
伴生微生物与松材线虫-媒介天牛互作关系的研究进展
2022-08-04 00:12:32封小慧张宾孙江华
中国森林病虫 2022年3期
封小慧,张宾,孙江华
(1. 中国科学院动物研究所农业虫害鼠害综合治理研究国家重点实验室,北京 100101;2. 中国科学院大学,北京 100049;3. 河北大学生命科学学院,生命科学与绿色发展研究院,河北 保定 071002)
松材线虫病(pine wilt disease,PWD)是全球森林生态系统的巨大威胁,对我国的生态系统造成了巨大经济损失[1]。 松材线虫Bursaphelenchus xylophilus为松材线虫病的主要致病因子,而墨天牛属Monochamus昆虫是松材线虫的主要传播媒介,二者在长期进化过程中形成了紧密的共生复合体。 松材线虫向新的寄主松树传播扩散必须依赖于媒介天牛的携带,而松材线虫危害的病木又为媒介天牛提供了适宜的产卵场所,帮助其繁殖,如此重复,导致了松材线虫病的传播蔓延与暴发成灾(图1)。 研究表明,化学信号在松材线虫-媒介天牛复合体的形成和维持中发挥着重要作用。 如松墨天牛Monochamus alternatus羽化时产生的C16 -C18 脂肪酸乙酯能够诱导松材线虫幼虫从扩散型3 龄(LⅢ)转型到扩散型4 龄(LⅣ)[2]。 而小分子信号物质蛔甙(ascarosides)能够协调松材线虫和松墨天牛的一致发育[3];松墨天牛呼吸产生的高浓度CO2能够驱动松材线虫溢出天牛气管,侵染健康松树[4]。然而,越来越多的证据表明,伴生微生物(如细菌和真菌)在松材线虫-媒介天牛复合体中同样至关重要。 近年来,随着微生物高通量测序技术的快速发展,伴生微生物与松材线虫-媒介天牛间互作关系成为松材线虫病研究的前沿热点。
1 松材线虫-媒介昆虫伴生微生物组成及其地理变异
1.1 伴生微生物组成
1.1.1 松材线虫伴生微生物
松材线虫伴生微生物种类繁多,王慧利 等[5]从不同疫区以及不同松树种类的病木中,分离到泛菌属Pantoea、假单胞菌属Pseudomonas和消化链球菌属Peptostreptococcus三大类,共58 个菌株。 松材线虫的虫体、病死树内、媒介天牛幼虫坑道和蛹室中也存在着大量的伴生真菌。 例如,赵桂华 等从松材线虫疫木中分离出小型大单孢Haplosporella minor、长喙壳Ceratocystisspp. 、柳生球壳孢Sphaeropsis salicicola、环带头孢Cephalosporium zonatum和木生长喙壳Ophiostoma lignicola等多个松材线虫的伴生真菌菌株[6]。 伦莹莹 等从媒介天牛坑道中分离培养出10 多种优势长喙壳类真菌[7]。 松材线虫与其所携带的微生物经历长期协同进化,形成了从互利共生到拮抗等多种互作关系[8],影响着松材线虫的侵染循环和传播扩散[9]。
1.1.2 媒介昆虫伴生微生物
