
中度火干扰后兴安落叶松群落物种组成及多样性变化1)
2022-08-04 01:48蓝俊杰刘艳红
东北林业大学学报 2022年7期
蓝俊杰 刘艳红
(北京林业大学,北京,100083)
林火兼具破坏性和生态性,林火不仅是影响森林群落维持和发展的活跃生态因子之一,也是森林生态系统结构和功能的重要组成部分[1-3]。林火干扰在一定程度上具有加快物种繁殖更新、促进植物生长、改良群落结构和丰富物种多样性等积极作用[4-6]。但更多的森林火灾直接烧毁大量植被,破坏森林现有的稳定结构,影响生态系统的稳定性,甚至干扰群落正常的演替方向[7]。大兴安岭林区是森林火灾的高发区,长期以来受到林火干扰,森林资源安全频繁受到威胁[8]。
物种组成与多样性是反映群落生态系统过程和功能的重要指标,是探究群落恢复和演替的重要内容[9-10]。研究表明,林火是森林更新和演替的主要影响因子[11],火烧迹地恢复初期,林下物种多样性为波动变化,整体表现为先升高后逐渐降低的趋势[12-14]。由于火烧在造成乔木树种大量伤亡的同时,改变了土壤中根蘖和种子的更新状态,因此火后乔木龄级和分布格局发生改变,直接影响次生演替格局及动态[15-17]。在大兴安岭地区,低强度林火可以促进白桦林向落叶松林演替[18];中重度的火烧,在恢复初期多以阳性或固氮植物为先锋物种占据优势地位[19],到了恢复后期,随着群落郁闭,群落物种数变化减小,林内环境稳定,生态功能得到恢复[20]。
目前,大多数关于火后森林群落恢复的研究多集中于不同林火强度的对比[21-24],少数以时间为尺度的研究也集中于恢复前期[25]。因此,本文以30 a恢复时间为研究尺度,以大兴安岭地区地带性植被类型兴安落叶松(Larixgmelinii)林为研究对象,通过对漠河地区中度林火强度下、不同恢复时间的火烧迹地进行研究,探究天然落叶松林火烧迹地恢复中的群落组成特征和植物多样性变化规律,为火烧迹地的植被恢复提供参考。
1 研究区概况
研究区位于黑龙江省大兴安岭地区漠河市(50°11′~53°33′N、121°12′~127°E),地处大兴安岭山脉北麓,以低山丘陵地形为主,一般地区较为平缓,海拔范围在200~1 400 m,平均海拔为600 m。漠河属于典型的寒温带大陆性气候,年平均气温为-4.2 ℃,年降水量350~500 mm,降水主要集中在每年的7—8月份。漠河典型植被类型为寒温带针叶林,主要建群树种为落叶松和樟子松(Pinussylvestris),常见伴生树种有辽东桤木(Alnussibirica)、白桦(Betulaplatyphylla)、山杨(Populusdavidiana)等。除地带性原生植被类型外,还有一些天然或人为形成的次生白桦林、山杨林、杂木林等。林下灌木和草本种类较为丰富。林下常见植物有:笃斯越桔(Vacciniumuliginosum)、越桔(V.vitis-idaea)、杜香(Ledumpalustre)等灌木;小叶章(Calamagrostisangustifolia)、齿叶风毛菊(Saussureaneoserrata)、细叶苔草(Carexrigescens)等草本。
2 研究方法
2.1 野外调查
