
甘蓝型油菜油脂合成相关基因的动态表达分析
2022-08-06 06:14王衍坤沈怡范佳琪陈思旭夏法刚季彪俊
种子科技 2022年12期
王衍坤,沈怡,范佳琪,陈思旭,夏法刚,季彪俊
(福建农林大学农学院,福建 福州 350002)
甘蓝型油菜(下称“油菜”)是我国重要的油料作物之一,在世界上是仅次于大豆的油料作物[1]。油菜在为人类提供食用油的同时,也可以作为动物饲料、生物燃料等为人类生产生活提供丰富的物质材料,因此油菜的种植与研究对人类具有重要意义[2]。种子含油量是油菜最重要的品质性状之一,对油菜种子含油量进行遗传改良,可以满足人们的食用需求。不同油菜种子材料之间的含油量差别明显[3],提高油菜种子含油量,一直以来都是油菜育种工作者的研究重点和热点。
目前,在油菜种子含油量的遗传分析和QTL 定位[4]、调控油脂合成关键基因的功能研究、油脂合成相关基因的表达和代谢调控等方面取得了丰硕的研究成果。虽然油脂合成的代谢通路已经研究透彻,但影响油菜种子含油量和调控油脂合成的因素多种多样[5-6],阐释这些因素对油脂合成调控的机理,有助于加快油菜高含油量分子育种的进程。在种子发育的过程中,许多基因在不同阶段所发挥的作用也不尽相同,因此,分析种子发育过程中油脂合成相关基因的动态表达水平以及在蛋白质组水平上的代谢调控,有利于系统地解析油脂的合成过程,进一步了解含油量的决定因子。
本试验利用转录组测序技术,重点分析油菜种子不同发育时期油脂合成相关基因的动态表达水平,有利于加深对油菜油脂合成过程的了解,为后期高含油量油菜品种的选育以及转录组测序技术在油菜分子遗传上的有效运用提供有力帮助。
1 材料与方法
1.1 材料
甘蓝型油菜“中9”为研究对象,取发育中(授粉后15 d)和成熟期种子作为试验材料,每个材料3 次重复。
1.2 RNA 提取、文库构建和高通量测序
剥去种皮,只保留种子,用于RNA 提取。种子剥离后放入液氮冷冻,于-80 ℃保存。利用总RNA 提取试剂盒提取RNA,并于70%乙醇中-20 ℃长期保存。建库和测序由北京诺禾致源科技股份有限公司完成。
1.3 差异基因分析
