
西南印度洋龙旂热液区神盾螺共附生微生物多样性分析
2022-08-09 10:21牛圆圆何培青刘晨临
海洋科学进展 2022年3期
牛圆圆,何培青,刘晨临*
(1.自然资源部 第一海洋研究所海洋生物资源与环境研究中心,山东 青岛 266061;2.自然资源部 海洋生态环境科学与技术重点实验室,山东 青岛 266061)
深海热液区环境恶劣,黑暗、高压、低氧、温度梯度变化大、硫化物和重金属等有毒物质含量高[1],这种极端环境促使热液区动物形成了独特的生存机制,即与微生物的共附生关系。在深海热液区富含硫化物、氢气(H2)或甲烷(CH4)等,化能自养微生物利用这些还原性物质作为电子供体产生能量,并利用CO2合成有机物,为异养生物提供能源、碳源和其他营养物质,形成以化能合成为基础的生态系统[1]。大多数在热液区生活的无脊椎动物依赖这种初级生产力,以自由活动的细菌为食,或与化能合成细菌形成共生关系[2-4]。微生物以内共生(endosymbiont)或外共生(ectosymbiont)的方式分布在不同动物宿主的不同组织中,包括管状蠕虫的营养体、双壳类动物的鳃和腹足类动物的食道腺以及节肢动物(虾、蟹等)和软体动物的外壳表面等[4]。热液区生态系统中,微生物与地球化学环境、共生代谢和宿主生存之间存在着复杂的联系。微生物可以调节动物宿主与环境的相互作用,通过营养共生关系拓宽宿主动物的生态位,促进其生态和进化的多样性[5]。同时,热液区的动物也对其共生微生物具有高度选择性,同一属动物受环境化学条件和个体发育阶段的影响,也可能存在不同的宿主-共生关系[2,6]。
近年来,国内外科学家开展了盲虾[7]、贻贝[8]和管虫[9-10]等深海热液区动物共生微生物多样性、共生体能量利用及共生机制[11-13]等方面的研究。大部分热液区动物通过水平传播获得共生菌,即从周围环境获得自由生活的细菌,经过一个选择性的过程将其迁移到特殊的器官,内共生菌不再与外界环境直接接触。当动物死亡后,这些内共生菌被重新释放到环境中,营自由生活[9,12-14]。除了水平传播外,在深海热液蛤类中还发现了通过卵子从母代向子代垂直传播共生菌的情况[10]。
目前,在深海热液区已发现并报道了神盾螺属(Gigantopelta)的2 个种,分别为南大洋东斯科舍洋脊热液区的G.chessoia[15]和西南印度洋龙旂热液区的G.aegis[16],其共生菌主要分布于神盾螺的鳃和食道腺中[17]。印度洋G.aegis食道腺为双内共生菌系统,由γ-变形菌纲的硫氧化菌(Sulfide-Oxidizing Bacteria,SOB)和甲烷氧化菌(Methane-Assimilating Bacteria,MOB)同时为宿主提供能量[17]。南大洋G.chessoia膨大食道腺中主要分布硫氧化的γ-变形菌[18]。幼体G.chessoia(壳长≤7 mm)主要通过捕食细菌获得能量和营养,直到成体阶段才营内共生生活方式。与成体食道腺细菌组成相比,G.chessoia幼体细菌组成与成体鳃附着细菌组成更相似,包括多种γ-、ε-和δ-变形菌,这说明神盾螺可能通过水平传播获得共生菌[18]。
神盾螺共附生微生物是其与环境相互作用的重要媒介。以往研究结果表明,在南大洋和西南印度洋不同热液区生活的神盾螺共附生微生物的多样性存在差异[17],但在同一生境中的神盾螺不同个体间是否存在共附生微生物差异还未见报道。本文利用扩增子和宏转录组等高通量测序技术,分析西南印度洋龙旂热液区神盾螺G.aegis不同个体间共附生微生物多样性及其功能,为研究深海热液区化能合成微生物对腹足类动物宿主环境适应和海洋生态位分布的影响提供数据。
1 材料与方法
1.1 样品采集
2019 年4 月8 日中国大洋52 航次第三航段深海热液区底栖生物调查任务,利用“海龙Ⅲ”遥控水下机器人(Remote Operated Vehicle,ROV)在西南印度洋中脊龙旂低温热液区(49°38′59″E,37°42′01″S)采集获得神盾螺样品。采集样品后将其置于船载–80 ℃冰箱保存,以便在实验室进行后续分析处理。
