
基于高通量转录组测序技术构建猪脂肪沉积的lncRNA 和miRNA 相关ceRNA 调控网络
2022-08-13 02:57刘怡冰翟丽维刘华涛赵卿尧马福平王楚端
中国畜牧杂志 2022年8期
刘怡冰,邢 凯,敖 红,翟丽维,刘华涛,赵卿尧,石 咏,李 航,马福平,王楚端*
(1.中国农业大学动物科学技术学院,北京 100193;2.北京农学院动物科技学院,北京 102206;3.中国农业科学院北京畜牧兽医研究所,北京 100193)
脂肪组织是机体的重要组成部分,是动物体内能量的主要储存形式。对于生猪产业,脂肪沉积性状也是重要的经济性状,显著影响着生猪的生产效率和繁殖水平。猪肉是中国居民动物蛋白食品的主要来源。随着生活水平的提高,人们对猪肉品质的要求越来越高。脂肪性状是评价动物肉质的主要指标之一,也影响着消费者对猪肉的选择。同时,由于相似的遗传特点和生理特点,猪也逐渐成为研究人类肥胖症和代谢综合征的理想模式动物。寻找影响猪脂肪沉积性状的分子标记是加快其遗传进展的一个重要途径,因此,探讨猪脂肪沉积的调控因子和分子机制具有重要的意义。
脂肪沉积的过程是一个复杂而精确的正负调控网络,受到多种基因和转录因子的调控,非编码RNA 在这个过程中发挥重要的作用。非编码RNA 包括长链非编码RNA(lncRNA)、microRNA(miRNA)和环状RNA(circRNA)等。2011 年,Salmena 等人提出了“竞争性内源RNA(ceRNA)”假说,这是非编码RNA 与蛋白编码基因通过miRNA 反应元件(MRE)竞争性结合miRNA 的一种新的分子调控机制。例如,lncRNA或circRNA都可以作为miRNA 海绵体吸附miRNA发挥ceRNA 的作用,对miRNA 的功能产生影响,进而调节mRNA 的表达,最终对各种生物学过程产生调控作用。目前,ceRNA 的研究思路已大量应用于癌症等疾病相关的研究,而鲜见针对猪脂肪沉积性状的ceRNA 研究。
本文通过鉴定在具有极端背膘厚度的长白猪个体之间差异表达的lncRNA、miRNA,并对其进行靶基因预测和功能富集分析,筛选参与猪脂肪沉积过程的关键lncRNA、miRNA 及基因,构建脂肪沉性状相关的ceRNA 调控网络,以期为非编码RNA 调控猪脂肪沉积性状的相关研究提供新的靶点。