测序的下机数据使用FastQC 进行质控,符合分析标准的测序数据转而使用Trimmomatic 软件进行过滤,以去除低质量的Reads(Trimmomatic 软件参数如下:LEADING:3TRAILING:3SLIDINGWINDOW:4:15MINLEN:30)。使用STAR 软件,将过滤后的reads 比对到油菜参考基因组序列上。接着,使用RSEM软件对各个样本进行转录本定量。使用定量分析的结果生成矩阵,构造DESeq2 的输入文件,进行两个组合之间的差异表达分析。P≤0.05 且|log2foldchange|>1 的基因被认为存在差异表达。
1.4 GO 分析
通过基迪奥生信分析平台(https://www.omicshare.com/tools/)的GO 富集分析模块实现差异表达基因的GO 富集分析(P<0.05)。
1.5 KEGG 分析
使用KOBAS 3.0 软件分析KEGG 通路中差异表达基因的统计富集,以P 调整值作为阈值。
2 结果与分析
2.1 差异表达基因分析
对油菜种子成熟期和发育期表达基因进行差异表达基因的筛选,共筛选得到油菜种子成熟期相对于发育期差异表达基因总数为36 294 个,其中上调转录基因数量为13 423 个,下调转录基因数量为22 871 个。
2.2 差异表达基因GO 注释分析
通过对成熟期种子表达上调的基因进行GO 注释(如图1 所示)可知,生物过程中富集到基因条目数量最多的依次为代谢过程(GO:0008152,3 435 个基因)、细胞过程(GO:0009987,3 250 个基因)、单有机体过程(GO:0044699,2 091 个基因)等。细胞组件富集到基因条目数量最多的依次为细胞(GO:0005623,1 827个基因)、细胞部分(GO:0044464,1 827 个基因)等。分子功能富集到基因条目数量最多的依次为绑定(GO:0005488,6286 个基因)、催化活性(GO:0003824,3228 个基因)等。

图1 油菜成熟期种子上调表达基因GO 注释图
通过对成熟期种子表达下调的基因进行GO 注释(如图2 所示)可知,生物过程中富集到基因条目数量最多的依次为代谢过程(GO:0008152,6 262 个基因)、细胞过程(GO:0009987,5429 个基因)、单有机体过程(GO:0044699,4 589 个基因)等。细胞组件富集到基因条目数量最多的依次为细胞(GO:0005623,2 735 个基因)、细胞部分(GO:0044464,2 735 个基因)、膜(GO:0016020,2 294 个基因)等。分子功能富集到基因条目数量最多的依次为绑定(GO:0005488,8 925 个基因)、催化活性(GO:0003824,6 661 个基因)等。