媒介昆虫的伴生微生物同样丰富多样,其中,肠道微生物最为关键[10]。 媒介天牛肠道微生物的组成和多样性随着其生长发育而变化。 例如,松墨天牛幼虫不同龄期,肠道内主要细菌属各不相同。 欧文氏菌属Erwinia是1 龄和5 龄幼虫中最丰富的属,肠杆菌属Enterobacter在3 龄和4 龄幼虫中占主导地位,2 龄幼虫中的主要属为克雷伯氏菌属Klebsiella、乳球菌属Lactococcus和伯克霍尔德氏菌属Burkholderia[11]。 也有研究报道媒介天牛幼虫肠道中变形杆菌门Proteobacteria 和厚壁菌门Firmicutes是最具优势的两个类群,肠杆菌属Enterobacter是松墨天牛幼虫肠道中最丰富的属,而沙雷氏菌属Serratia在天牛蛹肠道中占主导地位[12]。 还有研究表明,媒介昆虫加洛墨天牛Monochamus galloprovincialis早期幼虫中肠道相关细菌的多样性高于后期幼虫,这可能与它们之间的饮食差异有关[13]。 变形杆菌属Proteus和厚壁菌门Firmicutes 是加洛墨天牛幼虫肠道中最主要的类群,肠杆菌科(γ-变形杆菌)中的属占主要部分,并且发生在所有取样的幼虫肠道中[13-14]。 研究昆虫微生物组表征不仅有助于更好地了解细菌在松材线虫病中的重要作用,而且有利于生物控制策略的制定[15]。
1.1.3 寄主植物伴生微生物
寄主植物伴生微生物种类繁多。 感病马尾松Pinus massoniana和黑松Pinus thunbergii疫木的伴生真菌超过了30 个属。 其中,长喙壳类真菌(ophiostomatoid fungi)和霉菌可能对线虫繁殖和分布至关重要。 长喙壳类真菌多成为松材线虫的伴生真菌,利于其生长繁殖,而霉菌多是线虫的生物因子。 研究表明,与健康的松树相比,受感染的松树中的细菌多样性显著增加。 根瘤菌属Rhizobium、伯克霍尔德氏菌属Burkholderia和分枝杆菌属Mycobacterium是土壤和受感染松树中的主要细菌类群;Gryllotalpicola和纤维单胞菌属Cellulomonas是受感染的松树中的主要属[16]。 抗性松树和健康松树之间微生物种类和丰富度有较大差别,研究寄主植物伴生微生物,有助于探索基于微生物的松材线虫控制的研究。
1.2 地理变异
伴生微生物虽然种类复杂,但其有一定的特异性和选择性[17],来源不同国家的松材线虫其体表携带伴生细菌种类可能不同,甚至差别巨大。 巨云为 等分离鉴定了来自中国和日本的松材线虫所携带的细菌发现:在日本,松材线虫优势伴生细菌为产气肠杆菌Enterobacter aerogenus、产吲哚金黄杆菌Chryseobacterium indologenes、松鼠葡萄球菌Staphylococcus sciuri和耳葡萄球菌S.auricularis,其多为一些革兰氏阴性菌;而在中国,多为假单胞杆菌属Pseudomonas和泛菌Pantoea[18]。 曾腓力 等的研究也表明,日本松材线虫的优势伴生细菌多以芽孢杆菌为主,美国则以泛菌为主,而中国以假单孢杆菌和泛菌为主[19]。 贲爱玲 等[20]从来源于美国、日本和中国的松材线虫虫株上分离并鉴定了其各自的优势菌,同样发现日本虫株优势菌为荧光假单胞菌或阴沟肠杆菌Enterobacter cloacae,美国的优势细菌为成团泛菌Pantoea agglomerans,中国的优势细菌为阴沟肠杆菌或泛菌属的Pantoea toletana。 这些研究都说明松材线虫表面虽然都携带多种细菌,但不同地区有不同的优势种。 由此看来,松材线虫的伴生细菌经过长期的演替,种类和组成确实发生了很大的地理变异。