2017年7月,在漠河市根据历史火灾记录确定了起火时间为1987年、1998年、2007年这3个年份的落叶松火烧迹地作为调查区。根据实地调查,在立地条件基本相似(即保持坡向、坡度、海拔相对接近)的火烧迹地上选取中度火烧区域(火后烧死木比例30%~70%)作为样地:2007年中度火烧迹地,恢复10 a(样地1);1998年中度火烧迹地,恢复19 a(样地2);1987年中度火烧迹地,恢复30 a(样地3)。在每类火烧迹地中随机设置3块20 m×20 m样地,并在未火烧的林地内设置3个对照样地,共12块样地,各类样地数据为3个重复样地的平均值。对每一块样地展开群落学调查,对乔木进行每木检尺,记录树高、胸径、冠幅等。在样地4角距边缘1 m处及中央各设1个5 m×5 m的灌木样方,并在每个灌木样方中沿对角线设置3个1 m×1 m的草本样方,调查灌木与草本的种类、株数、株高、盖度等指标。同时记录每个样地的海拔、地理坐标、坡向、坡度、土壤数据(土壤类型、土壤厚度、含水率等)、群落郁闭度等(见表1)。通过查阅以往研究资料,假定对照样地为恢复70 a的落叶松群落[26]。
2.2 数据处理
采用以空间代替时间的方法[27],通过对比未火烧群落,分析各火烧迹地物种组成与多样性特征随火烧后恢复时间变化的规律。运用EXCEL、SPSS等软件对数据分析处理;物种重要值分别对乔木层、灌木层、草本层进行计算;物种多样性选用的物种多样性测度指数(Simpson多样性指数、Shannon-Wiener多样性指数和Pielou均匀度指数)。

表1 样地基本概况
3 结果与分析
3.1 乔木层群落物种组成特征变化
由表2可以看出,与对照群落相比,火烧恢复初期落叶松重要值显著降低,且程下降趋势,但仍保持大于50%的比重;白桦、山杨、樟子松等伴生树种重要值由大到小的顺序为:对照、恢复10 a、恢复19 a、恢复30 a。在整个恢复过程中,乔木层密度变化较大,恢复10 a时,乔木层密度为最小值,只有对照群落的一半;恢复19 a时,乔木层密度达到最大值,明显高于对照群落;恢复30 a时,乔木层密度虽然接近对照群落,但树种组成已经差异明显。
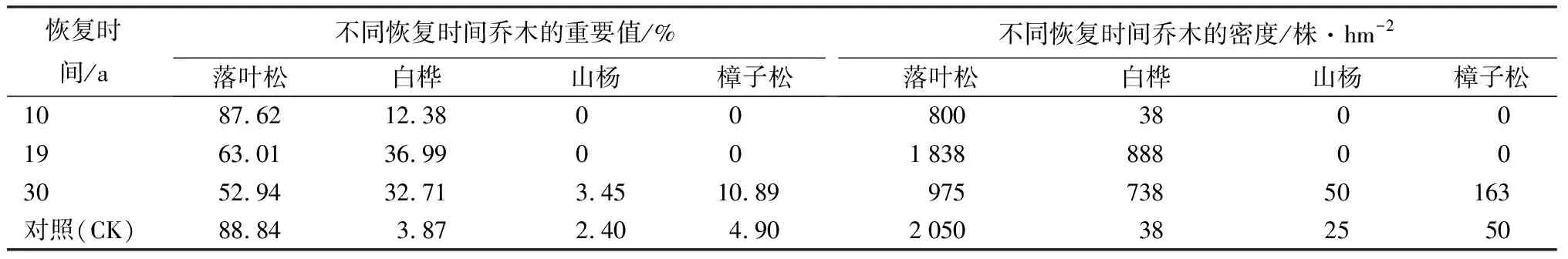
表2 中度火烧迹地不同恢复时间乔木重要值与密度
由表3可知,火干扰对恢复初期群落乔木层树种及优势树种落叶松的平均胸径、平均树高均存在显著影响,火烧后平均胸径、平均树高均较对照群落有所下降。其中乔木层的胸径和树高在恢复19 a时,均高于同一时期的落叶松;恢复30 a时,乔木层的胸径和树高已明显低于落叶松,且胸径显著低于其他恢复期和对照(P<0.05)。
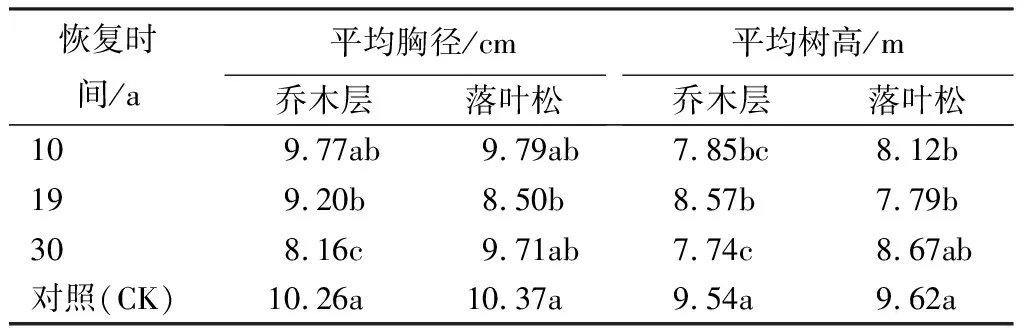
表3 火烧迹地不同恢复时间群落乔木层树种及优势树种落叶松的平均胸径和平均树高
3.2 林下群落物种组成及多样性特征变化
3.2.1 林下物种多样性变化
