
不同龄期蠋蝽对黏虫幼虫的捕食作用
2022-08-08 03:56孙婧婧王孟卿张礼生彩万志
植物保护 2022年4期
孙婧婧, 王孟卿, 张礼生, 彩万志, 李 虎*
(1. 中国农业大学植物保护学院昆虫学系, 北京 100193; 2. 中国农业科学院植物保护研究所, 北京 100193)
黏虫Mythimnaseparata隶属鳞翅目Lepidoptera夜蛾科Noctuidae,不仅能为害小麦、玉米、水稻等粮食作物,还能为害蔬菜、果树、烟草等经济作物。幼虫食叶,暴发时吃光作物叶片、咬断麦穗和谷穗造成作物减产甚至绝产,是严重威胁我国农作物生产安全的重要害虫[1-3]。黏虫无滞育性,在适宜的环境下一年可发生多代,通常二代和三代黏虫为害较重;且其具有较强的迁飞能力,在我国每年至少有4次大范围迁飞危害,除新疆未见报道外,在其他地区均有分布[4-5]。因其具有广食性、群聚性、暴食性和迁飞性的危害特点,其每次暴发均会对我国农业造成重大损失。2012年,三代黏虫在我国东北、华北和西南地区大暴发,全国发生面积达938.3万hm2,造成的实际损失达99.2万t,2013年-2018年每年的发生面积均超过300万hm2[1,5-6]。长期以来,我国对黏虫的防治非常重视,1958年将其列为要消灭的10种害虫中的第二位,2020年9月又将其与草地贪夜蛾Spodopterafrugiperda、飞蝗Locustamigratoria等10种害虫列入一类农作物病虫害名录。目前我国治理黏虫的方法比较单一,主要依赖施用化学农药,但因其成虫具有较强的迁飞能力以及幼虫具有较强的移动能力,使仅靠化学农药往往难以达到理想的防治效果,同时农药的长期施用也易造成农药残留、害虫抗药性、杀伤天敌等问题。而利用天敌昆虫来防控黏虫,在控制害虫种群数量的同时还能保护有益昆虫,以及解决农药不当使用带来的害虫再猖獗的难题。另外,天敌昆虫除通过捕食或寄生达到直接控制害虫数量的目的外,还可以通过间接作用影响害虫的生存或生殖[7]。如陈美云等[8]研究发现叉角厉蝽Eocantheconafurcellata能干扰斜纹夜蛾Spodopteralitura的交尾、产卵、栖息等行为,使其产卵量减少的同时卵块结构与大小等均发生改变。因此,研究天敌昆虫对黏虫的生物防治措施对害虫的可持续治理具有重要意义。
蠋蝽Armacustos,隶属于半翅目Hemiptera蝽科Pentatomidae益蝽亚科Asopinae,又名蠋敌,若虫期即可捕食鳞翅目、鞘翅目和半翅目等多个目的农林业害虫,是一种捕食能力较强的天敌昆虫[9-11]。20世纪70年代我国就有研究利用人工饲养的蠋蝽以1∶18的益害比在林间释放其老龄若虫来防治榆紫叶甲Ambrostomaquadriimpressum幼虫,结果发现13 d后的捕食率达到61.9%,能有效控制榆紫叶甲的种群数量[12];高卓等[13]利用F3代蠋蝽以1∶15的益害比在甜菜地释放其2龄若虫防治甜菜夜蛾Spodopteraexigua,结果显示释放13 d后防治效果达到63.8%,能有效抑制甜菜夜蛾的种群增长。