
海水鱼循环水养殖系统中细菌群落结构研究进展
2022-08-05 06:22:10谷雪勤王庆奎王洋
天津农学院学报 2022年2期
谷雪勤,王庆奎,王洋
海水鱼循环水养殖系统中细菌群落结构研究进展
谷雪勤,王庆奎通信作者,王洋
(天津农学院 水产学院 天津市水产生态及养殖重点实验室,天津 300392)
循环水养殖系统(Recirculating aquaculture system,RAS)是一种环境友好且具有良好发展前景的养殖模式。细菌群落在RAS中具有重要作用,其数量和结构与RAS的水质和养殖动物的健康状况密切相关。本文总结了海水鱼RAS细菌群落结构的研究进展,比较了微生物载体、海水、鱼体表和消化道内细菌群落组成、多样性及功能,为在RAS中开展海水鱼健康养殖提供参考。
循环水养殖系统;海水鱼;细菌;群落结构
循环水养殖系统(Recirculating aquaculture system,RAS)是一种实现养殖用水循环利用的养殖模式。它将养殖体系中的水经过滤、曝气、沉淀、生物净化、脱气、紫外线消毒、充氧等处理,达到去除养殖水中的残饵、粪便和代谢产物,降低水中氨氮、亚硝酸盐等有害物浓度的目的,从而使水质达到养殖的标准[1-2]。与室外养殖方式相比,RAS具有养殖环境相对可控、养殖效率高、节约水资源、占地面积小等显著优点[3]。细菌是RAS中的主要微生物,主要分布在生物载体和养殖水体中。细菌作为RAS中主要的分解者,对水质和养殖动物的健康至关重要[4-5]。养殖动物消化道和体表的共生菌不仅对其健康有重要影响,也与RAS中的细菌有一定关联[6-8]。因此,研究RAS中的细菌群落结构及其与养殖动物健康状况的关系,对开展循环水健康养殖有重要指导意义。
1 海水鱼RAS中的细菌
海水鱼RAS中生物滤池、养殖水体以及鱼体的细菌构成的RAS细菌群落,对RAS的运行效率和养殖鱼的健康极其重要。我国采用RAS养殖的海水鱼主要有石斑鱼(spp.)、大菱鲆()、半滑舌鳎()和红鳍东方鲀()等[9]。微生物作为海水鱼RAS中的主要分解者,把粪便、残饵、代谢有机物降解为无机物,将氨氮、亚硝酸盐转变为硝酸盐或分子氮,从而达到净化水质的目的[10-11]。目前通常采用比表面积较大的固定或移动式微生物载体,如塑料纤维丝、生物滤料等提高海水鱼RAS中的微生物载量,从而提高微生物净化水质的效率[11-13]。在RAS运行初期,微生物载体上的细菌群落结构简单,净化水质的能力较差。随着细菌群落的演替,细菌群落结构趋于成熟,细菌净化水质的能力增强[14-15]。
细菌群落结构与海水鱼的健康和患病密切相关。海水鱼体表、消化道等存在大量细菌,既包含正常菌群,也包含益生菌和病原菌。很多水产益生菌是从养殖动物体表或肠道分离得到的[16-18]。海水鱼肠道是外源益生菌作用的主要位点,如芽孢杆菌、乳杆菌、链霉菌、肠球菌等常用做水产养殖中的益生菌[19]。外源益生菌可能对RAS中鱼肠道细菌群落产生影响。然而,高密度养殖条件下RAS中的鱼类免疫水平较低,易感染细菌性疾病[20]。广泛存在于海水鱼类中的爱德华氏菌病由迟钝爱德华氏菌()引起[21]。该菌引起牙鲆()和塞内加尔鳎()爆发疾病,每年造成养殖鱼类总死亡率达30%~40%[22-23]。由链球菌属(spp.)和乳球菌属(spp.)细菌引起的链球菌病则会导致鲽形目鱼类爆发疾病。研究表明,致病菌分布在鱼体、残饵、池壁污物及循环水各处理环节,苗种和养殖工具也可携带致病菌。池壁水位线处积累的污物是RAS中藏匿细菌最多的地方[24]。新进鱼苗的体内和体表、养殖管理人员、工具、空气等均可携带病原菌,引起RAS中海水鱼患病[25]。
2 海水鱼RAS细菌群落结构
2.1 RAS中生物滤池的细菌群落结构
生物滤池中的优势菌通常包含变形菌门、拟杆菌门、变形菌门下的γ-变形菌纲和α-变形菌纲。养殖对象不同,RAS中的细菌组成也有一定差异。例如,大菱鲆和半滑舌鳎RAS生物滤池中优势菌分别为变形菌门和黄杆菌门,从中也分离鉴定出了亚硝化单胞菌属及黄杆菌属等[12]。斑石鲷()RAS生物滤池内优势菌为变形菌门、绿弯菌门、拟杆菌门[26]。褐点石斑鱼()和豹纹鳃棘鲈()RAS生物滤池内优势菌为变形菌门下的γ-变形菌纲和α-变形菌纲[13]。红鳍东方鲀()RAS生物滤池优势菌为放线菌门、拟杆菌门、变形菌门下的γ-变形菌纲和α-变形菌纲[27]。石斑鱼RAS 3个不同级别的滤池内优势菌均为变形菌门和拟杆菌门[11]。半滑舌鳎RAS生物滤池前、后段和污泥中共有的优势菌为变形菌门和拟杆菌门,各自独有的优势菌分别为硝化螺旋菌门、酸杆菌门和浮霉菌门[28]。半滑舌鳎RAS生物滤池中的细菌主要由拟杆菌门的黄杆菌纲和变形菌门下的α-变形菌纲、β-变形菌纲、γ-变形菌纲组成,其中优势菌包括拟杆菌门下的冷蛇形菌属、奥雷氏菌属、湖饲养者菌属、沉积杆菌属、泥滩杆菌属以及变形菌门下的雷辛格氏菌属、亚硝化单胞菌属[29]。表1汇总了近七年国内外海水鱼RAS生物滤池的细菌群落结构。