2 伴生微生物对松材线虫-媒介天牛复合体的影响
线虫和媒介昆虫的互作是利用遗传和分子手段研究免疫和寄生过程的良好模型。 昆虫和线虫各自都具有大量的伴生微生物,研究发现,伴生微生物与松材线虫-媒介昆虫天牛复合入侵体的关系密切。一些伴生微生物(如Sporothrixsp.1 等)能促进松材线虫种群数量增加,影响松材线虫的生长发育、繁殖、侵染传播等,也能提高松材线虫侵染、扩散概率,而有的微生物(如Serratia)则可作为松材线虫的防控因子(图1)。
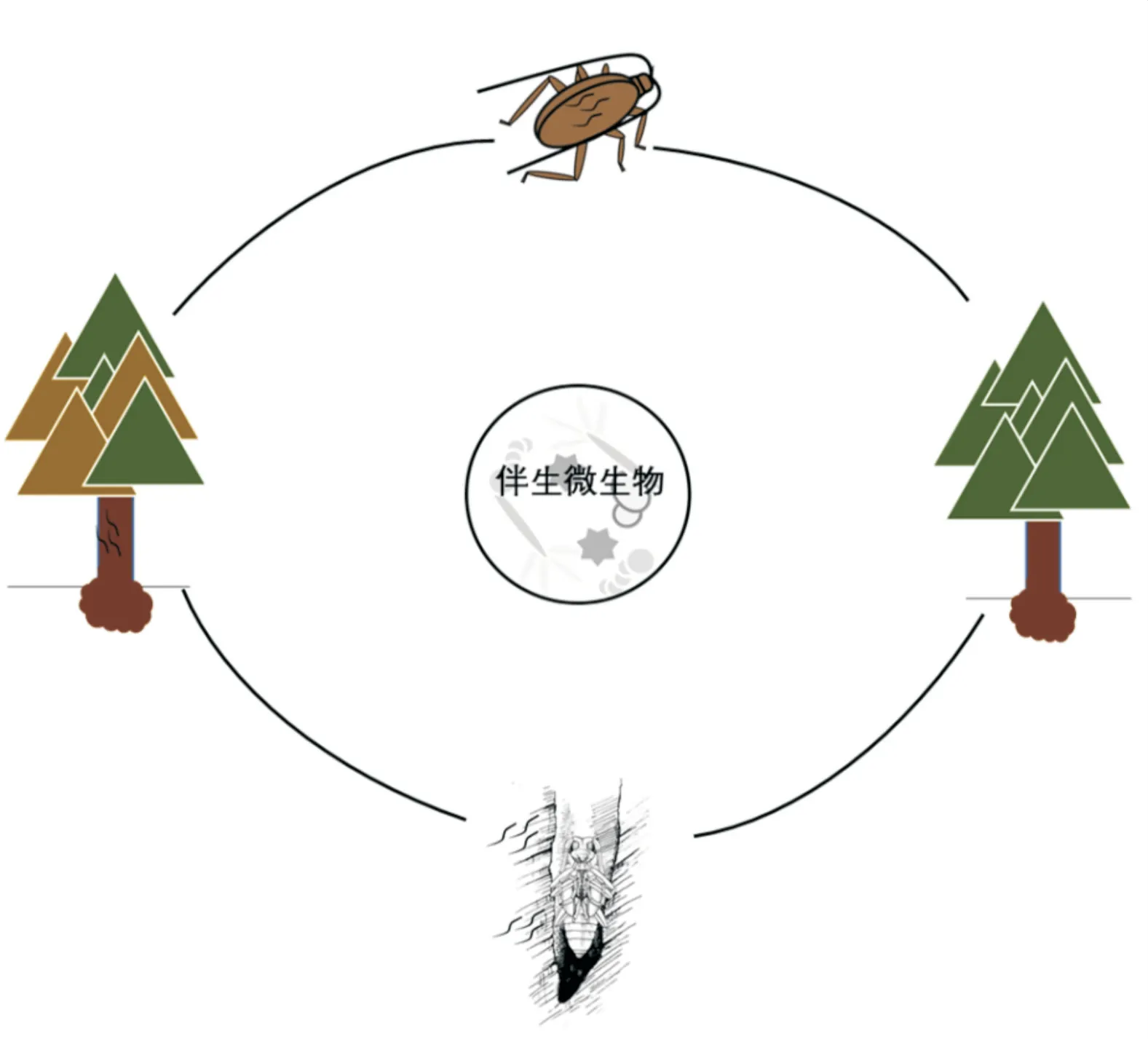
图1 伴生微生物与松材线虫-媒介天牛互作关系Fig.1 Interaction between associated microbes and PWN-Monochamus vector beetle complex
2.1 伴生微生物与松材线虫的生长繁殖
松材线虫与其携带的伴生微生物在长期协同进化后,形成了一种紧密的互利互惠共生关系。 赵博光 等[8]认为一些细菌附着在松材线虫表面,与松材线虫一同侵入寄主松树并为其生长发育以及繁殖提供一定的营养,形成对其有利的生存环境,同时提高了松材线虫的适应性和致病性。 松材线虫与其携带的伴生细菌存在某种营养共生,对双方的繁殖都有利,二者能互相提高适生性。 例如高浓度的坚强芽胞杆菌Bacillus firmus一定程度上可以增进松材线虫繁殖[21];恶臭假单胞菌Pseudomonas putida和荧光假单胞菌Pseudomonas fluorescens作为松材线虫的优势伴生细菌,它们的存在同样能够迅速提高松材线虫的繁殖速率,还可以促进松材线虫产卵[8,22]。多数伴生细菌能通过产生低浓度的苯乙酸来促进自身和松材线虫的繁殖,因此谈家金 等推测,苯乙酸在促进松材线虫繁殖中起重要作用[23]。 田雪亮 等[24]发现,伴生细菌能够通过改变某些代谢途径而降解松树体内的某些代谢产物,如通过降解对线虫有毒性的松萜类物质、苯甲酸类物质等,从而提高松材线虫的适生性,进而促进线虫的繁殖和种群数量的增加。 综上,伴生细菌不仅能为松材线虫降解某些毒性物质,而且能够分解某些营养物质,为松材线虫提供适宜的生存环境,从而促进其生长发育[24]。 在松材线虫的扩散期,松材线虫病木中的优势真菌,小孢拟盘多毛孢Pestalotiopsis microspora、松球壳孢菌Sphaeropsis sapinea、长喙壳菌Ceratocystisspp. 、木霉、曲霉、轮枝霉菌Verticilliumspp.等能显著增加媒介天牛携带松材线虫的数量;相同条件下,伴生优势真菌不同,松墨天牛羽化时所携带线虫的数量也不同[25-26]。 