高强等[14]利用人工繁育的蠋蝽以1∶15的益害比在烟田释放其5龄若虫和成虫防治斜纹夜蛾幼虫,结果显示虫口减退率随处理时间而增高,15 d后趋于稳定,能有效控制斜纹夜蛾的种群数量。而通过捕食功能反应的方法,发现蠋蝽对草地贪夜蛾[15]、小菜蛾Plutellaxylostella[16]、马尾松毛虫Dendrolimuspunctatus、思茅松毛虫D.kikuchii、云南松毛虫D.houi[17]等多种农林害虫也具有较好的捕食潜力。黏虫作为蠋蝽的猎物之一,能满足蠋蝽生长发育与繁殖的各项营养需求[10],理论上能够利用蠋蝽来防治黏虫,然而关于蠋蝽对黏虫捕食能力的研究还未见报道。因黏虫1~2龄幼虫仅取食叶肉形成小孔,危害较小,3龄后幼虫危害剧增,可将作物取食成光秆,所以通常在黏虫1~3龄时进行防治。因此,本研究选取食量和活动能力较高的黏虫3龄幼虫,在室内条件下测试蠋蝽3~5龄若虫及成虫对黏虫3龄幼虫的捕食能力以及其自身密度对其捕食作用的影响,初步评价不同龄期蠋蝽对黏虫幼虫的控害潜能,明确蠋蝽防治黏虫的最适龄期,以期为田间应用蠋蝽防治黏虫提供理论依据。
1 材料与方法
1.1 材料
供试昆虫:黏虫于人工气候室利用云南恒草生物科技有限公司提供的人工饲料饲养多代,发育至3龄幼虫期供试。蠋蝽在人工气候室用黄粉虫Tenebriomolitor幼虫及蛹连续多代饲养,发育至3~5龄若虫和成虫期供试。
供试植物:玉米品种为‘曲辰19号’,利用营养土在人工气候室内培养至3叶期供试。供试昆虫及植物均于温度(25±1)℃,相对湿度(60±5)%,光周期 L∥D=14 h∥10 h 的人工气候室内饲养。
仪器:养虫盒(长21 cm、宽14 cm、高9 cm)为盒盖上切割下一长方块(长12 cm、宽8 cm)后用120目的尼龙网纱密封的透明塑料盒;塑料杯(杯底直径5.5 cm、杯口直径7.5 cm、高9.5 cm)容积为300 mL;QHX-250BSH-Ⅲ人工气候箱,上海新苗医疗器械制造有限公司。
1.2 方法
蠋蝽对黏虫3龄幼虫的捕食试验和蠋蝽自身密度对其捕食的干扰反应试验均在温度(27±1)℃、光周期L∥D=14 h∥10 h、相对湿度(70±5)%的人工气候箱内进行。
1.2.1蠋蝽对黏虫3龄幼虫的捕食试验