1.2 实验试剂和仪器
PCR 扩增使用的酶Phusion®High-Fidelity DNA polymerase,New England Biolabs 公司,美国;PCR 扩增使用的缓冲液Phusion®High-Fidelity PCR Master Mix with GC Buffer,New England Biolabs 公司,美国;建库试剂盒Next®Ultra™ IIDNA Library Prep Kit,New England Biolabs 公司,美国;胶回收试剂盒,Qiagen 公司,德国;Trizol 试剂,Invitrogen 公司,美国。Bio-rad T100 梯度PCR 仪,Bio-Rad 公司,美国;测序上机仪器Illumina NovaSeq 6000,Illumina 公司,美国。北京诺禾致源生物信息科技有限公司于2021 年6 月22 日完成样品的DNA 提取和扩增子测序,2019 年10 月20 日完成RNA 提取和宏转录组测序。
1.3 扩增子测序及数据分析
1.3.1 基因组DNA 提取
利用液氮将3 个神盾螺个体(带壳)分别研碎,采用CTAB 法提取基因组DNA[19],并通过1%琼脂糖凝胶电泳检测DNA 纯度和浓度。
1.3.2 16S rRNA 基因V3-V4 扩增子测序
选用引物(正向引物515F:5′-GTGCCAGCMGCCGCGGTAA-3′,反向引物806R:5′-GGACTACHVGGG TWTCTAAT-3′)、Phusion®High-Fidelity PCR Master Mix with GC Buffer 和高保真酶PCR 扩增16S rRNA 基因V3-V4 区。PCR 的反应体系(30 μL)为:Phusion Master Mix(2×)15 μL,PrimerF(1 μmol/L)1 μL,PrimerR(1 μmol/L)1 μL,Genomic DNA(1 ng/μL)10 μL,最后使用ddH2O 补足30 μL 体系;PCR 的反应流程为:98 ℃预变性1 min;30 个循环(98℃,10 s;50 ℃,30 s;72 ℃,30 s);72 ℃,5 min。使用2%琼脂糖凝胶电泳检测PCR 产物,并根据PCR 产物浓度进行等量混样,充分混匀后再次使用2%琼脂糖凝胶电泳进行检测,然后采用胶回收试剂盒回收目的条带。使用Next®Ultra™ IIDNA Library Prep Kit 建库试剂盒构建文库,利用NovaSeq 6000 平台进行测序。
1.3.3 扩增子测序数据处理和分析
根据接头序列从下机数据中拆分出各样本数据,截去接头和引物序列后,使用FLASH(v1.2.11,http://ccb.jhu.edu/software/FLASH/)[20]进行拼接,得到原始序列(Raw Tags)。随后使用数据质控过滤软件Fastp 对得到的原始序列(Raw Tags)进行质控,得到高质量的序列(Clean Tags)。最后使用扩增子数据处理分析软件Vsearch(v2.15.0)将Clean Tags 与Silva 数据库进行比对检测并去除嵌合体[21],最终获得有效序列(Effective Tags)。
采用扩增子分析软件QIIME2(v202006)[22]中的DADA2 模块对有效序列进行降噪处理,并过滤掉丰度<5 的序列,最终获得扩增子序列变异(Amplicon Sequence Variants,ASVs)以及特征表。利用Classify-sklearn模块将获得的ASVs 与数据库比对从而获得每个ASV 的物种信息,并采用QIIME2 软件计算α 多样性指数[22]。
1.4 宏转录组测序及数据分析