不同优势真菌对蛹室中线虫数量有显著影响,而松墨天牛携带的线虫数目又与蛹室中线虫数目呈正相关[2]。 例如,当松墨天牛蛹室的优势真菌为蓝变菌时,松墨天牛羽化时可以携带蓝变真菌(如长喙壳菌)孢子,这些真菌能增加天牛蛹室内松材线虫的种群数量从而提高松材线虫的入侵性[25]。 研究还发现,存在长喙壳菌时其松枝上的松材线虫虫体生长最快、种群数量最多[25]。赵莉蔺 等[2]的研究还发现,伴生真菌能同时影响入侵线虫和其媒介天牛的发育。 在中国本地特有真菌Sporothrixsp.1 存在的情况下,线虫能产生更多的雌性后代并且发育加快;Sporothrixsp.1 中活性物质双丙酮醇刺激松材线虫产卵和雌成虫体长增加而有效地促进松材线虫种群数量增长[2]。 也有学者提出,通过改变蛹室周围真菌群落种类组成可以改变松墨天牛羽化所携带的松材线虫数量[27-28]。
2.2 伴生微生物与松材线虫的致病性
许多致病杆菌能够存在于侵染型的线虫宿主肠道中,待宿主线虫侵染昆虫后,致病杆菌被释放进入昆虫血淋巴,进而大量复制最终致死昆虫[29]。 另外,一些致病杆菌部分基因的表达,有助于其宿主线虫的生长发育,提高线虫的繁殖力以及致病性[30]。有学者提出松材线虫病致病因素为松材线虫及其伴生细菌,二者缺一不可[31]。 不同线虫携带伴生细菌的种类存在较大差异。 池树友[22]从松材线虫虫体上分离到栖稻黄色单胞菌Flavimonas oryzihabitans和荧光假单胞菌,荧光假单胞菌是松材线虫携带的主要伴生细菌并且其毒性最强。 研究发现黑松接种无菌松材线虫9 个月后,仍有部分松树维持健康状态,而对照组接种带菌松材线虫的松树则完全发病[22]。 从健康株中只能分离到松材线虫,而分离不到细菌;但是从发病的松树上可分离到大量线虫和细菌,表明了松材线虫在细菌的存在下可能更容易致病[22],从而证明细菌影响松材线虫致病能力。 梁波 等[32]发现,当发生松材线虫病时,由于微生物的存在,黑松体内pH 值降低,并且存在着某种毒性物质可使黑松细胞死亡,证实松材线虫携带细菌所产生的致萎毒素对其致病性有促进作用。 覃德文 等[33]指出了松材线虫与其携带的细菌之间存在密切相关,毒素在松材线虫伴生细菌的致病机制中起着重要作用。 洪英娣 等[34]发现无菌松材线虫不能使黑松愈伤组织褐变和无菌黑松苗枯萎,而用无菌线虫与荧光假单胞菌Ⅰ型(Pseudomonas fluorescensbiotypeⅠ)、荧光假单胞菌Ⅱ型(P.fluorescensbiotypeⅡ)和泛菌属某种Pantoeasp. 分别混合接种均能使黑松无菌苗发生枯萎及愈伤组织严重褐变,证明伴生微生物能增强松材线虫的致病性。 叶建仁 等[35]的研究表明,携带种类和数量相似细菌的松材线虫其致病性却可能不同,而在不同地区、不同寄主体上致病力相同的松材线虫,它们所携带的细菌种类和数量也可能是不同的。 因此,关于松材线虫携带的细菌在松材线虫致病过程中如何影响或发挥致病作用的,还需要进一步研究证实[36]。
2.3 伴生微生物与松材线虫的入侵性