蠋蝽3龄若虫和4龄若虫对黏虫3龄幼虫的捕食试验:选取蜕皮24 h内的蠋蝽3龄若虫和4龄若虫分别单头放入塑料杯内,在杯底放置一块浸湿的约6 cm2大小的脱脂棉供蠋蝽补充水分,杯口用120目的尼龙纱网和橡皮筋密封,置于温度(27±1)℃、光周期 L∥D=14 h∥ 10 h、相对湿度(70±5)%的人工气候箱内饥饿24 h。根据预试验的结果,对每头蠋蝽3龄若虫和4龄若虫设置的黏虫3龄幼虫密度梯度为3、6、9、12、20头/盒,按设计的密度将黏虫3龄幼虫和1头饥饿24 h后的蠋蝽若虫放入养虫盒内进行试验,每个养虫盒内放有2片新鲜玉米叶和适量人工饲料供黏虫幼虫取食。每个密度为1个处理,每个养虫盒为1个重复,每个处理重复10次。24 h后检查养虫盒内剩余黏虫幼虫的存活数,以计算蠋蝽若虫在不同猎物密度下的日捕食量以及日捕食率。日捕食量=(养虫盒内原有黏虫3龄幼虫数量-养虫盒内剩余黏虫3龄幼虫活虫数量)/1 d,日捕食率=日捕食量/养虫盒内原有黏虫3龄幼虫数量×100%。
蠋蝽5龄若虫和雌、雄成虫对黏虫3龄幼虫的捕食试验:除选取蜕皮24 h内的蠋蝽5龄若虫和羽化24 h内的蠋蝽雌、雄成虫,黏虫3龄幼虫密度梯度设为5、10、15、20、40 头/盒外,其他步骤均与蠋蝽3龄若虫和4龄若虫对黏虫3龄幼虫的捕食试验相同。
1.2.2蠋蝽对黏虫3龄幼虫的捕食功能反应
蠋蝽3~5龄若虫及成虫对黏虫3龄幼虫的捕食功能反应,根据1.2.1的试验结果拟合Holling Ⅱ圆盘方程:Na=aNTr/(1+aThN)[18]进行分析。其中Na为蠋蝽捕食黏虫3龄幼虫的头数,a为蠋蝽对黏虫3龄幼虫的瞬间攻击率,N为每盒放置的黏虫3龄幼虫的密度,Tr是试验的总时间(取1 d),Th是蠋蝽对黏虫3龄幼虫的处理时间,单位为d(即蠋蝽处理1头黏虫3龄幼虫所用的时间)。瞬间攻击率a和处理时间Th是评价天敌对猎物捕食能力的重要参数,1/Th为天敌对猎物的理论日最大捕食量,a/Th为天敌对猎物的控害效能。
1.2.3蠋蝽对黏虫3龄幼虫的搜寻效应
蠋蝽3~5龄若虫及成虫对黏虫3龄幼虫的搜寻效应利用1.2.2拟合Holling Ⅱ圆盘方程的2个参数“a”和“Th”,代入搜寻效应方程S=a/(1+aThN)[19]进行拟合分析。S为搜寻效应,N为黏虫3龄幼虫的密度。
1.2.4蠋蝽自身密度对其捕食的干扰反应
蠋蝽3龄若虫和4龄若虫自身密度对其捕食的干扰反应试验:选取蜕皮24 h内的蠋蝽3龄若虫和4龄若虫饥饿处理24 h(方法同1.2.1)。根据预试验的结果,将黏虫3龄幼虫和饥饿后的蠋蝽若虫按20∶1、20∶2、20∶3、20∶4、20∶5的密度接入养虫盒内,养虫盒内放入2片新鲜玉米叶和适量人工饲料供黏虫幼虫取食。每个密度为1个处理,每个养虫盒为1个重复,每个处理重复5次,24 h后检查养虫盒内剩余黏虫幼虫的存活数,计算蠋蝽若虫在不同自身密度下的日捕食量,计算方法同1.2.1。
蠋蝽5龄若虫和雌、雄成虫自身密度对其捕食的干扰反应试验:除选取蜕皮24 h内的蠋蝽5龄若虫和羽化24 h内的蠋蝽雌、雄成虫,黏虫3龄幼虫和饥饿24 h后的蠋蝽按40∶1、40∶2、40∶3、40∶4、40∶5的密度接入养虫盒内外,其他步骤均与蠋蝽3龄若虫和4龄若虫自身密度对其捕食的干扰反应试验相同。