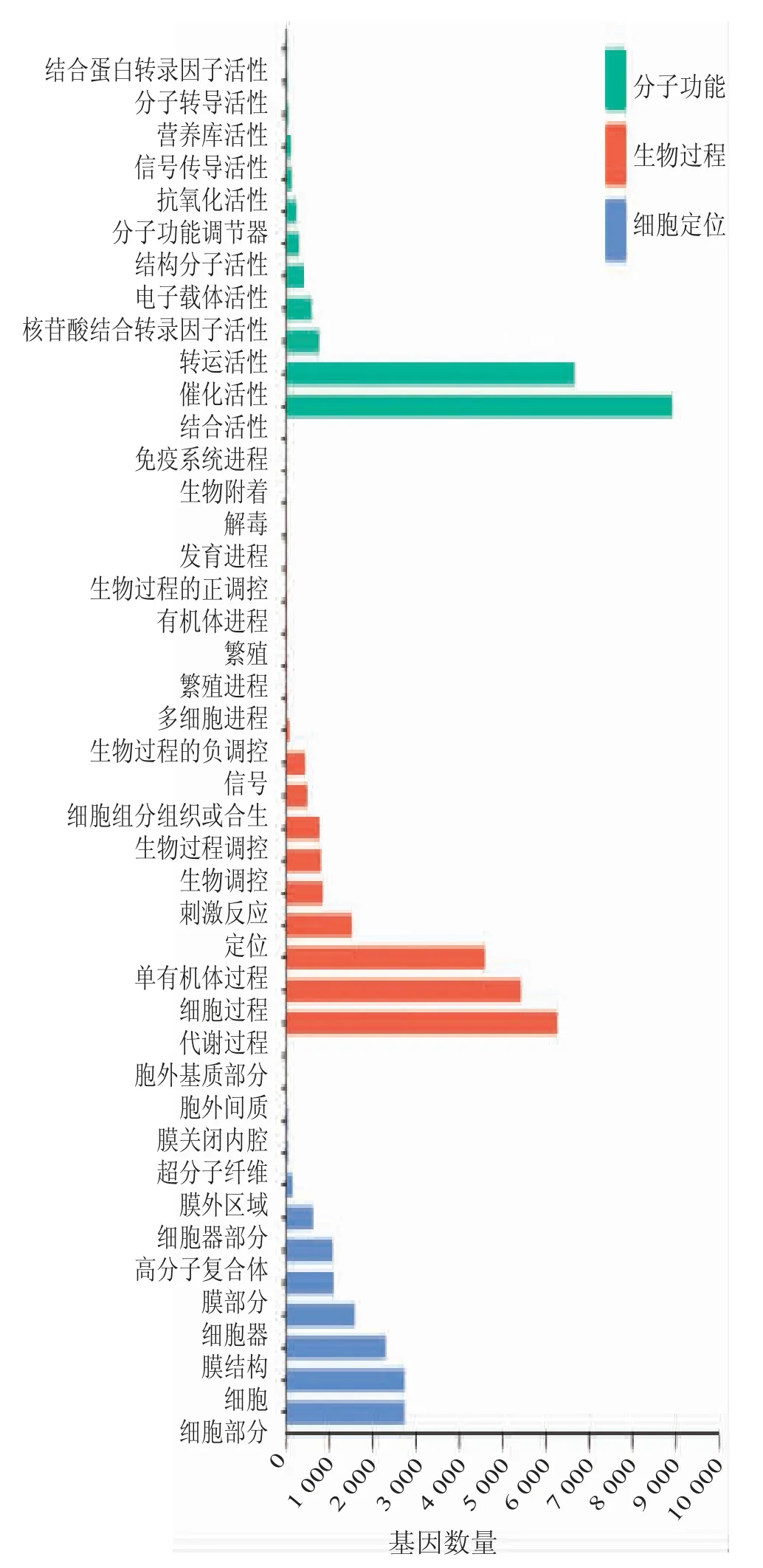
图2 油菜成熟期种子下调表达基因GO 注释图
差异表达基因GO 注释分析结果表明,成熟期的油菜种子和发育期的油菜种子中表达差异基因在生物过程中都富集于代谢过程,在细胞组件一类中富集于细胞、细胞部分、膜和细胞器,在分子功能一类中富集于绑定。乙酰辅酶A 羧化酶是脂肪酸合成过程中的关键酶,其主要存在于质体和胞质中,脂肪酸的合成是在叶绿体中完成。由此得以说明,油菜种子油脂合成和植物质体的代谢过程有关。
2.3 差异表达基因KEGG 分析
根据KEGG 数据库,对差异上调表达基因进行功能分类和Pathway 注释,如图3 所示,其中1 365 个基因注释到115 条代谢途径中,200 个基因富集到代谢通路途径,注释基因最多;156 个基因富集到核糖体途径;104 个基因富集到次生代谢物生物合成途径等。富集比例最高的是内酰胺类抗生素途径,富集比例为0.15,然后依次是0.14 的咖啡因代谢途径、0.125 的酮体合成和降解途径等。
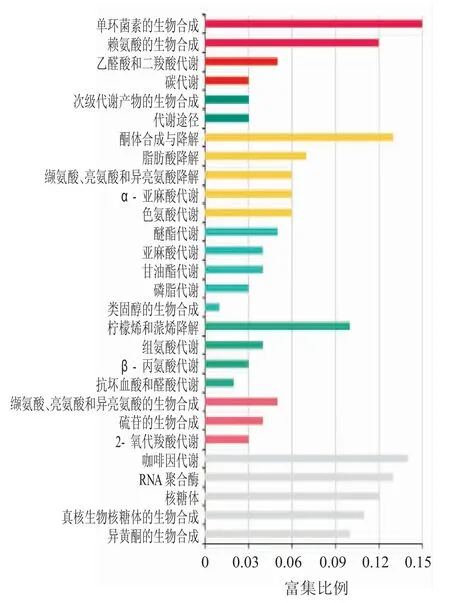
图3 油菜成熟期种子上调表达基因KEGG 富集分析
对差异下调表达基因进行功能分类和Pathway 注释,如图4 所示,其中1 653 个基因注释到115 条代谢途径中,337 个基因富集到代谢通路途径,注释基因最多;184 个基因富集到次生代谢物生物合成途径等。富集比例最高的是光合作用-天线蛋白通路途径和次级代谢物的生物合成通路途径,富集比例为0.295,然后是富集比例为0.237 的光合作用途径等。
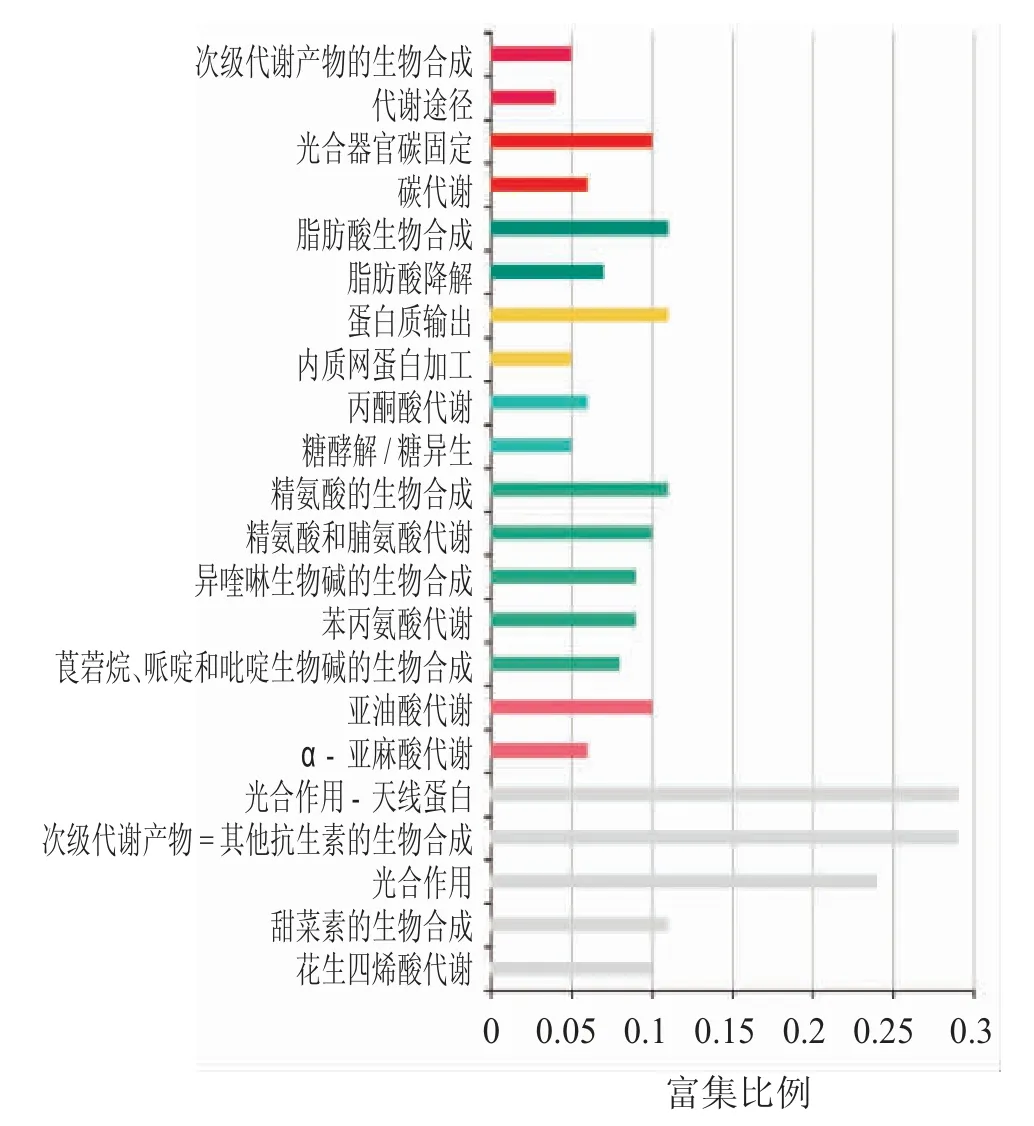
图4 油菜成熟期种子下调表达基因KEGG 富集分析
