
铁皮石斛AP3-3基因及启动子的克隆与分析
2022-08-12 07:12:16王义琴师凯辉王冬梅和凤美
福建农业学报 2022年5期
王义琴,孙 博,何 玲,师凯辉,黄 鑫,王冬梅,陈 宇,臧 睿,和凤美
(云南农业大学园林园艺学院,云南 昆明 650201)
0 引言
【研究意义】铁皮石斛(Dendrobium officinaleKimura et Migo)为 兰 科(Orchidaceae) 石 斛 属(Dendrobium)多年生草本植物,花具有较高观赏和药用价值。MADS-box是一类编码转录因子的基因家族,调控花器官的发育,主要由A、B、C、D、E等5类基因组成,AP3亚家族属于B类基因,包括AP3-1、AP3-2和AP3-3等3个进化枝,其中AP3-3通常只在花瓣特异性表达且表达水平很高[1]。克隆铁皮石斛AP3-3基因及启动子序列进行生物信息学分析,研究DoAP3-3的启动子活性,对于探究铁皮石斛花发育调控蛋白的功能及转录起始水平上的基因调控具有重要意义。【前人研究进展】MADS-box基因所编码的蛋白与目标DNA序列或蛋白结合产生互作,激活调控下游靶基因,影响花器官形态建成。B类AP3和PI蛋白以异源二聚体形式结合DNA,在一个自反馈调节环中以自身启动子维持转录[2]。在花瓣和雄蕊发育过程中,AP3/PI可通过控制靶基因NAP所编码的NAC家族蛋白NAP去诱导并控制细胞从分裂过渡到伸长[3]。AP3-3基因在时间和空间上的转变模式决定兰科植物不同类型花瓣器官的生长发育,参与花被形态多样性[4],在唇瓣发育中起着至关重要的作用,在惠兰、鹤顶兰、杂交兜兰等少数兰科植物中已通过RT-PCR方法成功克隆[5]。目前研究启动子活性常用启动子5’端缺失,研究内容主要集中在蔬菜水果、粮食及其他作物的生理生化方面,例如:番茄耐热基因SlDREBA4启动子在−1095~−730 bp 及−162~−38 bp 两个区间响应热胁迫[6];荔枝多酚氧化酶PPO基因在966、668、426 bp长度的3个启动子片段均具有活性,并在叶片及果皮表达[7];玉米II型H+焦磷酸酶基因ZmGAPP启动子在−219~−1 bp区域活性最高,响应干旱和盐胁迫[8];棉花GhAO1 启动子在−1760~−1600 bp 片段含 TGA 元件,可介导生长素信号通路,影响纤维细胞发育[9];也有少数关于花发育有关基因启动子的缺失分析研究,如二球悬铃木的PaAP3、PaPI和PaSTK[10]。【本研究切入点】兰科植物的基因启动子研究较少,尤其是MADS-box家族,主要集中在功能及表达情况,其中铁皮石斛AP3-3基因及启动子的相关研究鲜见报道。【拟解决的关键问题】克隆DoAP3-3基因cDNA及启动子序列,分析DoAP3-3蛋白结构功能及启动子序列含有的顺式作用元件,并构建全长及两个5’缺失启动子片段与GUS报告基因的融合表达载体,对铁皮石斛原球茎进行瞬时表达,验证DoAP3-3启动子的活性功能,为建立高效稳定的铁皮石斛花发育基因调控机制提供理论基础。
1 材料与方法
1.1 试验材料