1 材料与方法
1.1 实验样本选择 本研究的实验群体为长白后备母猪群体,来自天津宁河原种猪场,共132 头。在相同的条件(环境、饮水、日粮等)下饲养至100 kg 左右,使用HONGDA HS-1500 型兽用B 超仪测定猪只倒数第3、4 肋间的活体背膘厚度,再结合系谱信息,选择3 对具有极端背膘厚度的全同胞个体,按表型分为高背膘组(BH,9.86±1.14mm)和低背膘组(BL,3.70±0.80mm)。采集背部皮下脂肪组织,保存于液氮用于后续RNA 的提取。
1.2 RNA 提取及转录组测序 采用TRIzol 试剂盒提取背部皮下脂肪组织中的总RNA,使用NanoDrop 软件检测RNA 浓度及质量。反转录得到cDNA 后,构建链特异性文库并利用Illumina Hiseq 2500 测序仪进行双端RNA 测序和小RNA 测序,此工作由北京百迈客生物科技有限公司完成。
1.3 测序数据分析 首先,使用FastQC 和NGSQC Toolkit软件对测序数据进行质量评估及过滤,去除原始序列数据中含有接头及低质量的序列得到Clean reads。再使用Tophat2 软件将过滤后的测序数据与猪参考基因组(11.1)进行比对,之后使用Cufflinks及Cuffmerge 软件对转录本进行组装。接下来,使用CNCI、CPC 等编码能力预测软件预测lncRNA,使用mirDeep2 软件预测miRNA。
1.4 差异表达分析 使用DESeq2 软件获得lncRNA、miRNA 及mRNA 的表达量,并使用R 包edgeR 进行差异表达分析,筛选差异表达lncRNA、miRNA及mRNA 的条件为2 组中表达量差异倍数>2 以及FDR ≤0.05。
1.5 lncRNA、miRNA 的靶基因预测及功能分析 使用bedtools 软件搜索差异表达lncRNA 上下游50 kb 范围内的顺式靶基因,再对差异表达lncRNA 和基因的表达量进行spearman 相关性分析,预测lncRNA 的反式靶基因。使用Targetscan 软件及miRanda 软件预测miRNA 靶基因。使用在线网站DAVID(https://david.ncifcrf.gov/conversion.jsp)对靶基因进行GO 注释,使用在线网站WebGestalt(http://www.webgestalt.org/)进行KEGG 功能富集分析,值≤0.05 即为显著富集,得到的结果使用R 包ggplot2 进行绘图。