松材线虫伴生微生物在线虫侵染寄主和扩散过程中发挥重要作用,影响其入侵性。 研究表明,松材线虫经过细菌处理后,在松树体内的繁殖速度将得到提升。 袁为敏 等[37]通过电镜观察发现,松材线虫除了体表携带细菌,其体内也存在较多细菌,而且松材线虫体内环境较体表的复杂环境更为稳定。 松材线虫体内伴生细菌能帮助其适应松树体内的防御反应,提高其寄生能力,例如嗜麦芽窄食单胞杆菌Stenotrophomonas maltophiliaNSPmBx03 可以提高松材线虫的致病力和入侵性,提升染病马尾松枯萎速率[11]。 对于媒介天牛的共生微生物来讲,有些能够帮助宿主抑制线虫的侵染,增强自身免疫功能,具体表现在抑制寄生线虫在体内的生长,并且补偿由于线虫侵染而导致媒介昆虫损失的繁殖力[38]。 在松树枯死后松材线虫无法从松树获取足够营养时,伴生真菌就成为松材线虫唯一的食物来源,松材线虫取食伴生真菌,这样松材线虫种群在死亡松树上还能够繁殖扩散,增加了松材线虫的存活概率,同时也增加了松材线虫侵入新区的可能性[7]。 松材线虫的优势伴生真菌,不仅能显著促进扩散型松材线虫种群数量的增加,而且能显著提高松材线虫的扩散、传播能力[39-40]。 研究还发现,枯萎松树细菌和真菌的丰富度和多样性低于健康松树,表明嗜木性芽孢杆菌感染可能降低寄主植物内生微生物的丰富度和多样性,增强松材线虫的入侵性[41]。 尽管不同宿主昆虫之间优势细菌属的组成和丰富度存在很大差异,但来自不同宿主肠道的优势细菌属具有相似的代谢功能[42-43]。 已有研究表明:油松Pinus tabuliformis含有更高浓度和更多样化的次级代谢物,这使它成为云杉花墨天牛M.saltuarius的抗性树种;在取食油松的天牛幼虫肠道中,多样化的肠道微生物群有助于提高其代谢植物次生代谢物的能力,这有利于媒介天牛幼虫克服寄主抗性从而增强自身入侵性[44-45]。
2.4 伴生微生物影响线虫逃避天牛免疫
相对于繁殖型松材线虫、双胃线虫Diplogasteroides asiaticus、秀丽隐杆线虫Caenorhabditis elegans,扩散型4 龄松材线虫其体表携带更少的蛹室细菌。 研究明确了扩散型4 龄松材线虫几乎不会携带蛹室细菌进入松墨天牛气管,并因此实现对松墨天牛的保护,从而逃避天牛免疫,提高松材线虫-媒介天牛复合体的稳定性[46]。
电镜观察松材线虫发现,繁殖型松材线虫体表存在较多的分泌物,或许正是这层黏性物质能够使大量的伴生细菌牢牢地附着在松材线虫体表,增强伴生细菌的附着能力[47],而且这些伴生细菌能够从这些黏性物质中获得某些营养,从而形成牢固的共生关系。 而扩散型4 龄松材线虫表面的黏性分泌物较繁殖型各龄期松材线虫的黏性分泌物少,猜测正是这一特性使其表面粘附的伴生细菌携带量减少从而逃避天牛免疫,与媒介天牛形成很好的复合共生入侵体[47]。