蠋蝽自身密度对捕食的干扰效应采用Watt干扰模型:A=aP-b[20]进行分析。A为平均每只蠋蝽捕食黏虫3龄幼虫的数量,a为蠋蝽的最大捕食量,b为天敌昆虫蠋蝽之间的干扰系数,P为蠋蝽密度。
1.3 数据分析
利用 Excel 2019 处理数据,使用 Graph Pad 6.01 拟合Holling Ⅱ圆盘方程、搜寻效应方程和Watt干扰模型并作图,利用Tukey’s法进行差异显著性检验。
2 结果与分析
2.1 蠋蝽对黏虫3龄幼虫的捕食量
蠋蝽3~5龄若虫及雌、雄成虫均可捕食黏虫3龄幼虫。在试验设置的黏虫幼虫密度梯度范围内,不同虫态蠋蝽对猎物的日捕食量均随着黏虫幼虫密度增加而显著增加。在相同的猎物密度下,蠋蝽对黏虫幼虫的日捕食量随其龄期的增加而增加,且雌成虫的日捕食量大于雄成虫。如当猎物密度为20头/盒时,蠋蝽3~5龄若虫及雌、雄成虫对黏虫的日捕食量分别为3.4、6.6、7.2、8.6头和7.5头。而同一虫态蠋蝽在不同猎物密度下的日捕食量存在显著差异。当猎物密度达到6~20头/盒,蠋蝽3龄若虫的捕食量趋于稳定,捕食率低于50.0%,为17.0%~40.0%。当猎物密度达到9~20头/盒时,蠋蝽4龄若虫的捕食量趋于稳定,捕食率低于50.0%,为33.0%~47.8%(表1)。对于蠋蝽5龄若虫及雄成虫,当猎物密度达到15头/盒时,蠋蝽5龄若虫及雄成虫对猎物的捕食率低于50.0%,分别为43.3%和44.0%。而对于蠋蝽雌成虫,当猎物密度达到20头/盒时,蠋蝽雌成虫对猎物的捕食率低于50.0%,为43.0%(表2)。
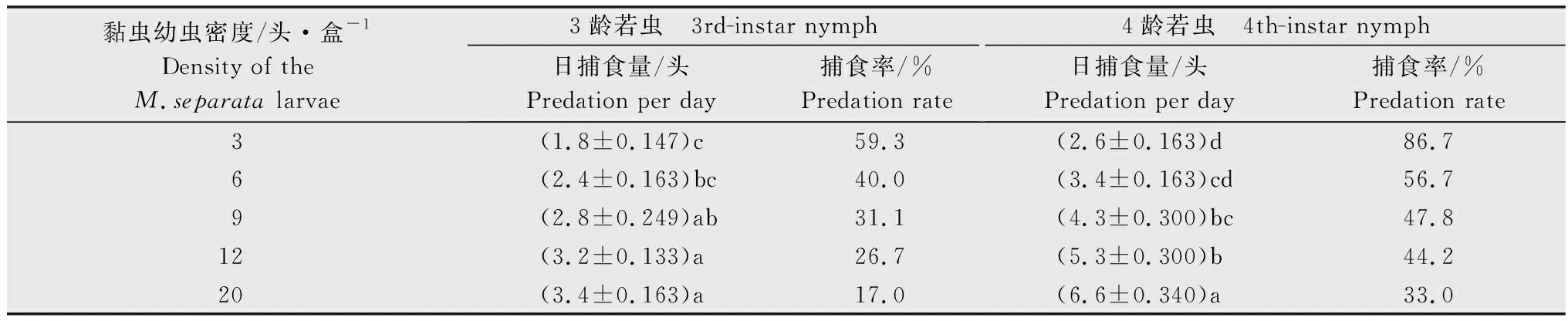
表1 蠋蝽3龄和4龄若虫对不同密度黏虫3龄幼虫的捕食效果1)Table 1 Predation effect of the 3rd-instar and 4th-instar nymphs of Arma custos against the 3rd-instar larvae of Mythimna separata
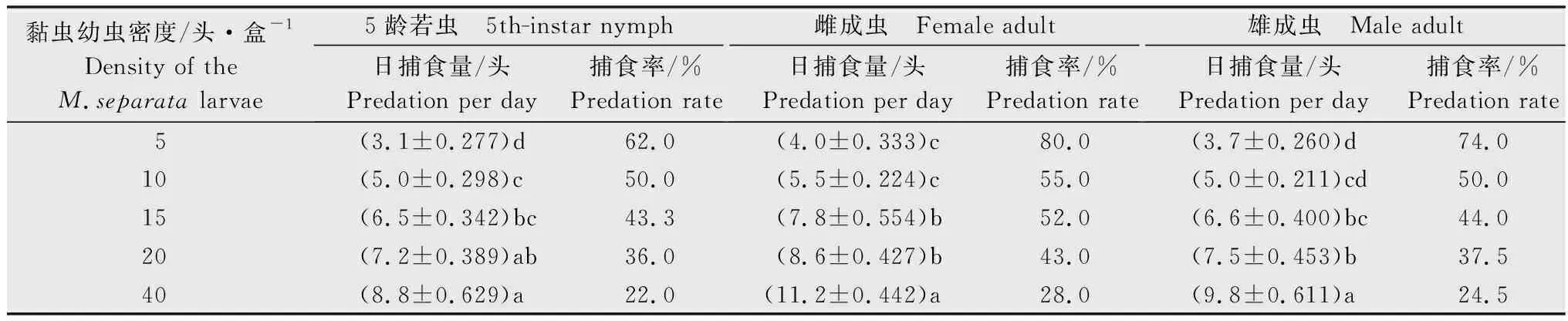
表2 蠋蝽5龄若虫和成虫对不同密度黏虫3龄幼虫的捕食效果Table 2 Predation effect of the 5th-instar nymphs and adults of Arma custos against the 3rd-instar larvae of Mythimna separata
2.2 蠋蝽对黏虫3龄幼虫的捕食功能反应