1.6 ceRNA 网络构建 使用RNAhybrid 软件通过序列互补情况预测lncRNA 与miRNA 的靶向关系,再结合lncRNA 与miRNA 的靶基因信息,使用cytoscape 软件进行ceRNA 网络的构建与绘制。
2 结果与分析
2.1 差异表达lncRNA、miRNA 及mRNA 的鉴定 本研究共鉴定得到1 975 个lncRNA,其中已知lncRNA 334 个,新预测得到的lncRNA 共1 641 个。进行差异表达分析之后得到220 个lncRNA 在高、低背膘组间差异表达,其中116 个在高背膘组表达上调,104 个在高背膘组表达下调(图1A)。本研究还鉴定得到351个已知miRNA,其中有18 个差异表达miRNA,6 个在高背膘组表达上调,12 个在高背膘组表达下调(图1B)。此外,本研究同样鉴定得到18 507 个基因,其中1 512 个在高低背膘组间差异表达,820 个在高背膘组表达上调,692 个在高背膘组表达下调。
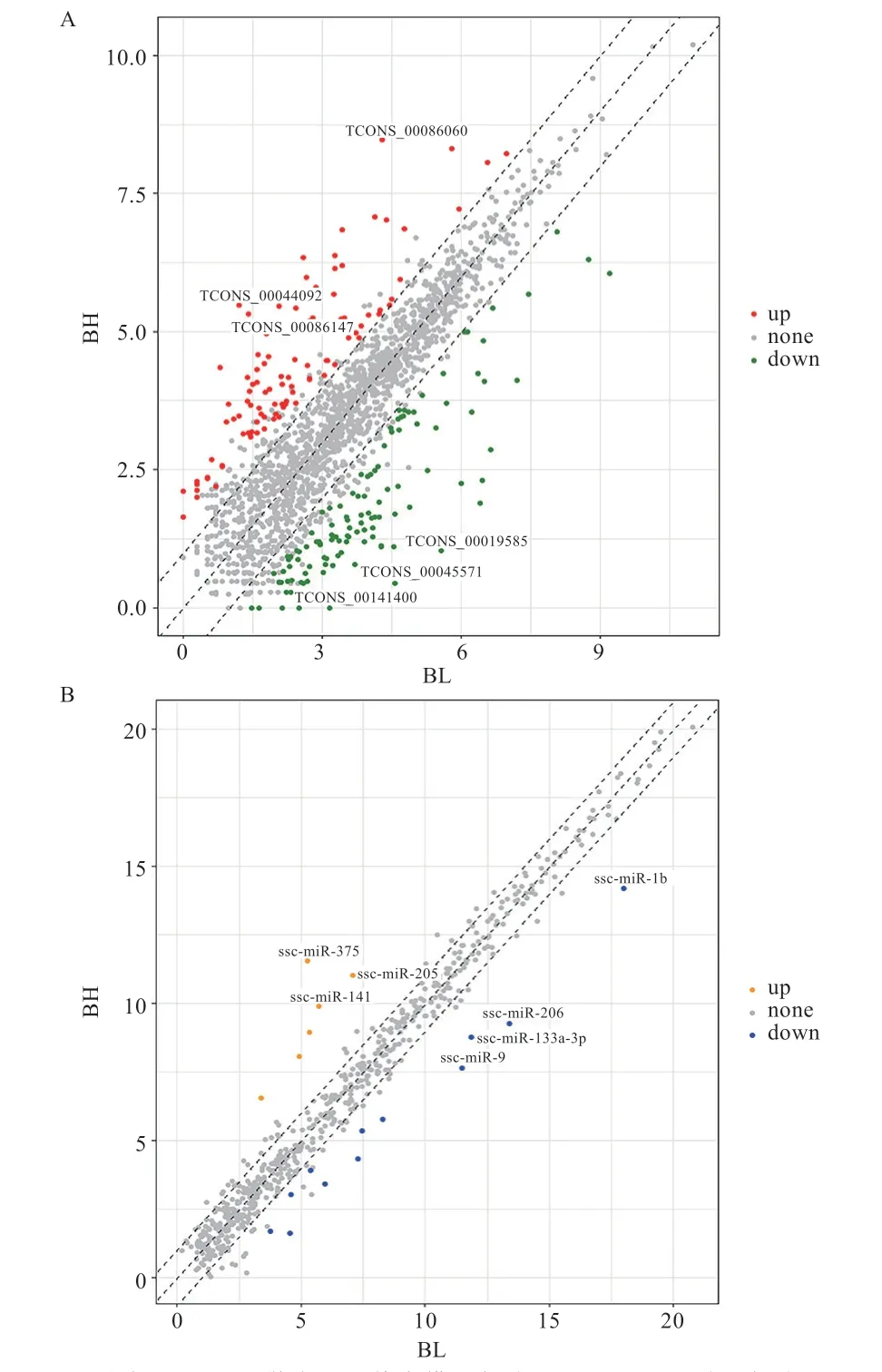
图1 lncRNA 和miRNA 差异表达分析
2.2 lncRNA 靶基因预测及功能富集分析 本研究共鉴定得到差异表达lncRNA 顺式靶基因67 个,反式靶基因116个。对这些靶基因进行GO 注释功能富集分析,生物过程类注释到的条目中,与糖类和脂类合成及代谢相关的条目包括“脂肪酸生物合成过程的正调控”(图2A)。接下来,将这些靶基因在KEGG 数据库中检索,在最显著的前20 条通路中与糖脂代谢相关的包括“脂肪细胞因子信号通路”、“糖酵解/糖异生”、“脂肪酸生物合成”和“类非酒精脂肪肝病(NAFLD)”等(图2B)。
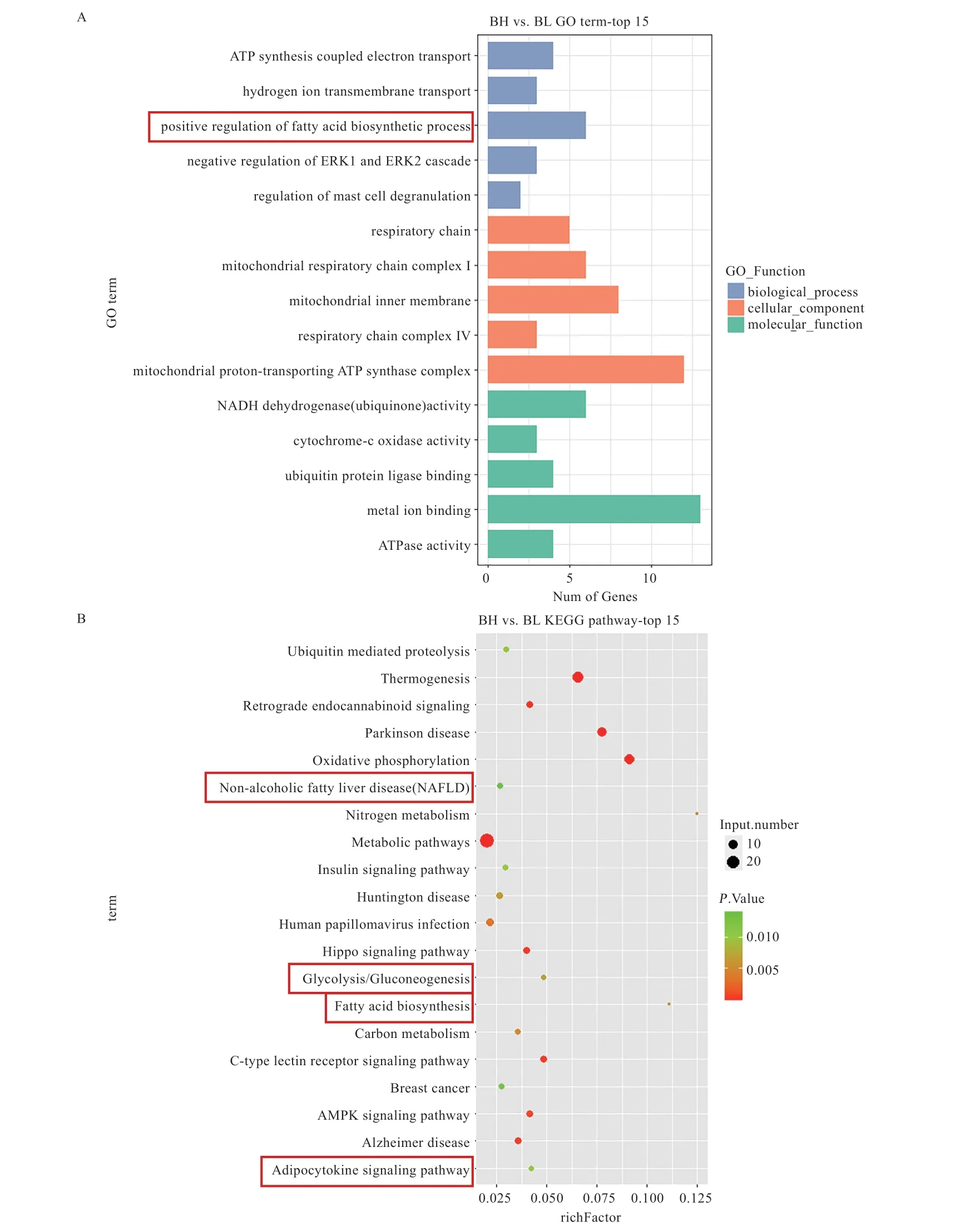
图2 lncRNA 靶基因的功能富集分析