实时荧光定量PCR 法分析并检测松墨天牛气管的细菌,结果表明松墨天牛其气管细菌总量在被松材线虫侵染后并没有显著变化[46]。 在大麦培养基添加蛹室细菌粘质沙雷氏菌Serratia marcescens后放入松墨天牛,在携带松材线虫的松墨天牛气管中并未检测到粘质沙雷氏菌,松墨天牛气管中的细菌数量也并没有因为携带松材线虫而显著变化。 此结果表明不携带伴生细菌维持松墨天牛气管的菌群稳定,确保了松墨天牛气管细菌菌群的相对稳态,有助于减少松墨天牛对松材线虫的免疫反应,提高复合体的共生性[46]。
3 松材线虫-媒介天牛对伴生微生物的影响
伴生微生物对松材线虫-媒介天牛复合体的形成和维持至关重要,而入侵复合体对其伴生微生物的结构和组成同样具有重要的影响。 不少研究表明,松材线虫-媒介天牛的复合共生体系也会反作用于其伴生微生物,对其伴生微生物的生长繁殖、种类分布产生影响。
3.1 松材线虫-媒介天牛对细菌繁殖的影响
松材线虫与伴生细菌之间存在某种营养关系,互相影响对方的生长发育,因此松材线虫的存在也能促进伴生细菌的生长发育,对其繁殖同样是有利的[48]。 研究表明,松材线虫不仅能促进其伴生细菌的增殖和产毒,其死尸还能给伴生细菌提供营养[49]。 郭道森 等[50]还证实了松材线虫的活体比死尸更加有利于洋葱伯克霍尔德氏菌Burkholderia cepacia菌株B619 的增殖,这表明不管是松材线虫活体还是死尸都可以促进其伴生细菌的繁殖。 松材线虫对其不同的伴生细菌有不同程度的促进作用,对荧光假单胞菌的促进作用最强,其次是泛菌,而对不解糖消化链球菌Peptostreptococcus asaccharolyticus却几乎没有作用[17]。 赵博光 等也发现松材线虫能明显增进荧光假单胞杆菌和恶臭假单胞杆菌的增殖[8]。
3.2 松材线虫-媒介天牛对寄主植物真菌群落的影响
松材线虫感染可降低寄主植物真菌的多样性。采用高通量测序结合功能预测(FUN Guild)对自然感染松材线虫松树的松针、根和土壤中及周围的真菌群落和功能结构进行探究[51],结果表明,病树针叶真菌丰富度、多样性和均匀度显著低于健康树,根系和土壤与健康树无差异。 主坐标分析(PCA)表明,真菌的群落和功能结构仅在病树和健康树木的松针中存在显著差异,在土壤和根部没有显著差异。这表明,PWN 感染主要影响黑松P.thunbergii针叶中的真菌群落和功能结构,但不影响根部和土壤[51]。 这些证明了受松材线虫感染后寄主植物真菌群落发生明显变化,并且在感染的早期微生物多样性的变化较为微小,随着感染症状的恶化,微生物变化的多样性显著增加。 也有研究发现,微生物群落多样性随病程进展而发生差异[52],进一步佐证了上面提到的寄主植物微生物群落因为受到松材线虫影响而发生变化。
4 应用与展望
松材线虫病在我国快速传播蔓延,且逐年加剧,迄今成为我国最为严重的林业外来有害生物之一[53],对其进行防控刻不容缓。 其伴生微生物群落的演变和进化可能是导致不同品系松材线虫存在致病性差异的主要原因之一[20]。 例如在美国,经历了长期共处的松树、媒介天牛、松材线虫和伴生细菌,四者发生了协同进化,伴生细菌可能发生了系列突变,从而进化到了一个较高级阶段,这些伴生细菌演变成了繁殖能力强和致病力低的优势菌种,这就导致了松材线虫病在其发源地美国却不是重要病害[19]。 