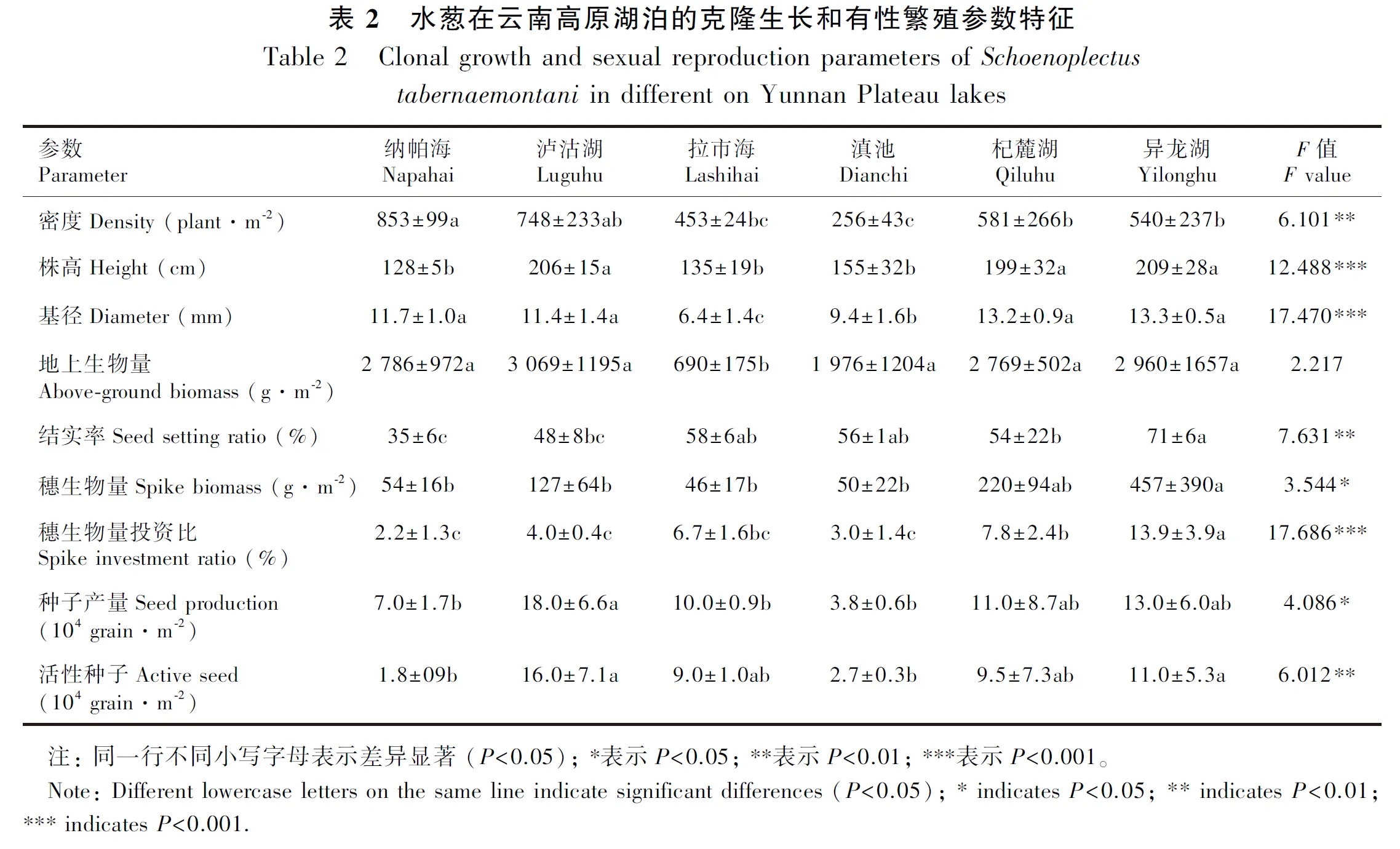
2.2 生长与繁殖的空间分布规律
水葱种群生长和繁殖指标在地理空间上存在一定的规律性变化(图1)。水葱密度随经度的增加而减小(图1:A),株高、结实率、穗生物量、穗生物量投资比指标随经度的增加而增大(图1:D,G,J,M);密度随纬度的增加而增加(图1:B),株高、结实率、穗生物量、穗生物量投资比等指标随纬度的增加而减小(图1:E,H,K,N);水葱密度随海拔的增加而增大(图1:C),株高、结实率、穗生物量、穗生物量投资比等指标随着海拔的增加而减小(图1:F,I,L,O)。
2.3 影响水葱生长与繁殖的因子及作用路径
水葱密度与地上生物量、种子产量呈极显著正相关(<0.01),株高与穗生物量、种子产量、活性种子呈显著正相关(<0.05),与穗生物量投资比呈极显著正相关(<0.01),基径与穗生物量呈显著正相关(<0.05),地上生物量与穗生物量、种子产量呈极显著正相关(<0.01),地上生物量与种子活性呈显著正相关(<0.05)(表3)。
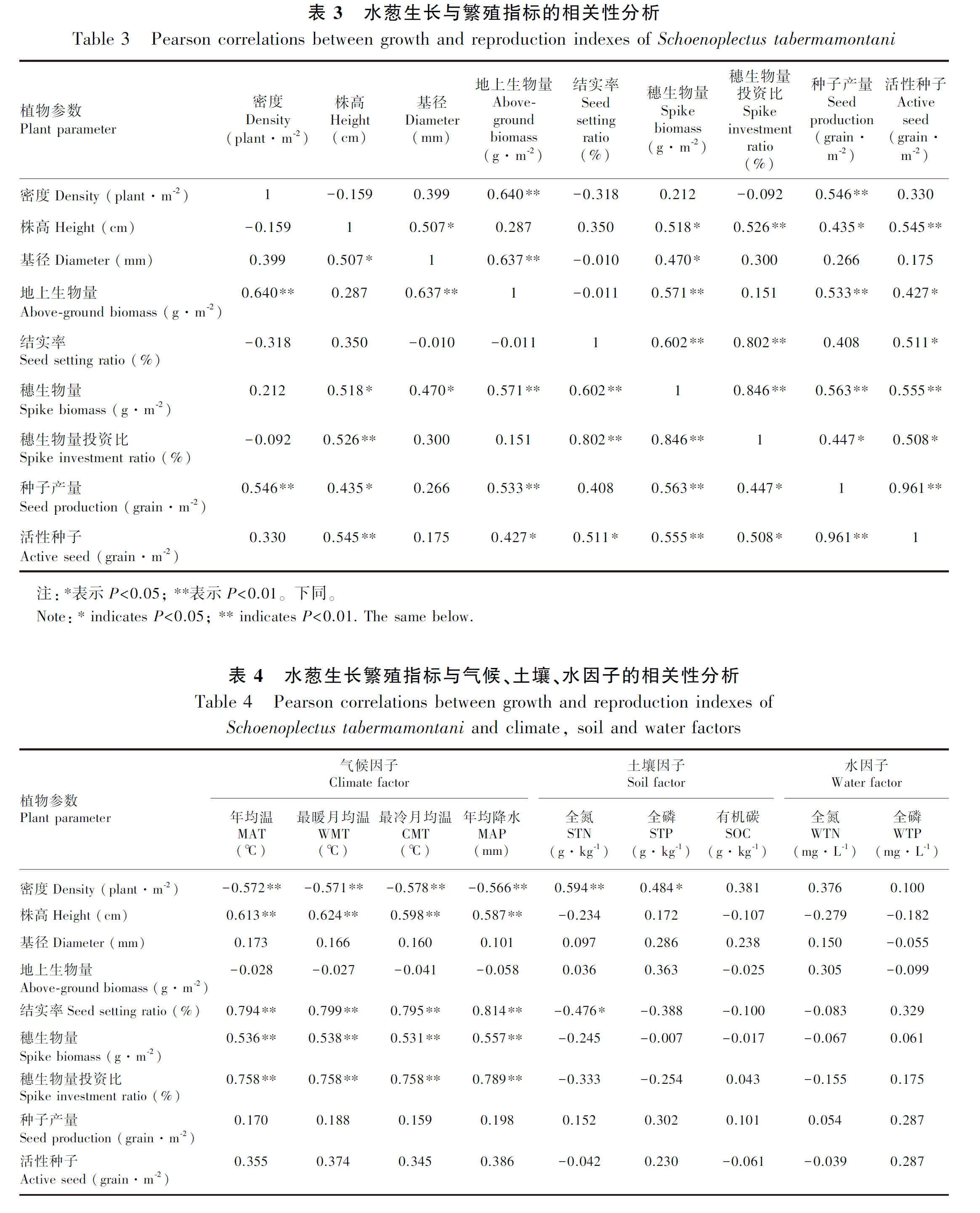
水葱生长和繁殖指标与区域气候因子和土壤因子存在显著相关,而与湖泊水因子无显著相关(表4)。其中,水葱密度与年均温、最暖月均温、最冷月均温、年均降水呈极显著负相关(<0.01),与土壤全氮呈极显著正相关(<0.01),和全磷呈显著正相关(<0.05)。植株株高与年均温、最暖月均温、最冷月均温、年均降水呈极显著正相关(<0.01),水葱基径和地上生物量与本研究环境因子无显著相关性(>0.05)。水葱结实率与年均温、最暖月均温、最冷月均温、年均降水呈极显著正相关(<0.01),与土壤全氮呈极显著负相关(<0.01)。穗生物量、穗生物量投资比与年均温、最暖月均温、最冷月均温、年均降水呈显著正相关(<0.05)。水葱单位面积种子产量、活性种子与本研究环境因子无显著相关性(>0.05)。