蠋蝽3~5龄若虫及雌、雄成虫对黏虫3龄幼虫的捕食量均随猎物密度的增加而增加,但曲线上升趋势逐渐减缓(图1),捕食功能反应均符合Holling Ⅱ模型(表3)。不同虫态蠋蝽对黏虫3龄幼虫的瞬时攻击率顺序为:3龄若虫(1.022)>雌成虫(0.961)>4龄若虫(0.933)>5龄若虫(0.892)>雄成虫(0.872),处理时间顺序为:雌成虫(0.063 d)<雄成虫(0.074 d)<5龄若虫(0.084 d)<4龄若虫(0.100 d)<3龄若虫(0.242 d),理论日最大捕食量顺序为:雌成虫(15.873头)>雄成虫(13.514头)>5龄若虫(11.905头)>4龄若虫(10.000头)>3龄若虫(4.132头)。从控害效能a/Th来看,蠋蝽雌成虫对黏虫3龄幼虫的控害效能最大,为15.254,3龄若虫的控害效能最小,为4.223,顺序与理论日最大捕食量顺序相同(表3)。
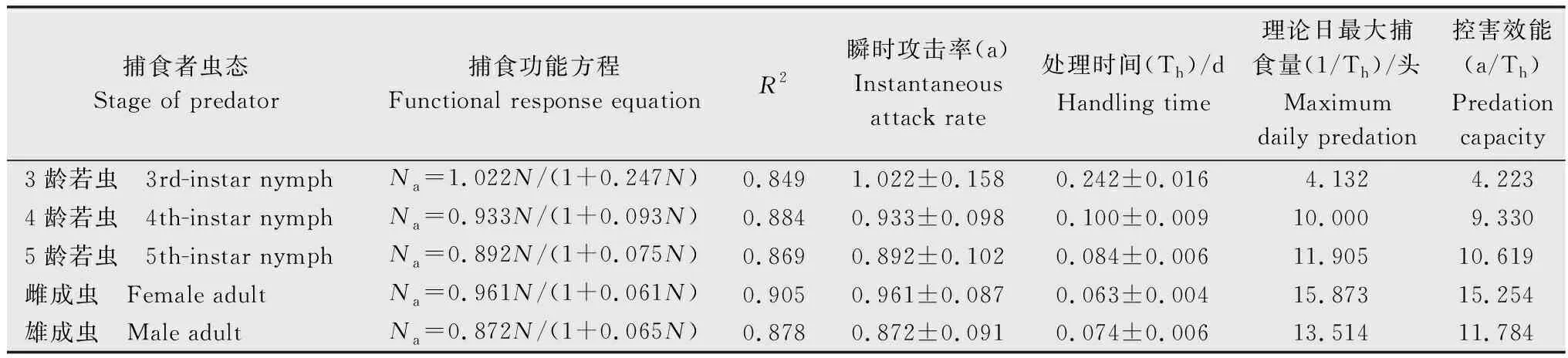
表3 蠋蝽对黏虫3龄幼虫的捕食功能反应1)Table 3 Predatory functional responses of Arma custos to the 3rd-instar larvae of Mythimna separata
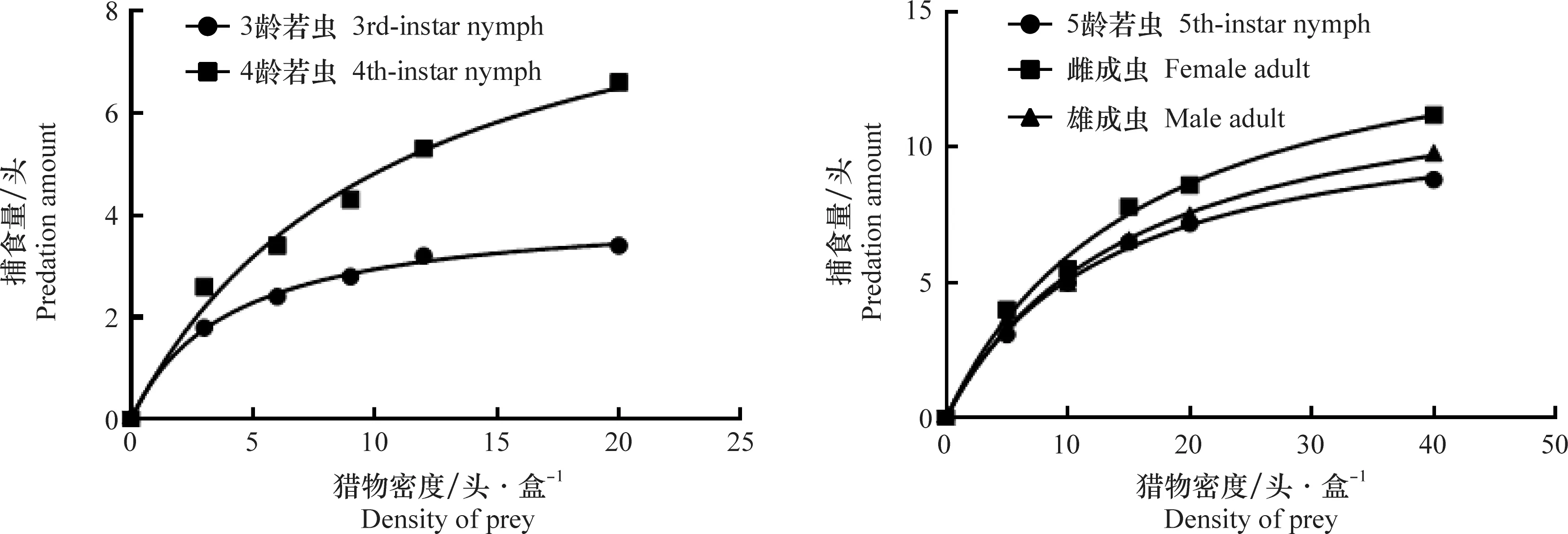
图1 蠋蝽3、4、5龄若虫和雌、雄成虫对黏虫3龄幼虫的捕食功能反应Fig.1 Predatory functional responses of the 3rd-instar, 4th-instar, 5th-instar nymphs and female, male adults of Arma custos to the 3rd-instar larvae of Mythimna separata
2.3 蠋蝽对黏虫3龄幼虫的搜寻效应
蠋蝽3~5龄若虫及雌、雄成虫对黏虫3龄幼虫的搜寻效应均随着猎物密度的增加而降低(图2)。在黏虫密度为3头/盒时,蠋蝽3龄若虫和4龄若虫的搜寻效应最高,分别为0.586和0.728;在黏虫密度为20头/盒时,蠋蝽3龄若虫和4龄若虫的搜寻效应最低,分别为0.172和0.324。在黏虫密度为5头/盒时,蠋蝽5龄若虫和雌、雄成虫的搜寻效应最高,分别为0.648、0.737和0.658;在黏虫密度为40头/盒时,蠋蝽5龄若虫和雌、雄成虫的搜寻效应最低,分别为0.223、0.280和0.242。

图2 蠋蝽3、4、5龄若虫和雌、雄成虫对黏虫3龄幼虫的搜寻效应Fig.2 Searching efficiency of the 3rd-instar, 4th-instar, 5th-instar nymphs and female, male adults of Arma custos to the 3rd-instar larvae of Mythimna separata
在相同的猎物密度下,蠋蝽4龄若虫对猎物的搜寻效应高于3龄若虫,蠋蝽雌成虫对猎物的搜寻效应高于雄成虫及5龄若虫。在黏虫密度为20头/盒时,不同虫态蠋蝽对猎物的搜寻效应顺序为:雌成虫(0.433)>雄成虫(0.379)>5龄若虫(0.356)>4龄若虫(0.324)>3龄若虫(0.172),可以看出蠋蝽对猎物的搜寻效应随龄期的增加而增大。