差异表达基因的通路分析表明,甘蓝型油菜成熟期种子中差异表达基因上调与代谢通路、核糖体途径等相关,上调基因富集于内酰胺类抗生素途径、咖啡因代谢途径、酮体合成和降解途径;差异基因下调则与代谢通路、次生代谢等相关,下调基因富集于光合作用-天线蛋白通路途径和次级代谢物的生物合成通路途径、光合作用途径。相关研究表明,作为脂肪酸合成首步反应的关键因子之一的ACCase,其活性受光限制,并且在油菜光合作用时,一些从叶绿体中产生的ATP和NADPH 会被用在脂肪酸的合成中。
2.4 油脂代谢相关基因动态表达分析
通过对差异表达基因的KEGG 富集分析发现,上调基因中参与脂肪酸降解、不饱和脂肪酸的生物合成、脂肪酸代谢、脂肪酸延伸的基因分别有11 个、4 个、7个和2 个,下调基因中参与脂肪酸生物合成、脂肪酸降解、不饱和脂肪酸的生物合成、脂肪酸代谢、脂肪酸延伸的基因分别有18 个、2 个、6 个、18 个和4 个,见表1。说明在油菜种子不同发育时期,参与脂肪酸生物合成、脂肪酸降解、不饱和脂肪酸的生物合成、脂肪酸代谢、脂肪酸延伸等与油脂代谢相关的基因表达水平发生了明显变化,共同调控油脂的合成与代谢。

表1 油菜种子油脂代谢相关差异表达基因汇总
3 结束语
油菜作为我国的主要油料作物,在我国农业生产和生活中占据着非常重要的地位。随着国民经济的飞速发展和市场的需求,提高油菜品种的油脂含量成为了育种者的主要目标之一。目前,国际上有许多关于油菜油脂合成相关基因的研究报道,但对这些基因如何调控油菜油脂合成等相关问题还有待科研工作者的后续研究。
本试验通过转录组测序技术,对甘蓝型油菜“中9”不同发育时期的种子样品的转录组数据进行分析,重点关注甘蓝型油菜种子不同发育时期与油脂合成有关的基因表达变化,为研究油脂合成相关基因代谢调控机理提供理论基础,同时也为后期选育高含油量油菜品种和RNA-seq 在分子生物学上的有效运用提供了支持。
猜你喜欢
今日农业(2021年19期)2022-01-12
今日农业(2021年20期)2022-01-12
房地产导刊(2020年9期)2020-10-28
疯狂英语·新读写(2020年3期)2020-06-06
西南农业学报(2019年10期)2019-11-14
环境与发展(2018年3期)2018-05-10
中国计算机报(2017年47期)2018-01-12
电影文学(2016年23期)2017-02-13
科学种养(2014年4期)2014-06-09
数理化学习·高一二版(2009年3期)2009-04-30