2.3 miRNA 靶基因预测及功能富集分析 本研究共鉴定得到差异表达miRNA 靶基因863 个。对这些靶基因进行GO 注释功能富集分析,显著富集到生物过程、细胞组分和分子功能3 类注释的条目分别为41、14、15条,其中与糖类和脂类合成及代谢相关的条目包括“糖异生过程”、“糖酵解过程”及“脂肪细胞分化”等(图3A)。而通过KEGG 富集共得到29 条显著富集的代谢通路,其中最显著的5 条通路分别为“蛋白质消化吸收”、“ECM-受体相互作用”、“粘着力”、“类固醇生物合成”和“胰高血糖素信号通路”(图3B)。
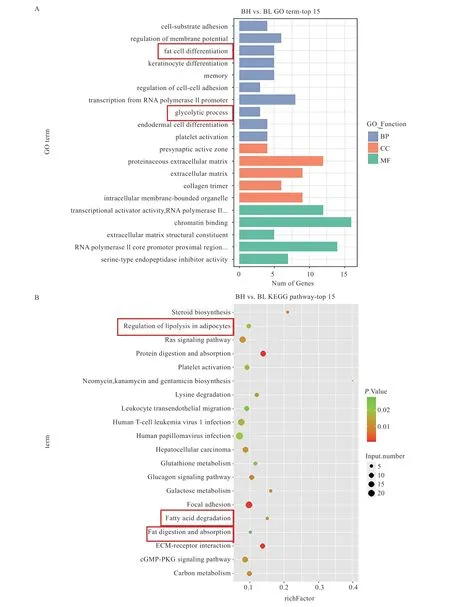
图3 miRNA 靶基因的功能富集分析
2.4 ceRNA 网络构建 差异表达lncRNA 共预测得到183 个靶基因,其中包括67 个顺式靶基因和116 个反式靶基因。再经过lncRNA 和miRNA 的靶向关系预测,本研究构建了一个脂肪相关的ceRNA 调控网络,包括9 个lncRNA、6 个miRNA 和25 个靶基因,如图4 所示。其中,有2 个lncRNA 为已知lncRNA(TCONS_00053896、TCONS_00142079),所有miRNA 均为已知miRNA。网络中所有lncRNA 及miRNA 在本研究中均在高、低背膘组间差异表达,所有靶基因均富集到脂肪生成或代谢相关的通路。

图4 脂肪沉积相关的ceRNA 网络构建
2.5 RT-qPCR 实验 为了验证RNA 测序结果的准确性,随机挑选在高低背膘组间差异表达的5 个基因、5 个lncRNA 和2 个miRNA 进行实时荧光定量PCR 实验,结果如图5 所示,同一基因在qRCR 实验和测序结果中表达趋势的变化相同,验证了转录组测序和分析的结果。

图5 RNA-seq 结果的RT-qPCR 验证
3 讨 论
ceRNA 的调控机制参与多种生物学过程,尤其是在人类的癌症发生过程中具有重要的作用。在Wang等的研究中发现lncRNA RHPN1-AS1 可以作为miRNA 海绵竞争性结合miR-596,从而提高基因的表达水平并激活FAK/PI3K/Akt 信号通路,促进卵巢癌肿瘤的发生和转移。并且,脂肪相关的非编码RNA 研究也在不断完善。Huang 等的研究发现,lncRNA MEG3 通过基因竞争性结合miR-21,抑制mTOR 通路,从而促进肝细胞内脂质积累,进一步诱导肝脏脂肪变性导致非酒精性脂肪肝的发生。在Wang 等的研究中,lncRNA IMFlnc1 通过吸附miR-199a-5p 促进的表达,从而通过cAMP 通路促进猪肌内脂肪细胞脂肪生成。