铁皮石斛(D.officinale)组培苗由本实验室保存,大肠杆菌菌株DH5α和根癌农杆菌菌株EHA105购自 TOLOBIO 公 司 。 Plant Genomic DNA Kit购自TSINGKE 公 司 , Universal DNA Purification Kit、TIANprep Mini Plasmid Kit和 RNAprep pure Plant Kit购自 TIANGEN 公司,RT reagent Kit购自 TaKaRa公司,Gusblue kit购自北京华越洋公司,TaqDNA聚合酶、T4-连接酶、Eco31 I限制性内切酶购自BioRun公司。引物合成及目的片段测序由昆明擎科生物科技有限公司完成。
1.2 试验方法
1.2.1DoAP3-3 基因及启动子序列片段克隆 本试验室前期已测序分析铁皮石斛基因组及转录组数据。选择NR数据库注释为AP3-3的基因设计扩增引物(表1)。以铁皮石斛新鲜采摘的花朵为材料,提取总RNA,反转录为cDNA,通过RT-PCR扩增目的条带,产物经1%琼脂糖凝胶电泳后,切胶回收650 bp左右大小条带,与pMD19-T载体进行连接,热激法转入大肠杆菌感受态细胞DH5α,阳性克隆鉴定后送测序。
选择DoAP3-3 基因 ATG 上游长度为 2000 bp 序列为预测启动子,设计引物克隆全长。使用SoftBerry在线网站(http://www.softberry.com/)预测分析克隆所得启动子序列所含转录起始位点,依此确定启动子缺失片段长度,设计已添加Eco31 I酶切位点的上下游引物克隆系列5′端不同启动子片段,设计菌检引物检测启动子片段是否与GUS表达载体连接(表1)。以铁皮石斛苗叶片为材料,提取总DNA为模板进行常规PCR扩增。产物电泳后切胶回收2000 bp大小条带,与pMD19-T载体进行连接,转入大肠杆菌感受态细胞DH5α,阳性克隆鉴定后送测序,克隆载体命名为pMD19-T-DoAP3-3。

表1 PCR引物信息Table 1 PCR Primer information
1.2.2DoAP3-3 基因生物信息学分析 使用 DNAMAN软件将核酸序列翻译为氨基酸序列并进行同源比对;MEGA6.0构建系统进化树;ProtParam(https://www.expasy.org/resources/protparam)分析蛋白质理化性质;SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测蛋白质二级结构,Swiss-Model (https://swissmodel.expasy.org/)预测三级结构,TMHMM 2.0(https://services.healthtech.dtu.dk/service.php?TMHMM-2.0)分析跨膜区域,PSORT(https://www.genscript.com/psort.html)分析蛋白质的亚细胞定位。
1.2.3DoAP3-3启动子区域的顺式作用元件预测 基于 Rombauts等[11]的方法,使用 Plant CARE 数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html)对克隆所得DoAP3-3启动子序列的顺式作用元件进行预测分析。