1.4.1 样品RNA 提取与宏转录组测序
利用液氮分别研碎5 个神盾螺个体(带壳),并采用Trizol 提取样品的总RNA。检测RNA 的纯度、完整性和浓度后,从总RNA 中去除核糖体RNA,并将获得的mRNA 随机打断成250~300 bp 的短片段,以片段化RNA 为模板,随机寡核苷酸为引物合成cDNA 第一条链,随后按照链特异性建库方式进行建库。在Illumina HiSeqTM2000 平台进行测序,最终生成的序列为150 bp 的双端序列。
1.4.2 数据质量评估和转录本拼接
将测序获得的原始数据过滤掉低质量读长(reads)后,用转录组组装软件Trinity(v2.4.0)对获得的净读长(Clean Reads)进行拼接[23],输出可变剪切亚型的全长转录本,并梳理对应于旁系同源基因的转录本。在Trinity 拼接基础上,使用序列聚类软件CD-HIT-EST(v4.8.1)去冗余(设定序列一致性阈值为0.95),得到非冗余基因(unigene)集合[24]。然后,以神盾螺全基因组序列[17]为参照,进一步筛选掉宿主基因,获得共附生微生物宏转录组序列。
1.4.3 物种和基因功能注释
使用比对软件Diamond(v0.8.22)[25]对数据库中抽提出的细菌(Bacteria)、真菌(Fungi)、古菌(Archaea)和病毒(Viruses)序列进行比对(e-value≤1×e−5);采取系统分类软件MEGAN[26]的 LCA (Lowest Common Ancestor)算法,将出现第一个分支前的分类级别,作为该序列的物种注释信息;根据LCA 注释结果及基因丰度表,获得各个样品在各个分类层级上的丰度信息。
采用Diamond 软件将宏转录组序列与各功能数据库进行比对(Blastp,e-value≤1×e−5),进行对比的功能数据库包括:京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)、碳水化合物活性酶数据库(Carbohydrate-Active enZYmes Database,CAZy),以及基因的进化谱系:非监督直系群数据库(Evolutionary Genealogy of Genes:Non-supervised Orthologous Groups Database,eggNOG)等,基于NR 和Pfam 两部分的蛋白注释结果(e-value=1×e−6),确定宏转录组序列的基因功能。
2 结果与讨论
2.1 扩增子序列分析
通过16S rRNA 基因V3-V4 区扩增子测序,分别分析3 个神盾螺个体共附生菌的组成。3 个样品共扩增获得573 个ASVs,其中115 个ASVs 在3 个神盾螺个体中均有分布,138 个ASVs 仅分布于样品Ga3 中,而104 和73 个ASVs 分别为Ga2 和Ga1 的特有序列(图1)。3 个样品测序覆盖率(Goods Coverage)均为1,Ga3 低丰度物种较多,其总ASVs 最高为363 个,Ga2和Ga1 中的ASVs 分别为336 和247 个。Ga2 的香农-威纳指数(Shannon)和辛普森指数(Simpson)最高,说明其共附生菌的群落多样性和物种分布均匀度最高(表1)。将573 个共附生细菌ASVs 与西南印度洋神盾螺的SOB 及MOB 内共生菌基因组序列[17]比对,发现8 个ASVs 与参考序列具有较高相似性(98%)。其中,7 个ASVs 属于SOB,丰度最高的ASV0 与SOB 序列相似性为100%,在3 个样品中均有分布。丰度较低的1 个ASV(ASV244)属于MOB,仅在1 个样品中被检测到(表2)。
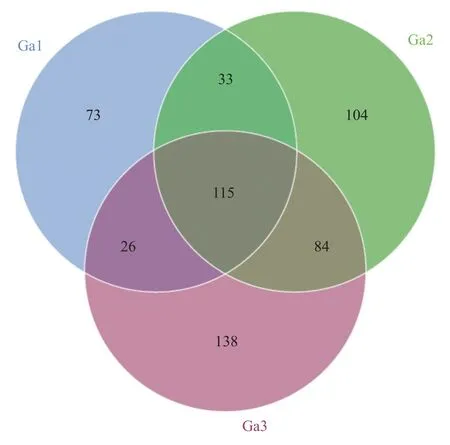
图 1 不同神盾螺个体ASV 组成的韦恩图Fig.1 Venn diagram of ASV composition in different individuals of Gigantopelta