表1 海水鱼RAS 生物滤池和水体中的细菌群落结构
地理位置养殖鱼种类环境因子取样部位细菌组成参考文献 山东省莱州市某养殖场斑石鲷水温、溶解氧、pH、总氨氮、亚硝态氮、硝态氮、化学耗氧量生物滤池中滤料优势菌为变形菌门、绿弯菌门、拟杆菌门[26] 浙江省温州市洞头县某养殖基地褐点石斑鱼、豹纹鳃棘鲈水温、盐度、pH、溶解氧、化学耗氧量、氨氮、亚硝酸盐、硝酸盐、磷酸盐等生物滤池变形菌门下的γ-变形菌纲和α-变形菌纲[13] 天津市滨海新区红鳍东方鲀氨氮、亚硝酸盐、硝酸盐等生物滤池放线菌门,拟杆菌门,变形菌门下的γ-变形菌纲和α-变形菌纲[27] 山东省莱州市某养殖场石斑鱼水温、溶解氧、pH、总氨氮、亚硝酸氮、硝酸氮、化学耗氧量生物滤池中的弹性滤料“毛刷”优势菌为变形菌门、拟杆菌门[11] 天津市滨海新区半滑舌鳎总氨氮、亚硝酸盐氮、盐度、pH生物滤池优势菌为变形菌门、拟杆菌门、硝化螺旋菌门、酸菌门、浮霉菌门[28]
续表
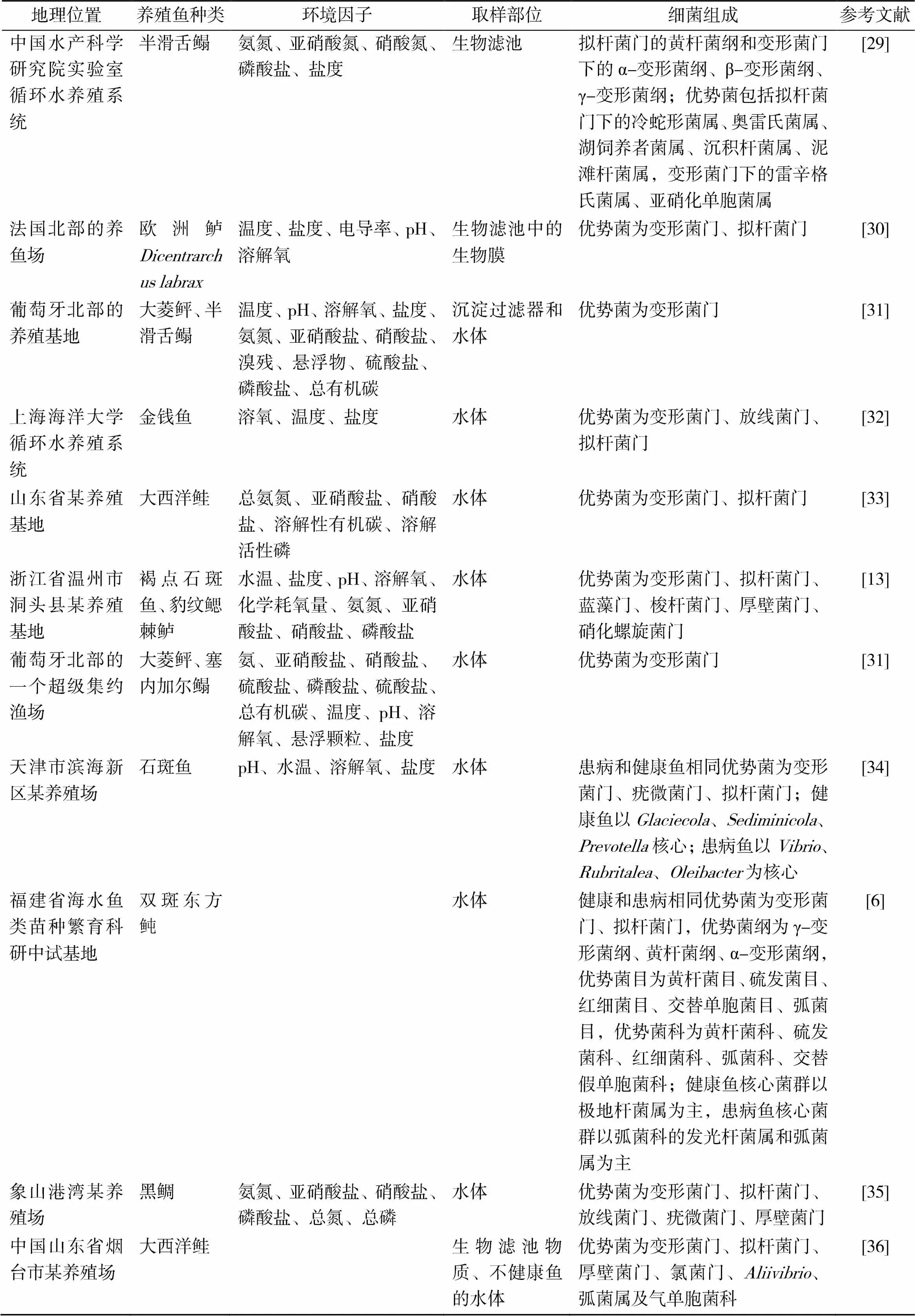
地理位置养殖鱼种类环境因子取样部位细菌组成参考文献 中国水产科学研究院实验室循环水养殖系统半滑舌鳎氨氮、亚硝酸氮、硝酸氮、磷酸盐、盐度生物滤池拟杆菌门的黄杆菌纲和变形菌门下的α-变形菌纲、β-变形菌纲、γ-变形菌纲;优势菌包括拟杆菌门下的冷蛇形菌属、奥雷氏菌属、湖饲养者菌属、沉积杆菌属、泥滩杆菌属,变形菌门下的雷辛格氏菌属、亚硝化单胞菌属[29] 法国北部的养鱼场欧洲鲈Dicentrarchus labrax温度、盐度、电导率、pH、溶解氧生物滤池中的生物膜优势菌为变形菌门、拟杆菌门[30] 葡萄牙北部的养殖基地大菱鲆、半滑舌鳎温度、pH、溶解氧、盐度、氨氮、亚硝酸盐、硝酸盐、溴残、悬浮物、硫酸盐、磷酸盐、总有机碳沉淀过滤器和水体优势菌为变形菌门[31] 上海海洋大学循环水养殖系统金钱鱼 溶氧、温度、盐度水体优势菌为变形菌门、放线菌门、拟杆菌门[32] 山东省某养殖基地大西洋鲑 总氨氮、亚硝酸盐、硝酸盐、溶解性有机碳、溶解活性磷水体优势菌为变形菌门、拟杆菌门[33] 浙江省温州市洞头县某养殖基地褐点石斑鱼、豹纹鳃棘鲈水温、盐度、pH、溶解氧、化学耗氧量、氨氮、亚硝酸盐、硝酸盐、磷酸盐水体 优势菌为变形菌门、拟杆菌门、蓝藻门、梭杆菌门、厚壁菌门、硝化螺旋菌门[13] 葡萄牙北部的一个超级集约渔场大菱鲆、塞内加尔鳎氨、亚硝酸盐、硝酸盐、硫酸盐、磷酸盐、硫酸盐、总有机碳、温度、pH、溶解氧、悬浮颗粒、盐度水体优势菌为变形菌门[31] 天津市滨海新区某养殖场石斑鱼pH、水温、溶解氧、盐度水体患病和健康鱼相同优势菌为变形菌门、疣微菌门、拟杆菌门;健康鱼以Glaciecola、Sediminicola、Prevotella核心;患病鱼以Vibrio、Rubritalea、Oleibacter为核心[34] 福建省海水鱼类苗种繁育科研中试基地双斑东方鲀 水体健康和患病相同优势菌为变形菌门、拟杆菌门,优势菌纲为γ-变形菌纲、黄杆菌纲、α-变形菌纲,优势菌目为黄杆菌目、硫发菌目、红细菌目、交替单胞菌目、弧菌目,优势菌科为黄杆菌科、硫发菌科、红细菌科、弧菌科、交替假单胞菌科;健康鱼核心菌群以极地杆菌属为主,患病鱼核心菌群以弧菌科的发光杆菌属和弧菌属为主[6] 象山港湾某养殖场黑鲷氨氮、亚硝酸盐、硝酸盐、磷酸盐、总氮、总磷水体优势菌为变形菌门、拟杆菌门、放线菌门、疣微菌门、厚壁菌门[35] 中国山东省烟台市某养殖场大西洋鲑 生物滤池物质、不健康鱼的水体优势菌为变形菌门、拟杆菌门、厚壁菌门、氯菌门、Aliivibrio、弧菌属及气单胞菌科[36]
海水鱼RAS生物滤池的不同部位,细菌群落的丰富度和多样性有差异。生物滤池一般分为3级,养殖水依次进入1、2、3级滤池,则3级生物滤池的细菌多样性最高。例如,石斑鱼RAS的3级滤池细菌群落丰富度和多样性高于1级和2级生物滤池,1级和2级生物滤池内细菌群落结构相似度较高[11]。斑石鲷()RAS的3级生物滤池细菌群落丰富度和多样性均高于1、2级生物滤池,2级和3级生物滤池细菌群落组成相似性最高[26]。