本研究成功构建了脂肪相关ceRNA 调控网络,其中包括9 个关键差异表达lncRNA,均为新预测得到的lncRNA。6 个关键差异表达miRNA 分别为sscmiR-486、ssc-miR-9、ssc-miR-34c、ssc-miR-31、sscmiR-205 和ssc-miR-133a-3p,他们对脂肪沉积过程的调控作用已在不同物种中被部分证实。Mentzel 等的研究发现,miR-9 在肥胖猪的皮下和腹部脂肪组织中显著高表达。而在人类糖尿病患者的血清中,miR-9 的表达水平高于健康人群。miR-34c 是miR-34 家族成员之一,研究发现在3T3-L1 前脂肪细胞分化过程中miR-34c 的表达上调。Jones 等构建的miR-34c 早 期过表达的小鼠,1 个月大时就会出现胰岛素抵抗,随着年龄的增长,会出现葡萄糖代谢受损和内脏脂肪生成失调,这些变化也是成人肥胖和糖尿病的症状之一,所以miR-34c 转基因小鼠可以作为研究成人肥胖的良好模型。有研究表明,miR-205 可以通过靶向基因改善非酒精性脂肪肝中的脂质积累。在高脂喂养的非酒精性脂肪肝(NAFLD)小鼠模型中,miR-205 的过表达会导致基因表达水平的降低,从而导致小鼠体重和肝脏重量下降,肝脏甘油三酯减少,甘油浓度升高,最终抑制脂质积累。Gottmann 等发现,miR-31在小鼠性腺白色脂肪组织中特异表达,靶向和等基因,从而参与脂肪生成和胰岛素信号传导通路。并且miR-31 在人和小鼠中的序列保守,在肥胖人群和二型糖尿病患者的内脏脂肪组织和血清中,miR-31 的表达显着高于健康人群,这说明miR-31 有可能可以作为治疗二型糖尿病的新型靶标。
另外,ceRNA 调控网络中的25 个靶基因中,也有一些是已被证实的脂肪沉积调控基因。是sscmiR-205 的靶基因,该基因是酰基辅酶A 合成酶家族成员,编码线粒体酰基辅酶A 合成酶,可催化中链脂肪酸代谢过程,进而减轻肝脏脂肪变性。是指维生素D 受体,同样受到ssc-miR-205 的调控,Xu 等的研究发现脂肪组织中的过度表达会导致小鼠体重、脂肪量和血清脂质水平增加,抑制产热基因的表达,进一步导致能量代谢下降,说明通过调节脂肪组织的重塑在糖脂代谢和能量稳态的调节中起关键作用。