表1 神盾螺共附生菌16S rRNA 基因V3-V4 区扩增子测序结果及alpha 多样性分析Table 1 Sequencing results of amplicon in V3-V4 region of 16S rRNA gene and analysis of alpha diversity of symbiotic and epiphytic microorganisms of Gigantopelta
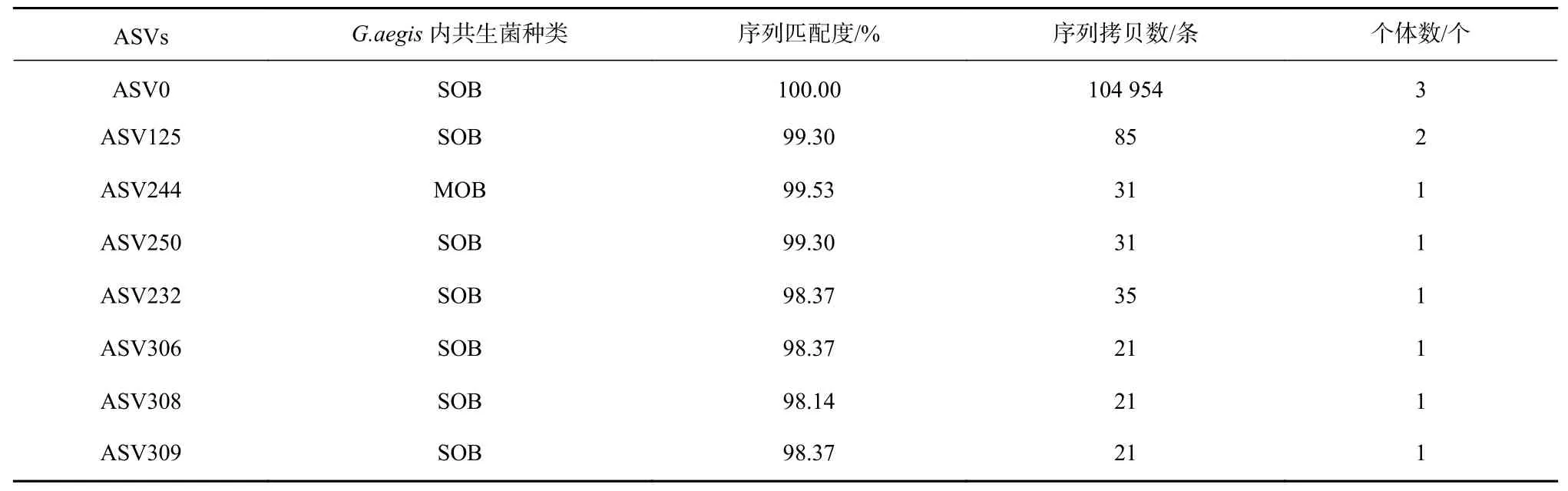
表2 与已报道的神盾螺的SOB 内共生菌和MOB 内共生菌基因组序列相似性较高的ASVsTable 2 ASVs with high similarity to the reported genome sequence of the sulfur-oxidizing endosymbionts and methano-oxidizing endosymbionts of Gigantopelta
测序获得的16S rRNA 基因V3-V4 区扩增子序列99.99%属于细菌域,主要包括10 个门和20 个纲(图2)。有125 个ASVs 属于变形菌门(Proteobacteria),在神盾螺共附生菌中丰度最高的包括γ-、ε-和α-变形菌纲,其中112 个ASVs 属于γ-变形菌纲。γ-变形菌纲在Ga1、Ga2 和Ga3 中的丰度分别为82.61%、 12.31%和81.25%。Ga1 和Ga3 的优势物种均为γ-变形菌纲Thiomicrospirales 目的SOB 内共生菌(ASV0),丰度分别为72.45%和67.60%。但是在Ga2 中ASV0 丰度仅为1.41%。此外,在3 个螺中γ-变形菌纲肠杆菌目(Enterobacterales)的Escherichia-Shigella属(ASV2)也有较高丰度,分别为5.69%、6.38%和7.53%。硫发菌目(Thiotrichales)、伯克氏菌目(Burkholderiales)、甲基球菌目(Methylococcales)和巴斯德氏菌目(Pasteurellales)的ASV 在3 个螺中平均丰度也均大于0.5%。

图2 基于16S rRNA V3-V4 区扩增子测序的神盾螺共附生细菌群落组成Fig.2 Community composition of symbiotic and epiphytic bacteria from Gigantopelta based on amplicon sequencing in 16S rRNA V3-V4 region
ε-变形菌纲在Ga1、Ga2 和Ga3 中的丰度分别为7.73%、68.73%和2.28%。有34 个ASVs 被鉴定为ε-变形菌纲弯曲菌目(Campylobacterales),其中24 个ASVs 来自硫卵形菌属(Sulfurovum),3 个ASVs 来自Nitratifractor属。Ga2 共附生优势菌(ASV1)为ε-变形菌纲的硫卵形菌属,其丰度达68.52%,而在Ga1 和Ga3 中ASV1 的丰度仅分别为7.65%和2.01%。与ASV1 序列相似性为100%的硫卵形菌属(Sulfurovum)在深海热液区生物和环境中广泛存在,如西南印度洋中脊热液区鳞足螺(Chrysomallon squamiferum)外壳附生菌(AY531602)[14]、大西洋热液盲虾(Rimicaris exoculata)肠道附生菌(FR839086)[27],以及深海喷口新暴露玄武岩表面生物膜细菌(KT257758)[5]等。