1.2.4 启动子 GUS 表达载体的构建 以 pMD19-TDoAP3-3为模板,克隆不同长度含Eco31 I酶切位点的启动子片段,电泳切胶回收目的片段,测序无误后对其进行Eco31 I酶切。pBWA(V)HG载体进行同样酶切去掉CaMV 35S启动子,经过电泳分离后回收载体骨架。将载体酶切骨架与启动子酶切产物合并纯化后,通过连接反应构建重组载体,分别命名为pB-DoAP3-3-A、pB-DoAP3-3-B、pB-DoAP3-3-C。将10 μL 重组质粒转化大肠杆菌感受态,涂(卡纳霉素)抗性培养皿,37 ℃培养12 h,筛选阳性克隆并测序,检测无误后通过冻融法将重组质粒转入农杆菌EHA105获得工程菌株。
1.2.5 启动子载体的瞬时表达及GUS活性检测 将转化成功的EHA105加入抗性(卡纳霉素) LB中培养过夜,离心进行菌液重悬,并加入 200 μL·L−1Silwet-77,保持OD600=0.6;将1~2 mm厚的铁皮石斛原球茎(颜色浅黄)切片,加入重悬菌液,在25 ℃、60 r·min−1条件下侵染 48 h,当 OD600=1 时,更换新的菌液 ;GUS 染色液 37 ℃ 染色 24 h,70%酒精进行脱色处理。
2 结果与分析
2.1 DoAP3-3 基因克隆
以反转录所得cDNA为模板,反应体系(50 μL):Nuclease-free Water 20 μL,Mixture 25 μL,上游引物1 μL,下游引物 (R) 1 μL,cDNA 3 μL;反应程序:95 ℃ 5 min,95 ℃ 25 s,58 ℃ 25 s,72 ℃ 1 min,30个循环,72 ℃ 3 min,16 ℃ 30 min。PCR 扩增后得到一条长度为500~750 bp的DNA片段(图1A) 。切胶回收与pMD19-T载体连接,菌落PCR鉴定后送交公司测序,结果表明DoAP3-3的cDNA大小为675 bp。
2.2 DoAP3-3 启动子片段克隆
以所提取的铁皮石斛基因组DNA为模板,反应体系(50 μL):Nuclease-free Water 20 μL,Mixture 25 μL,上游引物 2 μL,下游引物 (R) 2 μL,Template 1 μL;反应程序:94 ℃ 5 min,94 ℃ 30 s,50 ℃ 45 s,72 ℃2 min,30 个 循 环 , 72 ℃ 10 min, 16 ℃ 30 min。PCR 扩增后得到一条长度为 1500~2000 bp 的 DNA片段(图1B)。切胶回收与pMD19-T载体连接后送公司测序,结果表明克隆到正确条带,大小为1 885 bp。
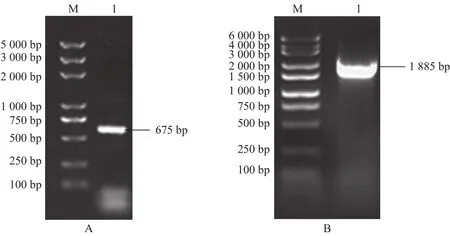
图1 PCR扩增电泳Fig.1 Electrophoresis of PCR amplification
2.3 DoAP3-3 蛋白质生物信息学分析