2.4 蠋蝽自身密度对其捕食作用的影响
在空间一定和黏虫幼虫密度固定的情况下,随着蠋蝽自身密度从1头/盒增加到5头/盒,其3~5龄若虫及雌、雄成虫对黏虫3龄幼虫的日平均捕食量不断降低,3龄若虫的日平均捕食量由3.80头逐渐下降至2.28头,4龄若虫的日平均捕食量由7.40头逐渐下降至3.60头,5龄若虫的日平均捕食量由8.60头逐渐下降至4.44头,雌成虫的日平均捕食量由10.40头逐渐下降至2.84头,雄成虫的日平均捕食量由8.80头逐渐下降至3.12头(表4)。
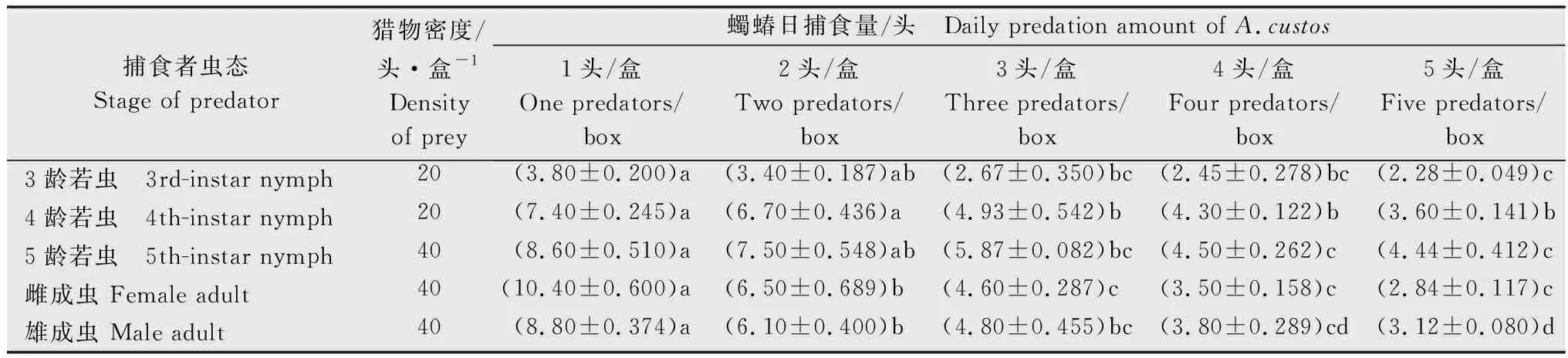
表4 不同密度的蠋蝽对黏虫3龄幼虫的日捕食量1)Table 4 Daily predation capacity of the 3rd-instar larvae of Mythimna separata by Arma custos with different density
蠋蝽3~5龄若虫和雌、雄成虫的自身密度干扰方程分别为A=3.904P-0.321(R2=0.943)、A=7.755P-0.408(R2=0.899)、A=8.935P-0.413(R2=0.920)、A=10.510P-0.767(R2=0.995)和A=8.911P-0.600(R2=0.991)。蠋蝽的干扰系数随其龄期的增加而增大,且雌成虫的干扰系数大于雄成虫,3~5龄若虫和雌、雄成虫的干扰系数分别为0.321、0.408、0.413、0.767和0.600。还可看出,不同虫态蠋蝽在无干扰(密度为1头/盒)时的日捕食量与蠋蝽对黏虫幼虫的理论最大捕食量(分别为3.90、7.76、8.94、10.51头和8.91头)相近。
3 讨论
明确天敌昆虫对害虫捕食能力大小是评价其是否具有生防应用价值的首要基础,而捕食功能反应是测试天敌昆虫对目标害虫捕食能力的经典方法[18,21-22]。本研究结果显示,蠋蝽3~5龄若虫及雌、雄成虫对黏虫3龄幼虫的捕食功能反应符合Holling Ⅱ模型,这与蠋蝽对其他多种农林业害虫的捕食功能反应模型一致[11,15,17]。在本试验中,蠋蝽对黏虫幼虫的捕食能力随其龄期的增大而增加,雌、雄成虫和5龄若虫对黏虫的控害潜能相对较高,实际日最大捕食量顺序为:雌成虫(11.2头)>雄成虫(9.8头) >5龄若虫(8.8头) >4龄若虫(6.6头) > 3龄若虫(3.4头),与理论日最大捕食量顺序相同:雌成虫(15.873头)>雄成虫(13.514头) >5龄若虫(11.905头) >4龄若虫(10.000头) > 3龄若虫(4.132头)。这与蠋蝽捕食斜纹夜蛾、小菜蛾和马尾松毛虫等鳞翅目害虫幼虫的试验结果一致,即雌、雄成虫和5龄若虫对害虫的捕食能力相对较高[14,16-17,23]。