在鉴定获得的神盾螺共附生菌中,厚壁菌门(Firmicutes)种类最多,共包含196 个ASVs,其中62 个ASVs 属于乳杆菌目(Lactobacillales),丰度最高的是毛绒厌氧杆菌属(Lachnoanaerobaculum);另外,Lachnospirales目、Oscillospirales 目和消化链球菌科(Peptostreptococcaceae)的Romboutsia属也在神盾螺中均有分布。拟杆菌门(Bacteroidetes)中包含135 个ASVs,其中Muribaculaceae 科、Prevotella属和Aestuariimonas属丰度最高。梭杆菌门(Fusobacteria)的22 个ASVs 中,梭杆菌属(Fusobacterium)和纤毛菌属(Leptotrichia)丰度最高。脱硫杆菌门(Desulfobacterota)中含有11 个ASVs,脱硫叶菌属(Desulfobulbus)和脱硫弧菌属(Desulfovibrio)在神盾螺中均有分布,其中在Ga2 中丰度最高。放线菌门(Actinobacteria)含有30 个ASVs,代表属为糖多孢菌(Saccharopo-lyspora)和双歧杆菌(Bifidobacterium)(图2)。
在深海热液区,优势化能自养菌主要为γ-变形菌和ε-变形菌的SOB[28]。对3 个神盾螺个体的16S rRNA基因V3-V4 区扩增子测序结果表明,同一热液区不同个体的共附生菌种类差异较大。个体Ga1 和Ga3 的共附生菌以γ-变形菌为主,而个体Ga2 的以ε-变形菌为主。大西洋热液盲虾的鳃附着大量细菌,以ε-变形菌和γ-变形菌为主。它们氧化硫化氢(H2S)或氢气,以不同途径固碳,在减小附生菌间直接竞争的同时,也扩大了盲虾栖息的环境范围[7]。研究发现,南太平洋毛螺(Alviniconcha)鳃中同时分布着化能自养的γ-变形菌和ε-变形菌[2],中印度洋中脊热液区毛螺以ε-变形菌内共生菌为主[4]。这些研究结果与我们的发现相一致,深海热液区动物的共附生菌组成具有相似性,以γ-变形菌和ε-变形菌为主,但本文获得的同一区域生活的宿主的不同个体间的优势共附生菌组成存在差异的结果为首次报道。
尽管3 个神盾螺个体共附生菌的种类不同,但它们均属于SOB,反映了神盾螺生存的龙旂热液生态系统以SOB 为主要的化能自养初级生产力,SOB 在热液环境中普遍存在,神盾螺的共附生菌可以营自由生活,也可作为热液动物群的外共生和内共生菌[29]。Ga2 的优势菌,ε-变形菌也在热液区生境中大量存在,多以外共生形式生活于多毛类蠕虫、软体动物和甲壳动物上,很少以内共生形式存在[30]。ε-变形菌能够氧化硫、甲酸盐和氢产生能量,用于自身生存和维持共生关系[2,4]。
2.2 基于宏转录组序列的神盾螺共附生微生物多样性分析
将从5 个神盾螺个体中提取的RNA 混样后,进行宏转录组测序。原始数据经过滤后的得到88 876 988个读长(reads),共13.33G 的碱基数据量。Phred 数值大于20 和30 的碱基占总碱基的百分比Q20 和Q30 分别为97.07%和91.92%,GC 含量为42.76%。数据拼接后共得到108 537 个转录本,去冗余后得到95 408 条基因,其N50 和N90 长度分别为438 和228 bp。
在神盾螺共附生微生物宏转录组序列中,去除宿主序列后共鉴定出39 个门,包括1 个病毒门、2 个古菌门、29 个细菌门以及7 个真核生物门。细菌序列占宏转录组序列的45.48%,主要为变形菌门(65.00%)和拟杆菌门(28.49%);真核生物相对丰度为5.53%,主要为真菌,包括子囊菌门(Ascomycota,93.76%)和毛霉门(Mucoromycota,2.29%);古菌序列相对丰度较低(0.17%),主要由广古菌门(Euryarchaeota,30.10%)和奇古菌门(Thaumarchaeota,67.04%)构成(图3)。