海水鱼RAS生物滤池中的细菌群落作为主要分解者,参与氮、硫的循环,可将对鱼有害的氨氮、亚硝酸盐、硫化氢转化为无毒物质。这一过程包括硝化、反硝化作用,其中亚硝化球菌、硝化螺菌、硝化螺旋菌门、亚硝化单胞菌属和硝化螺旋菌属等起重要作用。上述菌对海水鱼的健康有重要影响。从大菱鲆和半滑舌鳎RAS生物滤池中分离鉴定出亚硝化单胞菌属及黄杆菌属等,表明生物滤池中同时存在硝化和脱氮途径[12];石斑鱼RAS滤池内起硝化作用的菌主要是亚硝化单胞菌和硝化螺菌[11]。斑石鲷RAS生物滤池内起硝化作用的菌有亚硝化单胞菌属、硝化螺旋菌属[26]。
2.2 RAS水体细菌群落结构
大量研究发现,变形菌门和拟杆菌门是海水鱼RAS养殖水体中的常见菌门。养殖时间、养殖对象及其健康状态不同,海水鱼RAS中的核心菌群也不同。如金钱鱼()在RAS养殖一周后水体中优势菌为变形菌门的海单胞菌属和弧菌属,养殖一月后水体优势菌为拟杆菌门的和[32]。大西洋鲑()RAS养殖水体中细菌群落的优势菌门为变形菌门和拟杆菌门[33]。褐点石斑鱼和豹纹鳃棘鲈()RAS水体中优势菌为变形菌门、拟杆菌门、蓝藻门、梭杆菌门、厚壁菌门和硝化螺旋菌门[13]。大菱鲆和塞内加尔鳎RAS水体优势菌为变形菌门和拟杆菌门[31]。患病和健康青石斑鱼()RAS水体的优势菌为变形菌门,疣微菌门和拟杆菌门;健康鱼RAS水体以、和等对养殖动物有益或无害的菌属为核心,而患病鱼RAS水体则以、和等病原菌或有害菌属为核心[34]。健康和患病双斑东方鲀()RAS水体相同优势菌为变形菌门、拟杆菌门,优势菌纲为γ-变形菌纲、黄杆菌纲、α-变形菌纲,优势菌目为黄杆菌目、硫发菌目、红细菌目、交替单胞菌目、弧菌目,优势菌科为黄杆菌科、硫发菌科、红细菌科、弧菌科、交替假单胞菌科;健康鱼RAS水体核心菌群以为主,患病鱼RAS水体核心菌群以弧菌科的发光杆菌属和弧菌属为主[6]。根据海水鱼RAS水体细菌群落结构,表1汇总了近七年国内外海水鱼RAS海水的细菌群落结构。
海水鱼RAS海水中,细菌群落的多样性、丰富度、均匀度随养殖时间和鱼健康状态的不同而不同。随着养殖时间延长,养殖水体细菌群落多样性降低,菌群的丰度存在差异。如金钱鱼在RAS养殖一周后水体细菌群落多样性明显高于一个月后的。随着养殖时间延长,水体细菌的数量和种类逐渐减少,且不同时期水体的细菌群落组成有明显差异[32]。随养殖时间延长,红鳍东方鲀()RAS水体中弧菌属数量增长迅速,呈现聚集现象,均匀度低,而红细菌科数量均匀度高[37]。健康和患病双斑东方鲀RAS水体菌群多样性指数无较大差异,在同一分类水平上菌群丰度占比差异显著;二者核心菌群也差异明显,健康鱼以属为主;患病鱼以弧菌科的发光杆菌属和弧菌属为主[6]。健康和患病青石斑鱼RAS养殖水体中,细菌的相对丰度差异显著,而多样性无差异;健康鱼养殖水体中γ-变形菌门显著高于α-变形菌门;与患病鱼水体相比,健康鱼水中拟杆菌门的相对丰度明显升高,疣微菌门相对丰度明显降低[34]。
关于海水鱼RAS水体中细菌功能的研究报道较少。现有报道表明,水体中的细菌除具有净化水质作用外,某些菌还能抑制病原菌,维护海水鱼健康[6,37]。研究发现,海水鱼RAS水体中的细菌如皮特不动杆菌、哈夫尼希瓦氏菌、创伤弧菌、副溶血弧菌、枯草芽孢杆菌和光合细菌都能有效降低水中有毒有害物质,改善水体环境[38-41]。点带石斑鱼()、大菱鲆、红鳍东方鲀及半滑舌鳎RAS中的水体中皮特不动杆菌、哈夫尼希瓦氏菌、创伤弧菌和副溶血弧菌具有较强的除磷能力,去除氨氮的效果为哈夫尼希瓦氏菌>副溶血弧菌>皮特不动杆菌>创伤弧菌,其中皮特不动杆菌、哈夫尼希瓦氏菌和创伤弧菌除总氮的效果为皮特不动杆菌>哈夫尼希瓦氏菌=创伤弧菌[38]。从海水中分离的光合细菌能降解氨氮和亚硝酸盐,吸收水体氨态氮、硫化氢等有害物质,有效净化水质[39-41]。某些细菌还能抑制鱼体病原菌。如从水体中分离的荧光假单胞菌对鳗弧菌有抑制作用[42]。将大西洋鲑幼鱼浸浴于荧光假单胞菌液中,可降低压力引起的疖疮病的死亡 率[43]。从养殖水体中分离出的枯草芽孢杆菌可有效降低水体氨氮和亚硝酸盐,黄杆菌属、新鞘脂菌属和大单胞菌属等都具备降解污染物的能力[44]。
2.3 RAS中鱼体细菌群落组成
2.3.1 鱼消化道细菌群落结构