由图2可知,最暖月均温、最冷月均温、土壤全氮和全磷是影响水葱克隆生长的关键因子。其中,土壤全氮是影响水葱密度的主要因子,其路径系数为0.59,贡献度为35%;最暖月均温、最冷月均温、土壤全磷是影响水葱株高的主要因子,路径系数分别为0.74、-0.67、0.04,其对株高的贡献度超过90%(图2:A)。土壤有机碳、年均降水是影响水葱有性繁殖参数的关键因子,其中,年均降水直接影响水葱结实率、穗生物量、穗生物量投资比, 路径系数分别为0.81、0.56、0.82, 土壤有机碳直接影响水葱穗生物量投资比,其路径系数为0.32。年均降水对结实率和穗生物量的贡献度为66%和31%,年均降水和土壤有机碳对穗生物量投资比的贡献度为78%(图2:B)。

3 讨论与结论
植物生长和繁殖性状能够反映其对空间资源的利用和适应策略(陈玉宝等,2014)。本研究发现,水葱克隆生长(除地上生物量)和有性繁殖参数在不同分布空间存在较大差异,既是所在湖泊气候、土壤、水等环境因子协同作用的结果,也是水葱生长和繁殖策略对不同环境因子响应的体现。与前人对文冠果生长性状和水葱种子繁殖的研究结果一致(王志保等,2018b;张毅等,2019)。但是,不同研究地水葱地上生物量无显著差异,表明湖滨带植物生物量不受地理环境条件的影响,其生物产量适用于最终产量恒定定律。

克隆生长和有性繁殖是植物生活史策略中的两个重要方面,在不同的环境条件下植物不同性状间的关系反映了这些性状的功能联系。本研究中,水葱克隆生长与有性繁殖密切相关。水葱有性繁殖特征大小对植物个体大小表现出较强的依赖性,植物生长性状株高和基径与植物对光的捕获能力、抗机械损伤能力以及对有性繁殖器官的支持能力密切相关。