图3 基于宏转录组序列的神盾螺共附生微生物多样性Fig.3 Symbiotic and epiphytic microbial diversity of Gigantopelta based on metatranscriptomic sequences
与扩增子测序结果基本一致,宏转录组测序结果表明变形菌门在神盾螺共附生细菌中相对丰度最高,包括γ-(27.79%)、ε-(17.92%)、α-(9.79%)、δ-(4.12%)、β-(3.01%)和ζ-1.09%变形菌纲。其中,ε-变形菌纲细菌相对丰度最高,以硫卵菌(14.11%)为主,硫化螺旋菌(Sulfurospirillum)、硫单胞菌(Sulfurimonas)和Nitratifractor也有较高丰度。γ-变形菌中肠杆菌目、甲基球菌目、Cellvibrionales 目和硫发菌目丰度较高。δ-变形菌中黏球菌目(Myxococcales)、脱硫单胞菌目(Desulfuromonadales)、脱硫弧菌目(Desulfovibrionales)和脱硫杆菌目(Desulfobacterales)丰度较高。拟杆菌门中,黄杆菌目(Flavobacteriales)的Lutibacter属(2.43%)和极地杆菌属(Polaribacter)(1.19%)丰度较高。放线菌门中拟孢囊菌属(Kibdelosporangium)(1.06%)和Lawsonella(1.08%)丰度较高。黏胶球形菌门(Lentisphaerae,4.47%)和梭杆菌门中嗜冷杆菌(Psychrilyobacter,1.35%)的丰度较高。
除细菌序列外,在宏转录组中还鉴定获得29 个属的真菌序列,其中子囊菌门的Phialocephala属丰度占95.48%,其次为芽枝霉门(Blastocladiomycota)和异水霉属(Allomyces)。据报道,Phialocephala为植物内共生真菌,能促进宿主生长,并通过产生次级代谢产物提高宿主抗病和抗采食能力[31]。Chen等[32]从一株深海沉积物真菌Phialocephala中分离获得新的倍半萜对苯二酚,对白血病细胞显示了很强的增殖抑制活性。Phialocephala与神盾螺是否存在互惠共生作用还有待进一步研究。
在古菌序列中,甲烷微菌纲(Methanomicrobia)丰度占54.79%,古丸菌纲(Archaeo-globi)古丸菌属(Archaeoglobus)占16.81%,甲烷暖球菌属(Methanocaldoco-ccus)占5.65%。目前,已知的古丸菌纲都为从海洋热液系统或海上油田中分离获得的严格厌氧菌和超嗜热菌。古丸菌属类群均能够还原亚硫酸盐和硫代硫酸盐[33]。甲烷暖球菌为最早从深海热液喷口中分离获得的嗜热化能自养微生物,拥有地球上最古老的呼吸代谢途径之一,即利用氢气和二氧化碳(CO2)产生甲烷获得能量[34-35]。
2.3 基于宏转录组的共附生微生物功能分析
2.3.1 代谢相关基因
1)硫代谢
深海热液区海水、热液羽流以及无脊椎动物的共附生微生物中存在多种SOB。硫化氢和硫化物氧化是深海热液区化能自养代谢过程中最主要的能量来源。除了有氧氧化,硫氧化还与硝酸盐还原相耦合[36]。
硫醌氧化还原酶(Sulfide:Quinone Oxidoreductase,SQR)是高浓度硫化物中SOB 生长的关键基因[37]。硫醌还原酶将硫化物不完全氧化成单质硫,防止完全氧化生成硫酸盐造成环境的酸化[38]。在大西洋盲虾鳃腔内,共生菌的硫醌还原酶基因不仅可以起到防止有害酸化的作用,而且在硫化物浓度较高时能够以硫的形式储存能量[7]。神盾螺宏转录组中含有36 条硫醌还原酶基因片段,与ε-变形菌纲的硫卵形菌属和Nitratifractor属,γ-变形菌纲发硫菌属(Thiothrix),以及拟杆菌门黄杆菌纲(Flavobacteriia)等的硫醌还原酶基因有较高相似性。