但在蠋蝽捕食荔枝蝽Tessaratomapapillosa1龄若虫的试验中,蠋蝽4龄若虫对猎物的日最大捕食量(9.066头)明显高于5龄若虫(8.210头)和雌(8.354头)、雄成虫(5.903头),即蠋蝽4龄若虫对猎物表现出相对较高的捕食能力[11];在蠋蝽捕食枸杞棉蚜Aphisgossypii的试验中,蠋蝽3龄若虫对猎物的捕食量和搜寻能力也显著大于4龄、5龄若虫和成虫,即蠋蝽3龄若虫对猎物表现出相对较高的捕食能力[24]。这些差异可能与猎物类型、个体大小及营养有关[25-26],与鳞翅目幼虫相比,荔枝蝽1龄若虫和枸杞棉蚜体型较小、营养较差,虽能满足蠋蝽3龄若虫期和4龄若虫期生长发育的营养需求,但难以满足其5龄若虫期羽化和成虫期繁殖的营养需求,因此蠋蝽5龄若虫和成虫对这两种害虫的捕食潜能相对3龄和4龄若虫较低。这些差异还表明了蠋蝽对不同猎物的捕食能力并非均随其龄期的增大而增加,应用蠋蝽进行害虫生物防治时,要针对目标害虫的种类选择适宜的蠋蝽释放龄期。本试验结果表明,蠋蝽雌、雄成虫对黏虫幼虫的日捕食量最高、处理时间最短及搜寻能力最强,理论上应选择释放蠋蝽成虫来防治黏虫幼虫,但李发倩等[27]通过盆栽试验发现在植株上蠋蝽若虫的捕食积极性高于成虫。在捕食黏虫的过程中,蠋蝽5龄若虫对猎物的捕食能力与成虫接近,但受到的干扰作用却相对于成虫较弱,且同一时期释放的5龄若虫在田间的控害时间相对成虫较长,与成虫相比若虫无翅、不易逃逸。因此,为提高天敌利用率和节约成本,可在黏虫发生期考虑释放蠋蝽5龄若虫进行防治。
天敌对猎物捕食能力的大小,可以用控害效能(a/Th)来衡量[21-22,28]。在本试验中,蠋蝽雌成虫对黏虫3龄幼虫的控害效能最高,为15.254,与蠋蝽雌成虫对斜纹夜蛾3龄幼虫的控害效能(18.205)相近[23],虽小于蠋蝽对草地贪夜蛾3龄幼虫(63.588)、4龄幼虫(29.000)和小菜蛾4龄幼虫(90.500)的控害效能[15-16],但远大于蠋蝽成虫对马尾松毛虫3龄幼虫(3.293)、思茅松毛虫3龄幼虫(2.578)和云南松毛虫3龄幼虫(2.678)的控害效能[17]。这些差异可能与蠋蝽的捕食偏好性有关,即相较体表覆有毒毛的松毛虫,蠋蝽更喜捕食黏虫、斜纹夜蛾、草地贪夜蛾和小菜蛾等体表相对光滑的夜蛾科害虫,因此对其控害能力更高。与黏虫的其他捕食性天敌相比,蠋蝽对其控害效能与益蝽Picromeruslewisi5龄若虫对黏虫3龄幼虫的控害效能(12.902)相近[29],但低于叉角厉蝽成虫对黏虫3龄幼虫的控害效能(70.937)[30]。这些差异除与天敌昆虫对害虫的捕食能力有关外,还可能受到试验环境以及试验所设猎物密度梯度的影响;所以,采用(a/Th)指标来分析天敌对不同猎物控害效能时需慎重。有研究表明,当鳞翅目害虫为混合龄期时,叉角厉蝽几乎不捕食其1~2龄幼虫,选择捕食3龄及以上个体较大的中高龄猎物[31-33],而蠋蝽则对鳞翅目害虫1~2龄猎物具有较高的捕食效果[13-14],因此,建议在黏虫发生期,尤其是3龄前,选择释放蠋蝽5龄若虫进行防治;但当田间黏虫世代重叠严重时,为提高防治效果可考虑将蠋蝽与叉角厉蝽联合释放。