主要表现为株高越高越容易使植物伸展获得更多的光照,但抗机械损伤能力就越弱;基径越大植物的抗机械损伤能力越强,同时能支持更粗的维管结构(Sun et al., 2016),在植物良好生长的同时有性繁殖性状得到促进。随着植物个体的增大,单株植物种子总数及种子总重量会增加(Susko & Lovett-Doust, 1998),这可以解释为植物个体越大,获得的总资源量越多,投入到有性繁殖部分的资源量会增加。
本研究中,水葱株高与温度因子密切相关,表明水葱克隆生长主要受温度因子的影响。这是因为温度的升高可以提高植物的光合效率、抑制植物的呼吸作用,使植物生长得到促进,同时,在较高温度下,植物生长季延长,升温加速了有机质的分解和土壤矿化,植物提高了对土壤养分的利用效率,从而促进植物的营养生长和碳积累(杨兵等,2010)。本研究结果与增温条件下植物根茎长度变化的研究结果一致(Li et al., 2014),有研究者采用OTC增温技术对极地物种的研究表明温度升高能使有性植株的株高增加(Stenström et al., 1997)。然而,在以往的研究中发现温度增加对植物克隆生长有抑制作用,存在这种差异的原因可能是非生物应激条件下植物对资源的强烈竞争产生了负面影响。另外,本研究结果显示水葱在降水量高的湖泊有更高的有性繁殖投资,而水体氮磷含量对植物生长和繁殖的影响不显著,这表明丰富的降水能促进挺水植物的有性繁殖,这可能与降水对湖泊湖滨带水淹情况的影响有关。由于挺水植物生境的特殊性,因此水作为湿地生态系统的三大要素之一,通常包括水量和水质两个方面。对于云南高原湖泊而言,其径流主要由降水补给(杨岚和李恒,2010),水葱花果期为6—9月,而云南雨季集中在5—10月,且降水量从南到北(纬度增加)呈递减的趋势(晏红明等,2018),其湖滨带水位情况发生改变。已有研究发现,地面水位变为零不利于生葱生长(赵湘江等,2015);香蒲在0 ~ 0.5 m水深范围内,随着水深的增加,有性繁殖比例增大(Sorrell et al., 2012)。