硫代硫酸盐氧化复合酶体系(Sox 多酶复合体系)参与氧化硫化物和亚硫酸盐生成硫酸盐的途径。从神盾螺宏转录组序列中获得28 条Sox 基因序列,亲缘关系最近的基因主要包括热液或冷泉环境中的γ-和ε-变形菌纲,如硫卵形菌属和亮发菌属(Leucothrix)等;也包括大西洋热液盲虾,黑烟囱热液喷口螺以及冷泉管虫共生菌(表2)。
异化亚硫酸盐还原酶(Dissimilatory sulfite reductase,Dsr)在厌氧呼吸过程中以硫酸盐、亚硫酸盐或有机磺酸盐为末端电子受体,催化亚硫酸盐还原为硫化物,并抑制硫化物氧化[39]。此外,Dsr 能够重新利用细胞内储存的硫元素,以维持在低浓度硫化物环境下的能量代谢[40-41]。与本研究Dsr 基因相似度最高的序列为γ-变形菌纲,其中发硫菌属最常见。3 条Dsr 基因序列与已报道的印度洋神盾螺SOB 内共生菌序列相似性高于90%,但未达到100%。由此可见,本样品中的SOB 与之前报道的神盾螺SOB 内共生菌可能属于不同的基因型[17]。
2)氢代谢
氢化酶催化氢的可逆氧化。我们在宏转录组中发现了20 条氢酶基因与其他不同热液环境中细菌的氢酶基因具有较高相似性,包括γ-变形菌纲、ε-变形菌纲和拟杆菌门等,如热液喷口羽流的着色杆菌科(Chromatiaceae)和Lutibacter属、深海热液羽流和烟囱中的硫卵形菌属、热液沉积物中的Nitratifractor属,以及大西洋盲虾鳃共生菌等。这表明神盾螺共附生微生物能够参与氢代谢以提供能量。
在多种深海热液生物共生菌中均发现了氢酶基因,包括管状蠕虫(Riftia pachyptila)和大西洋盲虾等[42]。研究表明,生活在富含氢气的大西洋中脊Logatchev 热液区深海贻贝共生菌能够利用氢气为能量源,生活在氢气浓度较低的太平洋深海热液区的贻贝共生菌也具有氢酶基因[43];在不同热液区的腹足类毛螺共生菌,都能同时以氢气和硫化氢为能量来源,随环境中氢气和硫化氢浓度比例的下降,氢酶的基因表达量和酶活也降低[4]。这说明利用氢为能源的生存方式在热液区共生关系中可能广泛存在,尤其是在氢含量高的区域[4,43]。
3)甲烷代谢
尽管16S rRNA 扩增子测序结果表明MOB 内共生菌丰度极低,但宏转录组序列中发现了甲烷代谢相关基因,包括四氢甲烷蝶呤水解酶(Fae)和甲烷单加氧酶(Pom)等基因。将4 条宏转录组Fae 序列与神盾螺MOB 内共生菌的Fae 序列(WP_198245971)和热液贻贝共生菌的Fae 序列(WP_174483451)进行比对,发现基因c20819_g1 与神盾螺的Fae 基因相似性较高,但仍有明显序列差异(图4)。将宏转录组中发现的3 条pomC 基因与其他热液动物共生菌的pomC 基因构建系统进化树(图4),发现其中2 条与不同热液环境的深海贻贝以及神盾螺内共生菌的基因聚为一个分支,另1 条序列则与来自甲基球菌目的细菌进化关系最近。说明本研究的神盾螺宏转录组序列除共附生菌外可能也包括来自环境中的细菌,这些细菌将甲烷氧化成二氧化碳,并利用中间产物甲醛作进行同化作用,为神盾螺提供碳源。
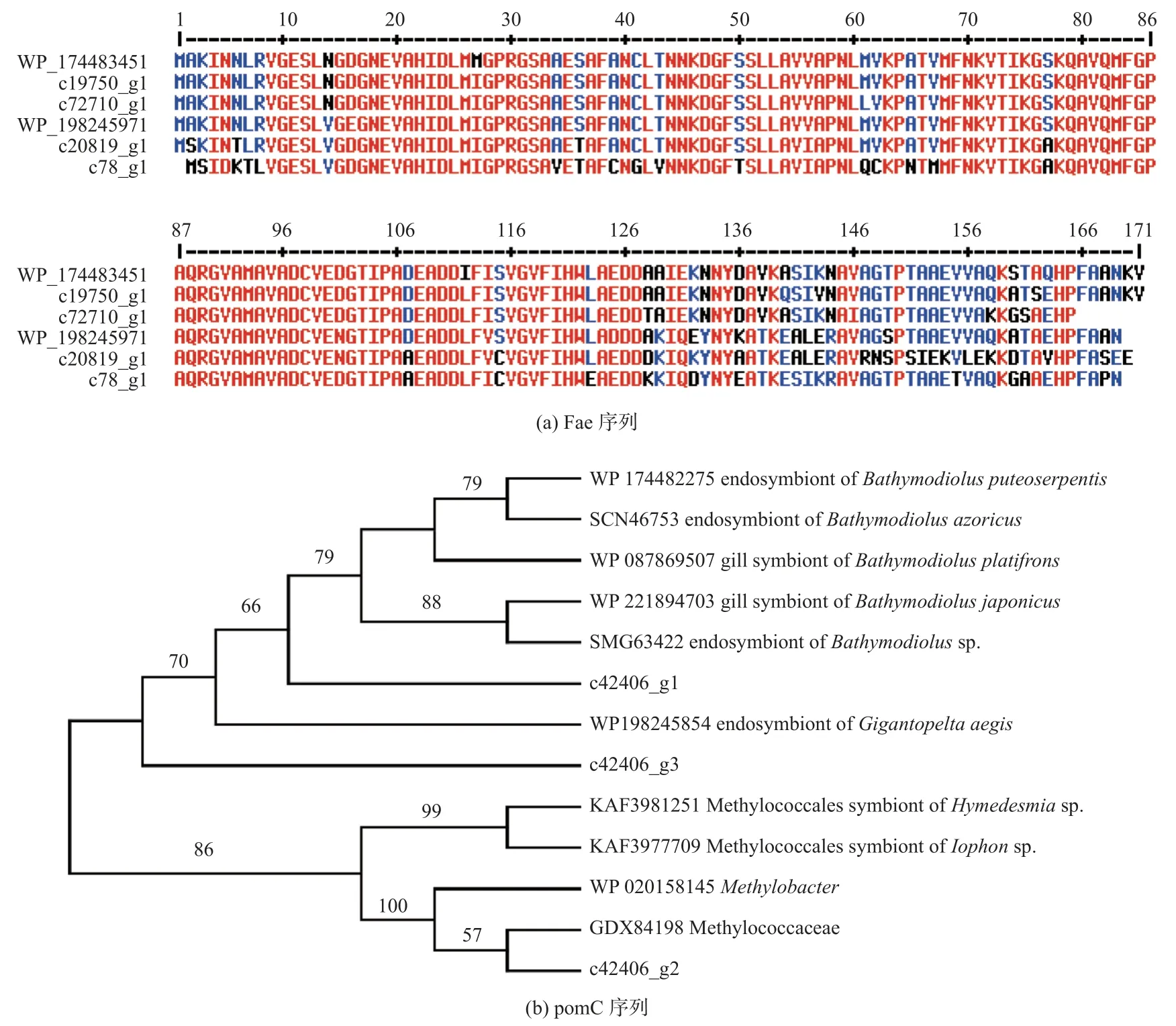
图4 神盾螺宏转录组中甲烷代谢基因与其他深海热液动物内共生菌参考序列的多重比对和系统进化分析Fig.4 Multiple alignment of methane-metabolizing genes with reference sequences of symbiotic bacteria from other deep-sea hydrothermal animals in the metatranscriptomic of Gigantopelta and Phylogenetic analysis
2.3.2 环境胁迫应答相关基因