海水鱼的食性、健康状态不同,其消化道细菌群落组成也有差异(表2)。海水鱼消化道菌门通常有厚壁菌门、变形菌门、拟杆菌门和放线菌门,菌属通常有发光杆菌属和弧菌属,它们对海水鱼的健康都有重要影响。研究发现,厚壁菌门通常在海洋草食性鱼类肠道中占主导地位,而变形菌门通常在海洋非草食性鱼肠道中占主导地位[45]。健康大西洋鲑()肠道中常见菌门有变形菌门、放线菌门、拟杆菌门和厚壁菌门,菌科有微球菌科、草酸杆菌科,菌属有鞘单胞菌属、链霉菌属、土生杆菌属、、伯克霍氏菌属和巴尔内莫单胞菌属;不健康鱼肠道菌门主要有变形菌门、拟杆菌门和厚壁菌门,菌属主要有和弧菌属[36]。大黄鱼()幼鱼消化道菌门主要有变形菌门、厚壁菌门和拟杆菌门,菌属主要有发光杆菌属、乳杆菌属和弧菌属[46]。患病半滑舌鳎肠道菌群中绿弯菌目高于抗病鱼[47]。另外,网箱养殖花鲈()的肠道优势菌门为厚壁菌门、变形菌门、拟杆菌门、梭杆菌门,优势属为发光杆菌属、梭菌属及鲸杆菌属[48]。
鱼肠道菌群多样性与鱼体发育阶段、食物组成、健康状态有关。与健康鱼相比,患病鱼肠道菌群多样性显著降低。大菱鲆仔稚鱼(5~36日龄)消化道中的细菌数量先升高后降低;17日龄前后,弧菌总量差异显著,数量先升高后稳定;17~26日龄仔稚鱼消化道可培养细菌数量显著高于其他日龄[49]。黑鲷()摄食含羽毛粉的饲料后,其肠道细菌丰富度、多样性显著降低,其中,纲水平的拟杆菌纲、梭菌纲、δ-变形菌纲,科水平的草酸杆菌科,属水平的青枯菌属的相对丰度显著降低[50]。健康卵形鲳鲹()肠道菌群多样性低于患病鱼;健康卵形鲳鲹肠道中存在大量鳆发光杆菌,而患病鱼肠道中却不存在;患病卵形鲳鲹肠道中螺旋体门和黑海弧菌相对丰度显著升高,厚壁菌门和拟杆菌门相对丰度显著降低[51]。患病半滑舌鳎肠道中菌群多样性显著降低[52]。
肠道菌群在肠道发育、消化吸收营养物质和免疫反应中发挥着重要作用。目前在淡水鱼方面的研究报道较多,而海水鱼方面鲜有报道。大量研究发现淡水鱼肠道中特基拉芽孢杆菌和维氏气单胞菌、气单胞菌属和黄杆菌属的菌、鲸杆菌属和拟杆菌属的菌具有益生菌作用,其对淡水鱼的健康有重要影响。分离自鲫()肠道的维氏气单胞菌、阿氏肠杆菌和特基拉芽孢杆菌对嗜水气单胞菌具有体外抑制作用,其中维氏气单胞菌和特基拉芽孢杆菌可提高鲫的消化、抗氧化能力和免疫力,还能增加鲫肠道有益菌丰度,降低有害菌丰度[53]。鲤()肠道中黄杆菌属中的气单胞菌和黄杆菌能降低鲤对嗜水气单胞菌的感染率[54]。斑点叉尾鮰()肠道中的优势益生菌鲸杆菌属可产生维生素B12和抗菌肽[55],促进营养吸收,提高鱼体抗病力;拟杆菌属帮助分解营养物质、提高肠道营养利用率及维持肠道微生态平衡[56]。研究表明,少量的希瓦氏菌属能显著降低斑马鱼肠道中性粒细胞反应,这表明希瓦氏菌属对宿主肠道健康有重要影响[57]。
2.3.2 鱼体表的细菌群落组成
海水鱼体表常见菌一般由变形菌门、拟杆菌门、放线菌门、厚壁菌门和弧菌属组成。体表菌群在鱼体耐受多变环境,抵御机会致病菌时发挥重要作用[58-60]。鱼体表菌群组成与鱼的健康水平、种类有关。体表溃疡的半滑舌鳎,其体表存在致病性的菌有哈维氏弧菌、溶藻弧菌及副溶血弧菌[61]。体表溃疡的大西洋鲑,伤口黏液和边缘菌群以弯曲杆菌科和黄杆菌门下的黄杆菌科为主。健康大西洋鲑体表黏液存在变形菌门下的弯曲杆菌科和弧菌科,以黄杆菌门下的黄杆菌科为主,还有Planococcaceae和丛毛单胞菌科[62]。旗鱼()和银元鱼()体表粘液优势菌门为变形菌门,且主要由变形菌门下的 γ-变形菌门组成[63]。黄尾鰤()体表存在变形菌门、拟杆菌门和厚壁菌门,其中变形菌门为优势菌[64]。伯氏肩孔南极鱼()和革首南极鱼()体表共有优势菌包括变形菌门、厚壁菌门、拟杆菌门、放线菌门、异常球菌-柄热菌门和软壁菌门等,其中变形菌门和厚壁菌门分别是革首南极鱼和伯氏肩孔南极鱼体表的第一大优势菌群。两种鱼体表特有菌群为异常球菌-栖热菌门和软壁菌门。两种鱼体表菌属有火山岩菌属、芽孢杆菌属、嗜冷杆菌属、青枯菌属、短波单胞菌属、假单胞菌属、嗜甲基菌属和厚壁菌门的厌氧芽孢杆菌属,它们体表的优势菌群为厚壁菌门的厌氧芽孢杆菌属[65]。根据RAS中海水鱼体表细菌群落结构,表2汇总了近六年国内外RAS中海水鱼体表的细菌群落结构。
表2 海水鱼RAS中鱼体细菌群落结构