2.3.1 DoAP3-3 氨基酸序列同源性分析 铁皮石斛DoAP3-3基因cDNA大小为675 bp,预测编码224个氨基酸,通过MEGA6.0构建铁皮石斛与其他物种AP3-3氨基酸序列(NCBI下载)的系统进化树。结果表明DoAP3-3与石斛杂交种(Dendrobiumhybrid cultivar)、鹤顶兰(Phaius tancarvilleae)、杂交兜兰(Paphiopedilumhybrid cultivar)、 惠 兰(Cymbidium faberi)、多节拉索兰(Brassavola nodosa)、直立山珊瑚(Galeola falconeri)等兰科植物亲缘关系较近,属于一个进化枝(图2)。使用DNAMAN将铁皮石斛与上述兰科植物的AP3-3氨基酸序列进行同源比对,结果如图3所示,总体一致性84.78%。
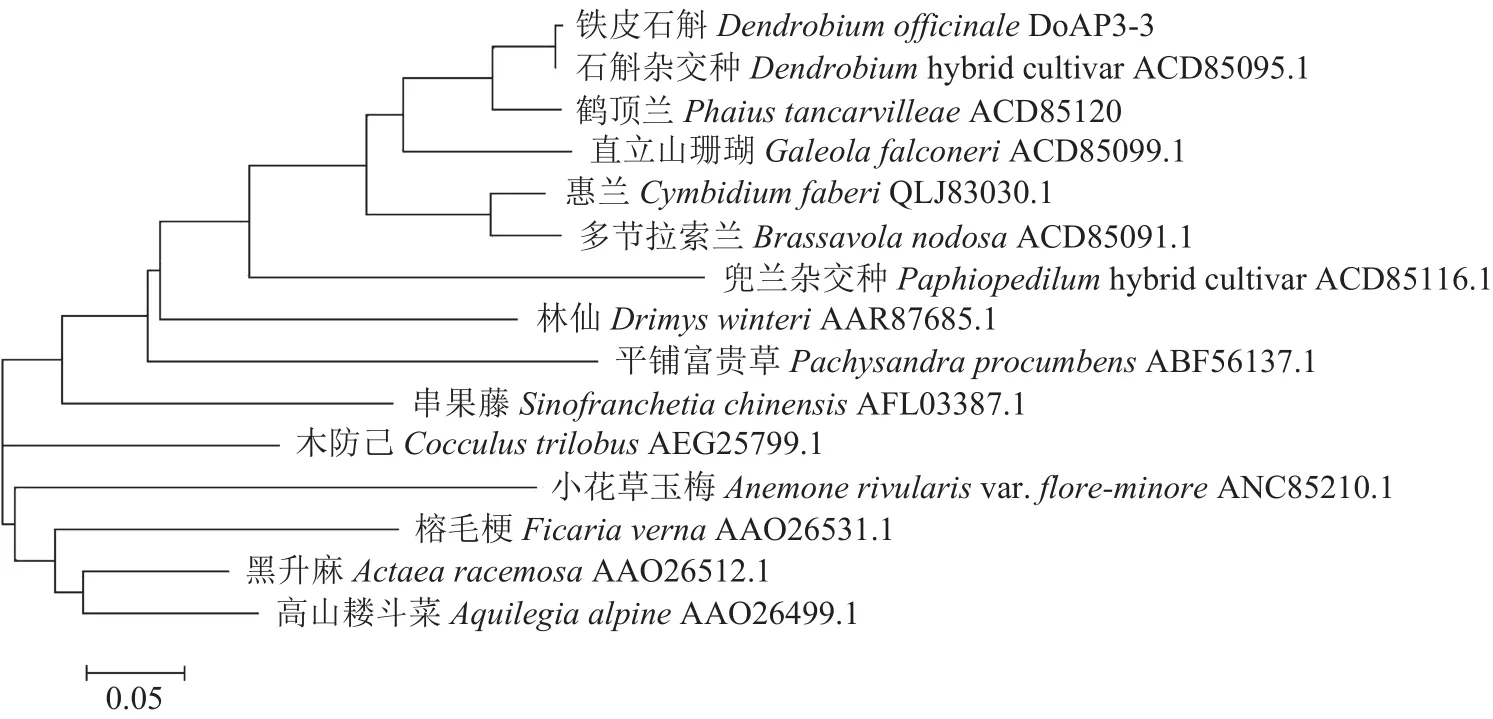
图2 DoAP3-3氨基酸序列与其他植物同源序列的系统进化树Fig.2 Phylogenetic tree of DoAP3-3 and homologous amino acid sequences in other plants

图3 DoAP3-3与其他植物AP3-3氨基酸序列的同源比对Fig.3 Homologous alignment of amino acid sequences between DoAP3-3 and AP3-3 in other plants
2.3.2 DoAP3-3 蛋白 质 结 构 及 功能预 测 ProtParam 分析结果表明:DoAP3-3蛋白质分子式为C1 129H1803N333O347S12,分子量25.98 kDa,负电荷残基总数(Asp+Glu)为31,正电荷残基总数(Arg+Lys)为34,等电点(pI)8.71,为碱性蛋白。该蛋白质预估半衰期:30 h(哺乳动物网织红细胞,体外)、大于20 h(酵母,体内)、小于10 h(大肠杆菌,体内)。脂肪指数71.79,不稳定性指数40.14,为不稳定蛋白。亲水性总平均值(GRAVY)为−0.823,小于0,为亲水蛋白。
使用SOPMA在线预测DoAP3-3蛋白质二级结构,结果(图4)显示 :该蛋白质的二级结构含有54.91% α-螺旋、13.84%延伸链、6.70% β-转角、24.55%无规则卷曲。使用Swiss-Model对DoAP3-3基因编码蛋白质的三级结构进行三维建模,结果如图5。
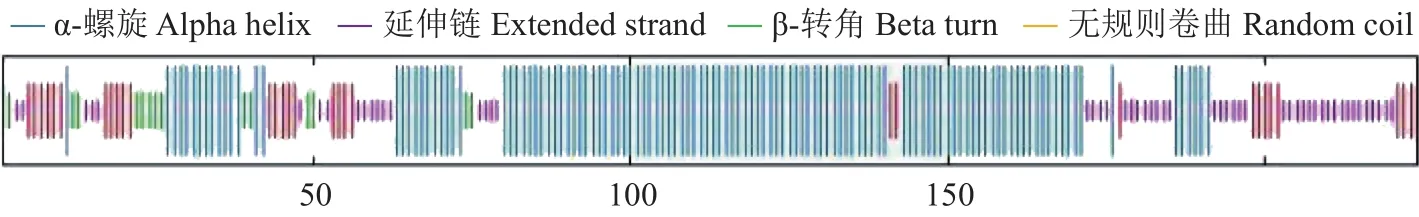
图4 DoAP3-3蛋白质二级结构预测Fig.4 Predicted secondary structure of DoAP3-3 protein
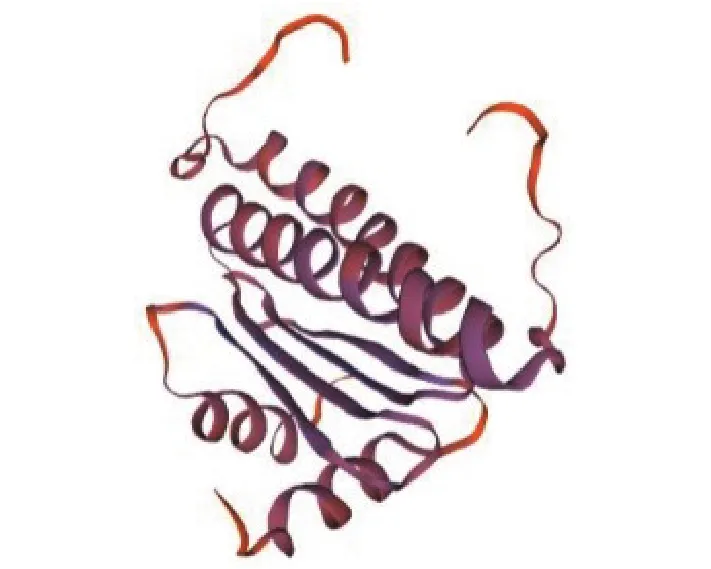
图5 DoAP3-3蛋白质三级结构预测Fig.5 Predicted tertiary structure of DoAP3-3 protein
使用TMHMM 2.0在线程序预测分析DoAP3-3蛋白可能存在的跨膜螺旋结构区域结果表明,DoAP3-3蛋白的1~224位氨基酸均不存在跨膜区域,且位于膜外,说明该蛋白不是膜受体蛋白,与细胞信号转导无关。通过PSORT对DoAP3-3蛋白质进行亚细胞定位在线预测,得分为:细胞核87.0%、线粒体8.7%、细胞质4.3%,在细胞核上的得分较高,预测该蛋白主要存在于细胞核中。