Fis(Factor for Inversion Stimulation)是一种高效的全局转录调节因子,在不同生长阶段以及非生物胁迫时作出响应[44]。Fis 可以调控毒力、运动和代谢等生物学过程,调节各种毒力性状、胞外酶和胞外多糖的产生、细胞游动和群体运动、生物膜的形成和细胞聚集,以及群体感应系统等,涉及到100 多个相关基因表达的增强或抑制[45]。Fis 家族转录调控因子高度保守且在细菌基因组中通常只含有1 个拷贝,在细菌指数生长期间高表达,在稳定期几乎测不到表达[46]。在宏转录组中本研究发现了多条Fis 序列,与西南印度洋神盾螺SOB 内共生菌、海绵甲基球菌目共生菌、热液喷口亮发菌属,以及热液羽流的甲烷冷泉甲基球菌科(Methylococcaceae)和Methylomarinum属等序列相似性较高,说明这些微生物的近缘种在神盾螺共附生群落中有较高的活力。
在深海热液环境中含有大量有毒性重金属,微生物对重金属的抗性机制主要是通过各种金属外排ATP酶或化学渗透阳离子反向运输泵向外运输有毒离子[47]。在神盾螺宏转录组中发现了大量重金属抗性基因,包括7 条Cd2+/Zn2+外排ATP 酶CadA 和6 条Cu2+外排ATP 酶CopAB 基因。钴-锌-镉抗性蛋白(Cobalt-zinc-cadmium resistance protein,CzcA)是阳离子反向运输泵的核心,是由一段肽链编码的同三聚体,可将离子从细胞质转运出来[48],在细菌的重金属抗性方面发挥着重要作用。本研究发现了7 条czcA 基因,与来自硫卵形菌属,Lutibacter属和Flavobacterium属等物种的基因同源。
此外,在神盾螺宏转录组中还找到139 条编码转座酶(transposase)的基因。转座酶可以在基因组内或基因组间实现水平基因转移,在中大西洋海脊热液区的碳酸盐烟囱的生物膜宏基因组中,超过8%的宏基因组与转座酶序列相似,比在其他环境中的高一个数量级。通过转座酶实现的水平基因转移,使生物膜成员具有更高的表型多样性,以适应不利的生存环境[49]。宏转录组中大量的转座酶基因,说明神盾螺共附生菌通过基因水平转移,获得更好的环境适应性。
3 结语
采用16S rRNA 基因V3-V4 区扩增子和宏转录组测序方法分析了西南印度洋龙旂热液区神盾螺共附生微生物群落结构及生态功能。研究发现SOB 是神盾螺共附生微生物群落中的优势类群。扩增子测序表明,同一群落神盾螺不同个体间的优势菌的种类不同,优势类群分别为γ-变形菌纲和ε-变形菌纲。其中γ-变形菌纲代表种的序列与已报道的神盾螺SOB 内共生菌序列相似度为100%。ε-变形菌纲代表属为硫卵形菌属,为深海热液区广分布类群。宏转录组测序结果也表明,硫卵形菌属、Nitratifractor属、亮发菌属、Flavobacterium属和甲基球菌目等为神盾螺共附生微生物优势类群。除硫氧化途径外,宏转录组中还发现了参与氢氧化和甲烷的基因。与深海热液环境微生物或热液动物共生菌的序列相似度最高,说明这些微生物能够通过化能自养合成途径,为自身和宿主提供能量和碳源。因此,深海热液区动物个体共附生菌具有独特性,但其生态功能有一定的共性,反映了共附生关系的复杂性。
本研究为深海热液区无脊椎动物生存机制的进一步研究提供了理论参考。但是,由于样品量有限,同时缺乏实时环境理化参数,尚无法确定不同神盾螺个体共附生微生物组成对宿主能量获取的影响,也无法明确参与能量代谢过程的基因表达是否与环境中还原物质浓度密切相关。
猜你喜欢
小学生作文(低年级适用)(2020年10期)2020-11-10
浙江大学学报(理学版)(2019年5期)2019-10-16
福建基础教育研究(2019年2期)2019-09-10
福建基础教育研究(2019年2期)2019-05-28
中国金属通报(2019年9期)2019-01-03
军工文化(2017年12期)2017-07-17
学苑创造·A版(2017年4期)2017-05-13
学苑创造·A版(2017年5期)2017-05-09