地理位置养殖鱼种类环境因子取样部位细菌组成参考文献 中国山东省烟台某养殖场大西洋鲑 肠道内容物健康鱼肠道优势菌为变形菌门、放线菌门、拟杆菌门、厚壁菌门,微球菌科和草酸菌科,链霉菌属、土壤杆菌属,细杆菌,伯克霍尔德氏菌、巴尔内莫单胞菌。不健康鱼肠道菌门主要有变形菌门、拟杆菌门和厚壁菌门,菌属主要有Aliivibrio和弧菌属[36] 福建省某养殖场 大黄鱼幼鱼Larimichthys crocea 消化道菌门主要有变形菌门、厚壁菌门、拟杆菌门,菌属主要有发光杆菌属、乳杆菌属、弧菌属[46] 山东省烟台市开发区某养殖场黑鲷 肠道纲水平有拟杆菌纲、梭菌纲、δ-变形菌纲,科水平的草酸杆菌科,属水平的青枯菌属[50] 山东省日照市东鑫现代渔业技术研究所半滑舌鳎 水温、盐度、pH值、溶氧、氨氮肠道优势菌为变形杆菌门、拟杆菌门、酸杆菌门、厚壁菌门、绿弯菌门、芽单胞菌门、疣微菌门、变形杆菌纲、绿弯菌目、6 -酸杆菌目[47] 澳大利亚南部温带水域的水产养殖场黄尾鰤 体表优势菌为厚壁菌门、变形菌门、拟杆菌门[64] 葡萄牙南部福尔摩沙某养殖场欧洲鲈 温度、光照周期体表优势菌为变形菌门、拟杆菌门、黄杆菌科、红杆菌科、弧菌科、红藻属、假单胞菌属[66] 天津市滨海新区某养殖场半滑舌鳎 体表病灶和肠道优势菌为哈维氏弧菌、溶藻弧菌、副溶血性弧菌[61] 罗浮敦和奥斯陆峡湾大西洋鳕Gadus morhua水温、盐度肠道优势菌为无色杆菌属、微球菌属、棒状杆菌属、交替单胞菌属、弧菌属、Clostridiales、Brevinematales、弧菌目、拟杆菌目[67]
鱼体表细菌多样性与鱼的种类和健康水平有关。一般健康鱼体表菌群多样性高于患病鱼。体表的不同部位,菌群组成的丰度有差异,而多样性无差异。研究发现,健康大西洋鲑体表菌群多样性高于体表溃疡鱼,而溃疡部位菌群多样性高于溃疡边缘部位[62]。同一地区的不同珊瑚礁鱼类,其体表菌群相比,丰富度具有显著性差异的菌有变形菌门的γ-变形菌纲,尤其是弧菌科、Altermonodales、根瘤菌目和梭菌目[68]。淡水鱼不同部位菌群多样性研究中,健康尼罗罗非鱼()体表菌群中的绿弯菌门、厚壁菌门、放线菌门、芽单胞菌门和酸杆菌门的相对丰度高于鳃样品,其中绿弯菌门、放线菌门、芽单胞菌门和酸杆菌门差异显著,而异常球菌-栖热菌门、变形菌门和梭杆菌门相对丰度则低于鳃样品,其中变形菌门和梭杆菌门差异显著[58]。
鱼体表共生菌群可能对宿主的免疫系统具有增强和保护的作用。研究表明,鲶()体表粘液提取物对铜绿假单胞菌有较好的抑制效果,推测体表粘液提取物中可能含有抑制铜绿假单胞菌的共生菌[69]。健康尼罗罗非鱼体表存在优势菌群包括特吕珀菌属、硫杆菌属、弓形杆菌属、海单胞菌属和弧菌属,这些菌群可能是尼罗罗非鱼体表的共生菌。人工感染无乳链球菌后,存活的尼罗罗非鱼体表菌群与感染前无显著差异;而濒死鱼体表菌群多样性降低,其中弓形杆菌属、假交替单胞菌属、海单胞菌属、假单胞菌属和弧菌属显著降低,链球菌属成为优势菌[58]。
3 总结与展望
海水鱼RAS中生物滤池、养殖水体以及鱼体存在的细菌,构成了RAS的微生态系统。其中生物载体上常见的菌为变形菌门、拟杆菌门,特有功能菌有硝化螺旋菌门、亚硝化单胞菌属、硝化螺旋菌属;养殖水体中优势菌有变形菌门、拟杆菌门、蓝藻门、梭杆菌门、厚壁菌门、疣微菌门和硝化螺旋菌门;鱼体表粘液优势菌有厚壁菌门、变形菌门、拟杆菌门、放线菌门和疣微菌门;鱼肠道优势菌包括变形菌门、拟杆菌门、放线菌门、厚壁菌门、弧菌属、发光杆菌属、假单胞菌属和。各部分的细菌组成会受到养殖鱼类、取样池塘以及养殖阶段的影响,与RAS的运行效率以及养殖鱼类的健康密切相关。
目前,关于海水鱼RAS细菌群落研究较少,有如下问题亟待解决:(1)尚未开展RAS中机会致病菌的组成、数量与致病机理的关系研究,盐度、温度、养殖密度和pH等因素与RAS细菌群落结构的关系尚不明确。有必要在弄清上述科学问题的基础上,建立基于RAS细菌群落结构健康评估方法和病害预警、防控技术。(2)大量研究表明,RAS生物滤池、鱼体以及水中细菌会发生迁移且相互影响,但不同细菌的迁移能力及各部分细菌群落间的影响机制尚不明确。
[1] 黄秀玉,荣浩翔,高月淑,等. 循环水养殖系统中反硝化技术研究进展[J]. 水处理技术,2019,45(2):7-12.
[2] 丁建乐,鲍旭腾,梁澄. 欧洲循环水养殖系统研究进展[J]. 渔业现代化,2011,38(5):53-57.
[3] 杨超,孙建明,徐哲,等. 循环水高密度养殖珍珠龙胆石斑鱼效果研究[J]. 渔业现代化,2016,43(3):18-22.
[4] BOUTIN S,BEMATCHEZ L,AUDET C,et al. Network analysis highlights complex interactions between pathogen,host and commensal microbiota[J]. Plos One,2013,8(12):84772.