因此,曾腓力 等[19]曾提出增殖培养美国松材线虫携带的这种弱致病力的优势细菌,来替代中国松材线虫携带的强致病力的伴生细菌,从而降低中国松材线虫致病力的想法,以达到生物防治的目的。 据专家分析,我国所有松树种类都有可能感染松材线虫病[54],但目前防控的主要问题为松材线虫病的整体防控策略相对比较单一,效果不太理想[55]。 因此,从破坏松材线虫-媒介天牛复合体的角度去阻遏其传播,将是非常理想的防控手段。 弄清松材线虫伴生微生物的种类及优势种,以及松材线虫对伴生微生物优势种的选择性、特异性和专一性,探究松材线虫与伴生真菌、细菌间的互作机制,筛选优势伴生微生物,弄清其致病机理,以便推广运用于松材线虫病的生物防治中。 例如,淡紫拟青霉Paecilomyces lilacinusPT1 菌株发酵滤液对松材线虫能起到强劲的毒杀作用,拥有理想的松材线虫病防治效果;白僵菌也是防治松材线虫的有效微生物之一,利用球孢白僵菌Beauveria bassiana对松墨天牛的强致病性,配合松褐天牛引诱剂一同使用,引诱剂发挥作用时,松墨天牛羽化聚集在引诱点周围的松树上取食时即会感染球孢白僵菌,并传染至其他天牛,从而实现其感染致死松墨天牛的作用[56]。 李亮亮 等[57]从南京湿地松Pinus elliottii上分离出蜡样芽孢杆菌Bacillus cereusNJSZ -13 菌株。 该菌株菌悬液处理线虫48 h 后线虫死亡率达到80%以上,并且该菌株在马尾松体内定殖能力强[58],对松材线虫病具有良好的生物防治潜能。
已有研究证明,遗传重组技术能够用于微生物的改造与菌株创新[59]。 因此,可以从伴生微生物遗传改造的角度进行松材线虫病防治。 挖掘伴生微生物的关键致病因子和通路,对伴生微生物进行基因改造,从而影响松材线虫和媒介天牛的互作。 例如,遗传改造松材线虫-媒介天牛复合体的主要伴生真菌Sporothrixsp.1,使其表达外源毒素,或通过代谢相关基因和通路的敲除从而积累毒性物质,可能是控制松材线虫和松墨天牛种群数量的有效途径之一[60]。 综上,深入了解松材线虫-媒介天牛与伴生微生物的互作关系,将为发展松材线虫病导向性防控技术提供重要的理论依据。
猜你喜欢
现代畜牧科技(2021年3期)2021-07-21 08:42:20
现代畜牧科技(2021年4期)2021-07-21 06:13:34
中国森林病虫(2020年6期)2020-11-28 07:50:32
亚热带植物科学(2020年4期)2020-09-28 03:08:32
绿色中国(2019年13期)2019-11-26 07:11:02
现代园艺(2017年21期)2018-01-03 06:42:16
现代园艺(2017年21期)2018-01-03 06:42:16
河北林业科技(2016年5期)2016-11-08 03:12:53
现代农业(2016年4期)2016-02-28 18:42:13
生物灾害科学(2015年2期)2015-03-11 16:38:31