2.4 DoAP3-3 启动子生物信息学分析
使用Plant CARE对克隆所得启动子片段进行在线分析(表2),结果显示DoAP3-3的启动子区域除含多个转录起始核心元件TATA-box外,还分布了涉及脱落酸、赤霉素、生长素、水杨酸的反应元件,分生组织表达相关调控元件,光响应元件,MYB结合位点等多个顺式作用元件。

表2 DoAP3-3启动子顺式作用元件预测分析Table 2 Predicted DoAP3-3 promoter cis-acting element
2.5 启动子与 GUS 的重组质粒构建
以pMD19-T-DoAP3-3为模板,设计4条引入Eco31 I酶切位点的上下游引物,克隆大小为 1885 bp、1604 bp、750 bp 的启动子目的片段,结果如图6(A)所示。经酶切后与pBWA(V)HG连接。酶切反应体系 : Nuclease-free Water 13 μL,10× Buffer 2 μL,Eco31 I 1 μL,启动子 4 μL;程序:37 ℃,1 h。连接反应体系:Nuclease-free Water 5.5 μL,10×Buffer 1 μL,T4-ligase 1 μL,纯化产物 2.5 μL;程序:25 ℃,1 h。重组质粒转入大肠杆菌,使用菌检引物扩增部分启动子片段及载体GUS基因的特异片段,结果如图6(B)所示,证明3个启动子片段与pBWA(V)HG成功连接,将GUS重组表达载体转入农杆菌获得工程菌株。
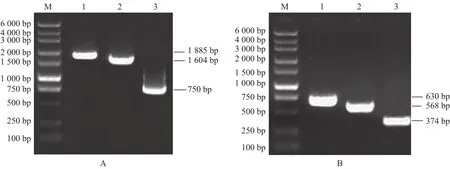
图6 启动子融合GUS重组载体构建Fig.6 Construction of fusion recombinant vector of promoter and GUS
2.6 启动子活性分析
通过农杆菌介导法将pB-DoAP3-3-A、pB-DoAP3-3-B、pB-DoAP3-3-C分别转化铁皮石斛原球茎,瞬时表达2 d后进行GUS染色,由图7可知−1885~0 bp 染上的蓝色最深,−1604~0 bp 染色次之,−750~0 bp染色最浅。说明DoAP3-3启动子能够驱动GUS基因的表达,且随着启动子缺失长度增加,启动活性逐渐减少。
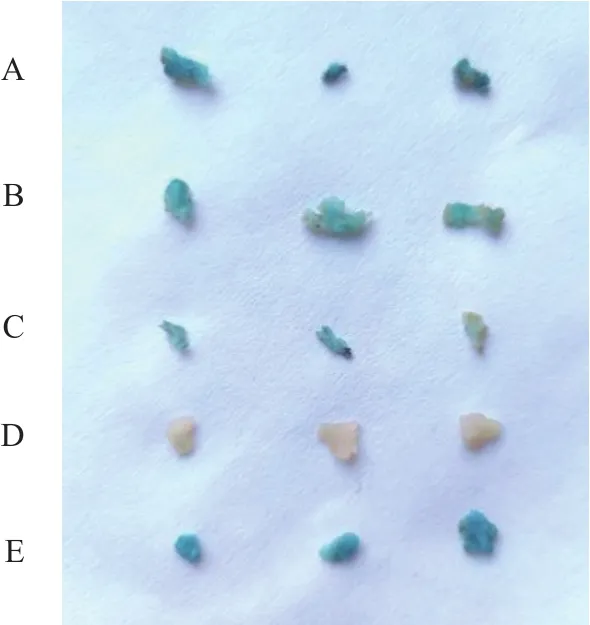
图7 转基因铁皮石斛原球茎GUS染色Fig.7 GUS stained protocorm of transgenic D.officinale
3 讨论
AP3-3作为花瓣身份特征基因[12],对铁皮石斛的花器官发育具有重要作用。本研究通过RT-PCR方法克隆得到铁皮石斛DoAP3-3的cDNA序列,长度675 bp,编码的氨基酸序列与兰科植物的进化亲缘关系较近,序列一致性较高,证明克隆所得序列是铁皮石斛AP3-3基因。DoAP3-3蛋白分子式C1129H1803N333O347S12,分子量 25.98 kDa,为碱性亲水蛋白,理化性质不稳定易失活;DoAP3-3不存在跨膜结构区域,说明该蛋白不是膜受体蛋白,与细胞信号转导无关。亚细胞定位预测结果在细胞核上的得分较高,说明该蛋白可能主要在细胞核中存在。Jing等[13]研究结果证明枇杷EjAP3的亚细胞定位在细胞核内,与铁皮石斛DoAP3-3预测结果一致,这可能与DoAP3-3作为转录因子通过与下游靶基因启动子结合,进而调控表达的特性有关。