[5] ZHANG D,WANG X,XIONG J,et al. Bacterioplankton assemblages as biological indicators of shrimp health status[J]. Ecological Indicators,2014,38:218-224.
[6] 吴建绍,朱志煌,李雷斌,等. 双斑东方鲀循环水养殖水体细菌群落结构分析[J]. 渔业研究,2018,40 (4):9.
[7] LOKESH J,KIRON V. Transition from freshwater to seawater reshapes the skin-associated microbiota of Atlantic salmon[J]. Scientific Reports,2016,6(1):1-10.
[8] SUN F,WANG C,CHEN L,et al. The intestinal bacterial community of healthy and diseased animals and its association with the aquaculture environment[J]. Applied Microbiology and Biotechnology,2020,104(2):775-783.
[9] 刘宝良,雷霁霖,黄滨,等. 中国海水鱼类陆基工厂化养殖产业发展现状及展望[J]. 渔业现代化,2015,42(1):1-5.
[10] 韩娜,马振兴,张伟,等. 对生态系统中分解者的定义及其所包含生物的探讨[J]. 生物学教学,2018,43(10):64-65.
[11] 黄志涛,宋协法,李勋,等. 基于高通量测序的石斑鱼循环水养殖生物滤池微生物群落分析[J]. 农业工程学报,2016,32(S1):242-247.
[12] 张海耿,马绍赛,李秋芬,等. 循环水养殖系统(RAS)生物载体上微生物群落结构变化分析[J]. 环境科学,2011,32(1):231-239.
[13] 吴越,马建忠,郑伊诺,等. 石斑鱼循环水养殖系统微生物群落结构[J]. 中国水产科学,2017,24(5):1045-1054.
[14] JIANG W,TIAN X,LI L,et al. Temporal bacterial community succession during the start-up process of biofilters in a cold-freshwater recirculating aquaculture system[J]. Bioresource Technology,2019,287:121441.
[15] ROALKVAM I,DRØNEN K,DAHLE H,et al. A case study of biofilter activation and microbial nitrification in a marine recirculation aquaculture system for rearing Atlantic salmon(L.)[J]. Aquaculture Research,2021,52(1):94-104.
[16] CARNEVALI O,MARADONNA F,GIOACCHINI G. Integrated control of fish metabolism,wellbeing and reproduction:The role of probiotic[J]. Aquaculture,2017,472:144-155.
[17] LAZADO C C,CAIPANG C M A,ESTANTE E G. Prospects of host-associated microorganisms in fish and penaeids as probiotics with immunomodulatory functions[J]. Fish & Shellfish Immunology,2015,45(1):2-12.
[18] MURALL C L,ABBATE J L,PUELMA T M,et al. Invasions of host-associated microbiome networks[M]// Advances in ecological research. San Diego:Academic Press,2017.
[19] 孙冬岩,孙笑非,王文娟. 宿主相关益生菌在水产养殖业中的研究进展[J]. 饲料研究,2021,44(20):93-95.