本研究结果表明,不同虫态蠋蝽对黏虫幼虫的搜寻效应随着猎物密度的增加而降低;当空间与猎物密度固定时,随着天敌密度增加天敌间的干扰作用不断增强,不同虫态蠋蝽对猎物的平均捕食量也不断降低。这与大红犀猎蝽Sycanusfalleni捕食草地贪夜蛾3龄幼虫[21]、东亚小花蝽Oriussauteri捕食草地贪夜蛾1龄幼虫[22]、叉角厉蝽捕食斜纹夜蛾3龄幼虫和黏虫3龄幼虫[30,33]等捕食性天敌对猎物的捕食试验结果一致。因此,为达到最佳防治效果,在应用蠋蝽防控黏虫时不仅需要考虑田间害虫密度,还需要考虑天敌间的干扰对捕食作用的影响,在大规模应用前进行田间试验,以制定最适的释放益害比及释放方案。且还有研究结果表明,捕食者对猎物的捕食作用不仅受到猎物密度和自身密度的影响,还受到作物种类及种植密度的影响。如Clercq等[34]发现,黑刺益蝽Podisusnigrispinus在甜椒和茄子上对甜菜夜蛾幼虫的捕食功能反应符合Holling Ⅱ模型,在番茄上则为Holling Ⅲ模型,认为是番茄叶片表面的纤毛干扰了捕食者的搜寻效率和捕食能力。陈雪梅等[35]发现,烟草植株密度对叉角厉蝽捕食斜纹夜蛾的捕食效率具有负效应,而叉角厉蝽的释放数量对其捕食效率具有显著的正效应。因此在应用蠋蝽来防治害虫时在考虑其对目标害虫的捕食能力的同时,还要考虑释放时作物种类、种植密度、植株大小等问题。
蠋蝽作为一种适应性较强的天敌昆虫,能够在释放地定殖形成稳定种群达到持续控制害虫种群密度的目的,是一种值得开发的捕食性天敌资源[13]。本研究在室内条件下初步探究了蠋蝽3~5龄若虫及雌、雄成虫对黏虫3龄幼虫的捕食能力,明确了蠋蝽5龄若虫和雌、雄成虫对黏虫幼虫具有较好的控害潜能。然而与环境条件稳定的室内相比,农业生态系统中的寄主植物、气候因子、环境条件、其他生物等因素均会影响到蠋蝽对黏虫的捕食效果。因此在应用蠋蝽防控害虫前,还需进一步进行田间试验,研究天敌对目标害虫的捕食效果以及天敌的释放技术。
猜你喜欢
农业工程学报(2022年10期)2022-08-22
天津农业科学(2022年5期)2022-05-31
现代农业科技(2022年9期)2022-05-19
植物保护(2021年4期)2021-11-12
云南农业(2021年11期)2021-11-12
饲料博览(2020年11期)2020-12-30
今日农业(2020年13期)2020-12-15
云南农业科技(2020年5期)2020-10-14
今日农业(2019年13期)2019-08-12
今日农业(2019年10期)2019-06-26