本研究发现,土壤养分含量对植物生长和繁殖有重要影响,其中土壤氮含量对植物的克隆生长影响较大,这是因为植物主要从土壤中获取养分,而氮元素能促进细胞的分裂和增长,植物叶面积增长快,有更多的叶面积来进行光合作用,进而促进植物的克隆生长,本研究结果与增加土壤养分对植物个体生长的研究一致(Nasto et al., 2019)。本研究通过对植物克隆生长和有性繁殖特征的环境影响因子研究发现,其贡献度表现为气候因子>土壤因子>水因子。这表明在云南高原湖泊,相对于土壤、水等环境因子,区域气候因子(最暖月均温、最冷月均温和年均降水)是影响云南高原湖泊湖滨带植物水葱生长和繁殖的主要环境因子。因此,区域气候条件在高原湖泊植物生长和繁殖过程中影响最大,土壤养分含量对植物生长和繁殖有着显著影响,在未来环境变化下水环境因子对高原湖泊植物繁殖特征的影响需要进一步深入研究。
猜你喜欢
现代苏州(2022年5期)2022-03-21 22:57:09
今日农业(2020年19期)2020-12-14 14:16:52
少儿美术(快乐历史地理)(2019年4期)2019-08-27 00:51:40
阅读(低年级)(2018年4期)2018-05-14 17:39:57
小学阅读指南·低年级版(2017年2期)2017-03-23 13:07:24
新疆农业科学(2016年6期)2016-07-23 01:51:10
中国科技信息(2015年2期)2015-11-16 08:18:32
IT时代周刊(2015年9期)2015-11-11 05:51:51
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
少儿科学周刊·儿童版(2014年1期)2014-03-10 06:23:18