[20] PULKKINEN K,SUOMALAINEN L R,READ A F,et al. Intensive fish farming and the evolution of pathogen virulence:the case of columnaris disease in Finland[J]. Proceedings of the Royal Society B:Biological Sciences,2010,277:593-600.
[21] HAN H J,KIM D H,LEE D C,et al. Pathogenicity ofto olive flounder,(Temminck & Schlegel)[J]. Journal of Fish Diseases,2006,29(10):601-609.
[22] CASTRO N,TORANZO A,DEVESA S,et al. First description ofin Senegalese sole,(Kaup)[J]. Journal of Fish Diseases,2012,35(1):79-82.
[23] LIU F,TANG X,SHENG X,et al.outer membrane protein C:an immunogenic protein induces highly protective effects in flounder()against Edwardsiellosis[J]. International Journal of Molecular Sciences,2016,17(7):1117.
[24] 陈君. 工厂化循环水养殖半滑舌鳎主要细菌性疾病及其控制[D]. 上海:上海海洋大学,2012.
[25] 曹宝鑫. 生物絮凝和循环水养殖吉富罗非鱼(GIFT):越冬、大规格苗种培育及投喂频率的研究[D]. 上海:上海海洋大学,2019.
[26] 张皓天,宋协法,刘兴震,等. 斑石鲷循环水养殖系统生物滤池微生物群落对水质变化的动态响应[J]. 环境工程学报,2020,14(8):2176-2185.
[27] 陈珠. 红鳍东方鲀循环水养殖系统生物滤池内可培养微生物的多样性研究[D]. 青岛:中国科学院研究生院(海洋研究所),2013.
[28] RUAN Y J,GUO X S,YE Z Y,et al. Bacterial community analysis of different sections of a biofilter in a full scale marine recirculating aquaculture system[J]. North American Journal of Aquaculture,2015,77(3):318-326.
[29] 李秋芬,傅雪军,张艳,等. 循环水养殖系统生物滤池细菌群落的 PCR-DGGE 分析[J]. 水产学报,2011,35(4):579-586.
[30] ROQUIGNY R,MOUGIN J,BRIS C,et al. Characterization of the marine aquaculture microbiome:A seasonal survey in a seabass farm[J]. Aquaculture,2021,531:735987.
[31] MARTINS P,CLEARY D F,PIRES A C,et al. Molecular analysis of bacterial communities and detection of potential pathogens in a recirculating aquaculture system forand[J]. Plos One,2013,8(11):80847.
[32] 王振毅,张俊彬. 金钱鱼循环水养殖系统水体细菌的多样性[J]. 湖南农业大学学报(自然科学版),2018,44(4):6.
[33] 魏发奕,韩萃,李丽,等. 海水循环水系统中环境因子对大西洋鲑体内土腥味物质含量的影响[J]. 中国海洋大学学报(自然科学版),2021,51(3):34-44.
[34] 侯婷婷,钟志平,刘缨,等. 青石斑鱼海水循环水养殖水体的细菌群落特征[J]. 微生物学报,2016,56(2):253-263.
[35] 李卢国,骆云慧,徐善良. 循环水养殖黑鲷 ()水体中循环率对水质因子和细菌群落多样性的影响[J]. 海洋与湖沼,2020,51(2):318-327.
[36] WANG C,SUN G X,LI S S,et al. Intestinal microbiota of healthy and unhealthy Atlantic salmonL. in a recirculating aquaculture system[J]. Journal of Oceanology & Limnology,2018,36(2):414-426.
[37] 肖坤,郝婧薇,王艺,等. 循环水养殖系统微滤机过滤对调节水体细菌群落结构的影响[J]. 渔业现代化,2021,272(3):67-73.
[38] 周胜杰,路斌,贾婷婷,等. 养殖不同海水鱼的同种循环水系统中除磷细菌组成及特性[J]. 江苏农业科学,2017,45(12):120-123.
[39] 王雨婷,周荣翔,李霁虹,等. 抗弧菌光合细菌的分离鉴定及对氨氮和亚硝态氮的降解特性[J]. 南方水产科学,2021,17(5):26-33.
[40] 董艳珍,亓东明. 一株光合细菌的净水能力分析[J]. 西昌学院学报(自然科学版),2013,27(3):13-15.
[41] YANG A,ZHANG G,MENG F,et al. Nitrogen metabolism in photosynthetic bacteria wastewater treatment:A novel nitrogen transformation pathway[J]. Bioresource Technology,2019,294:122162.
[42] GRAM L,MELCHIORSEN J,SPANGGAARD B,et al. Inhibition ofbyAH2,a possible probiotic treatment of fish[J]. Applied and Environmental Microbiology,1999,65(3):969-973.
[43] SMITH P,DAVEY S. Evidence for the competitive exclusion offrom fish with stress- inducible furunculosis by a fluorescent pseudomonad[J]. Journal of Fish Diseases,1993,6(5):521-524.
[44] 朱莉飞,李伟,周雨琪,等. 罗非鱼养殖水体水质参数与菌群多样性分析[J]. 华北农学报,2020,35(S1):416-423.
[45] EGERTON S,CULLOTY S,WHOOLEY J,et al. The gut microbiota of marine fish[J]. Front Microbiol,2018,9:873.
[46] 姜燕,徐永江,柳学周,等. 工厂化和网箱养殖大黄鱼幼鱼消化道微生物群结构与功能分析[J]. 饲料工业,2019,40(6):35-43.
[47] 汪林庆. 半滑舌鳎肠道微生物及C9基因在免疫反应中的应答特征[D]. 福州:福州大学,2017.
[48] 林能锋,潘滢,许斌福,等. 养殖花鲈()肠道菌群的多样性分析[J]. 水产养殖,2021,42(6):1-7.
[49] 史秀清,张正,王印庚,等. 大菱鲆()仔稚鱼发育期消化道可培养细菌的菌群特征分析[J]. 渔业科学进展,2015,36(4):73-82.
[50] 邓惠文,刘峰,李桂豪,等. 黑鲷()肠道细菌群落对饲料中添加不同羽毛粉的响应[J]. 海洋与湖沼,2019,50(6):1309-1317.