本研究克隆DoAP3-3启动子,对其序列进行分析,发现含有多个转录起始位点核心元件,以及大量的光响应元件,脱落酸、赤霉素、生长素、水杨酸等植物激素的多个反应元件,分生组织表达相关调控元件,以及MYB转录因子结合位点等多个顺式作用元件。已有研究发现,延长光照时间会使墨兰花期延长,水杨酸影响花原基的淀粉含量,不同时期假鳞茎中ABA、GA3、IAA的高含量可能影响花序分化[14]。而寒兰假鳞茎中较高浓度的ABA/GA3、叶片中较低的ZR/IAA和ZR/GA3有利于花芽发育[15]。受光照、激素植物激素诱导的基因上游启动子区域内含多个激素感应因子结合位点[16]。结合前人研究,说明光照、植物激素以及MYB转录因子等可能通过与DoAP3-3启动子相互作用,调控该基因在分生组织表达,进而影响铁皮石斛的花形态建成,对于探究铁皮石斛花发育的表达调控途径具有重要意义,但具体的调控方式还需深入研究。
李泽卿[10]通过构建不同长度的二球悬铃木AP3的启动子表达载体,GUS染色程度随启动子缺失长度减少而增加。瞬时表达常用拟南芥或烟草作为转化植物[17,18],本研究构建长度为−1885~0、−1604~0、−750~0 bp 的 3 个启动子片段与GUS的融合表达载体,通过农杆菌介导对铁皮石斛原球茎进行瞬时表达,植物组织进行GUS化学染色,3个表达载体的转基因植物组织均有不同程度的染色,随着启动子缺失长度减少,GUS表达逐渐增加,推测−1885~−1604、−1604~−750 bp 区域含有增强子,但是未能被在线网站预测到,导致3个启动子片段均可驱动GUS基因表达,启动子片段活性强弱为−1885~0 bp>−1604~0 bp>−750~0 bp。
4 结论
铁皮石斛DoAP3-3基因的cDNA长度 675 bp,生物信息学分析预测其编码蛋白质具有碱性、亲水性和不稳定性,无跨膜结构域,亚细胞定位于细胞核中。克隆所得DoAP3-3的启动子受光激素等多种因子调控,具备启动活性,且随启动子缺失长度减少呈显增加趋势。
猜你喜欢
基层中医药(2021年3期)2021-11-22 08:08:08
数学大王·中高年级(2020年12期)2020-12-31 07:28:26
趣味(语文)(2020年4期)2020-07-27 01:43:14
亚热带植物科学(2019年3期)2019-10-16 05:21:24
中成药(2018年5期)2018-06-06 03:11:56
现代园艺(2018年2期)2018-03-15 08:00:05
现代工业经济和信息化(2016年2期)2016-05-17 05:34:16
广西林业科学(2016年3期)2016-03-16 05:43:37
电子工业专用设备(2015年4期)2015-05-26 09:10:40
汽车维修与保养(2015年8期)2015-04-17 03:33:01