而是ssc-miR-205 和lncRNA TCONS_00026612的靶基因。有研究表明,基因可以通过抑制过氧化物酶体增殖物激活受体2()的转录来抑制脂肪生成,它是脂肪生成的主要促进剂。是ssc-miR-133a-3p 的靶基因。该基因编码的蛋白质作为游离脂肪酸(包括omega-3)的受体,通过快速激活cAMP 的产生促进脂肪生成。Ichimura 等发现,在肥胖人群脂肪组织中的表达明显高于对照组,其突变体p.R270H 增加了欧洲人群肥胖的风险,因此该基因在控制人类和啮齿动物的能量平衡方面具有关键作用。基因产生的脂联素是脂肪细胞分泌的一种内源性生物活性多肽,在肥胖患者中表达下调。Gao 等的研究证明过表达可以增强3T3-L1 成纤维细胞增殖,加速脂肪细胞分化,沉默该基因则会抑制猪前脂肪细胞的分化和脂质积累。在肝脏中,可以增强脂肪酸的氧化,以减轻肝脏脂质积累。
4 结 论
本研究以3 对具有极端背膘厚度的长白猪全同胞个体为研究对象,通过链特异性建库RNA 测序和小RNA测序,鉴定了猪脂肪组织中的lncRNA 和miRNA。进行差异表达分析之后共得到220 个差异表达lncRNA 和18 个差异表达miRNA。进一步对他们进行靶基因预测和靶向关系分析,最终本研究成功构建了一个包含9 个lncRNA、6 个miRNA 和25 个靶基因的与脂肪沉积过程相关的ceRNA 调控网络,为探究猪脂肪沉积性状的分子调控机制提供了新的见解。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
中国医学物理学杂志(2022年9期)2022-10-09
表面技术(2022年9期)2022-09-27
辐射研究与辐射工艺学报(2022年4期)2022-08-29
中国现代医生(2022年19期)2022-08-25
中国典型病例大全(2022年11期)2022-05-13
中国典型病例大全(2022年7期)2022-04-22
汽车工程师(2021年12期)2022-01-18
科学导报(2021年29期)2021-06-03
科海故事博览·下旬刊(2019年6期)2019-04-16