[51] 熊向英,王贤丰,彭银辉,等. 健康和患病卵形鲳鲹肠道菌群结构的差异[J]. 水产学报,2019,43(5):1317-1325.
[52] 张正,廖梅杰,李彬. 两种疾病发生对养殖半滑舌鳎肠道菌群结构的影响分析[J]. 水产学报,2014,38(9): 1565-1572.
[53] 孙龙龙. 鲫肠道潜在益生菌的筛选与功能研究[D]. 重庆:西南大学,2020.
[54] 迟骋. 鲤肠道优势菌对其免疫功能的影响[D]. 陕西:西北农林科技大学,2014.
[55] ROESELERS G,MITTGE E K,STEPHENS W Z,et al. Evidence for a core gut microbiota in the zebrafish[J]. The ISME Journal,2011,5(10):1595-1608.
[56] BÄCKHED F,DING H,WANG T,et al. The gut microbiota as an environmental factor that regulates fat storage[J]. Proceedings of the National Academy of Sciences,2004,101(44):15718-15723.
[57] ROLIG A S,PARTHASARATHY R,BURNS A R,et al. Individual members of the microbiota disproportionately modulate host innate immune responses[J]. Cell Host Microbe,2015,18(5):613-620.
[58] 王淼,卢迈新,衣萌萌,等. 尼罗罗非鱼表皮和鳃黏膜共生菌结构特征及其与鱼体健康状况相关关系[J]. 水产学报,2017,41(7):1148-1157.
[59] 姜红烨,黄艳,余新炳. 鱼的黏膜免疫研究进展[J]. 热带医学杂志,2015,15(8):1150-1153.
[60] HURST C J. The Rasputin effect:when commensals and symbionts become parasitic[M]. Berlin:Springer,2016.
[61] 李文悦,左志晗,张晶晶,等. 半滑舌鳎溃疡病病原菌的分离、鉴定及其致病性分析[J]. 水产学报,2020,44(4):672-680.
[62] KARLSEN C,OTTEM K F,BREVIK Ø J,et al. The environmental and host-associated bacterial microbiota of Arctic seawater-farmed Atlantic salmon with ulcerative disorders[J]. Journal of Fish Diseases,2017,40(11):1645-1663.
[63] SYLVAIN F É,CHEAIB B,LLEWELLYN M,et al. pH drop impacts differentially skin and gut microbiota of the Amazonian fish tambaqui()[J]. Scientific Reports,2016,6(1):1-10.
[64] LEGRAND T P,CATALANO S R,WOS-OXLEY M L,et al. The inner workings of the outer surface:skin and gill microbiota as indicators of changing gut health in yellowtail kingfish[J]. Frontiers in Microbiology,2018,8:2664.
[65] 赵文静,许强华. 伯氏肩孔南极鱼和革首南极鱼皮肤微生物多样性的研究[J]. 大连海洋大学学报,2019,34(6):792-797.
[66] ROSADO D,XAVIER R,SEVERINO R,et al. Effects of disease,antibiotic treatment and recovery trajectory on the microbiome of farmed seabass()[J]. Scientific Reports,2019,9(1):1-11.
[67] RIISER E S,HAVERKAMP T H A,VARADHARAJAN S,et al. Metagenomic shotgun analyses reveal complex patterns of intra-and interspecific variation in the intestinal microbiomes of codfishes[J]. Applied and Environmental Microbiology,2020,86(6):2788.
[68] CHIARELLO M,AUGUET J C,BETTAREL Y,et al. Skin microbiome of coral reef fish is highly variable and driven by host phylogeny and diet[J]. Microbiome,2018,6(1):1-14.
[69] 李婷婷,张慧芳,姜杨,等. 鲶鱼体表粘液粗提物对铜绿假单胞菌的抑菌机理初探[J]. 现代食品科技,2015,31(7):67-73.
Research progress on bacterial community structure in recirculating aquaculture system for marine fish
Gu Xueqin, Wang QingkuiCorresponding Author, Wang Yang
(Tianjin Key Laboratory of Aqua-Ecology and Aquaculture, College of Fisheries, Tianjin Agricultural University , Tianjin 300392, China)
Recirculating aquaculture system(RAS)is an environment-friendly and promising aquaculture model. Bacteria play multiple functions in RAS. Its quantity and community structure are closely related to water quality and fish health in RAS. This paper summarized the research progress on the bacterial community structure in RAS for marine fish and analyzed the constitute, diversity and function of the bacterial communities in microbial vectors, seawater, fish skin and digestive tract. This paper provides a reference for the healthy culture of marine fish in RAS.
recirculating aquaculture system; marine fish; bacteria; community structure
1008-5394(2022)02-0076-09
10.19640/j.cnki.jtau.2022.02.015
S917.1
A
2021-12-21
国家重点研发计划(2020YFD0900600);现代农业产业技术体系专项资金资助(CARS-47);天津市海水养殖产业技术体系创新团队(ITTMRS2021000);天津市高等学校“创新团队培养计划”现代水产生态健康养殖(TD13-5089);天津市水产生态及养殖重点实验室平台项目(无编号)
谷雪勤(1994—),女,硕士在读,主要从事海水鱼循环水健康方面的研究。E-mail:2215566502@qq.com。
王庆奎(1978—),男,教授,博士,主要从事海水鱼循坏水健康养殖方面的研究。E-mail:wqkmail@qq.com。
责任编辑:张爱婷
猜你喜欢
化工管理(2022年14期)2022-12-02 11:44:26
当代水产(2022年8期)2022-09-20 06:45:40
科学养鱼(2021年5期)2021-11-30 20:55:06
食品安全导刊(2021年20期)2021-08-30 06:39:56
供水技术(2020年6期)2020-03-17 08:18:20
当代水产(2019年11期)2019-12-23 09:03:56
当代水产(2019年5期)2019-07-25 07:50:56
当代水产(2017年3期)2017-02-02 22:14:26
广东海洋大学学报(2015年3期)2015-12-22 10:05:26
环境科技(2015年6期)2015-11-08 11:14:26