由表4可知,在中度火烧迹地恢复过程中,灌木层多样性指数随时间变化出现了两种情况,从Simpson指数和Shannon-Wiener指数来看,灌木层的多样性在恢复19 a时到达了较高水平,与其他3个恢复时期存在显著差异(P<0.05);而Pielou均匀度指数表现为各时期基本保持一致;灌木层多样性整体呈现由大到小的顺序为:恢复19 a、恢复10 a、恢复30 a、对照。草本层多样性指数随时间变化也出现了两种情况。在恢复10 a时,3种多样性指数均处于较低水平,之后的3个时期的Simpson指数和Pielou均匀度指数基本保持一致;而Shannon-Wiener指数表现为对照群落显著高于恢复19 a和恢复30 a时期,整体呈现出由小到大顺序为:恢复10 a、恢复19 a、恢复30 a、对照。
3.2.2 林下主要物种重要值变化
由图1可知,在林下物种恢复过程中,各类灌木和草本植物重要值均波动变化显著。灌木层在恢复10 a时,柴桦(Betulafruticosa)、笃斯越桔和杜香等灌木重要值处于较高水平,占据群落优势地位;在恢复19 a时,刺蔷薇(Rosaacicularis)和曲萼绣线菊(Spiraeaflexuosa)的重要值达到10%以上;而在恢复30 a和和对照群落内则是笃斯越桔、杜香和越桔等灌木重要值达到10%以上,且3种灌木重要值之和大于50%。

表4 中度火烧迹地不同恢复时间灌木层和草本层多样性指数
草本层在恢复10 a时,小叶章在群落中保持着绝对优势地位,重要值超过50%;在恢复19 a时,群落出现了山柳菊(Hieraciumkrameri)、秀丽马先蒿(Pedicularisvenusta)和酸浆(Physalialkekengi),3个物种的重要值之和近40%,齿叶风毛菊和细叶苔草重要值也处于较高水平;在恢复30 a时,齿叶风毛菊、地榆(Sanguisorbaofficinalis)、小叶章和细叶苔草占据优势地位,这与对照群落内的优势物种组成仍存在差异。
综合来看,越桔、笃斯越桔和杜香为该地区落叶松群落的优势灌木,齿叶风毛菊和小叶章为优势草本。
3.2.3 林下物种盖度变化
由图2可知,在中度火烧迹地恢复过程中,灌木层盖度变化与恢复时间呈显著正相关。而草本层盖度变化与恢复时间呈显著负相关。从整体来看,在恢复10 a时,群落灌木层盖度处于一个中等水平,随后逐渐上升,盖度最终趋于0.9左右;在恢复10 a时,群落草本层处于一个中等水平,随后逐渐下降,盖度最终趋于0.15左右。但在恢复19 a时,群落整体的灌木层盖度低于预期水平,这与草本层盖度在这一时期的变化恰好相反。由此得出,中度火烧迹地恢复过程中,灌木层盖度与草本层盖度变化随时间的增长显示出相反的趋势,但均在恢复19 a时存在异于预期水平的变化,总体呈互补的状态。
3.3 恢复时间与火烧迹地群落特征相关性
由表5可知,在所研究的中度火烧迹地上,恢复时间与乔木层平均树高、灌木盖度、草本盖度及部分灌木、草本多样性指数间呈显著相关关系(P<0.05);乔木层平均树高与平均胸径间存在极显著正相关关系(P<0.01),同时它们共同与灌木各多样性指数、灌木盖度、优势灌木重要值、草本盖度间呈显著相关关系(P<0.01);乔木密度与草本各多样性指数间呈极显著正相关关系(P<0.01)。
4 结论与讨论
林火干扰会影响群落乔木层树种组成,改变森林小气候和土壤养分环境,破坏森林原有的生态平衡,甚至改变森林演替方向[28-29]。在本研究中,群落乔木层受到林火干扰,恢复期内物种组成特征发生较为明显的变化。火烧后的群落在10 a恢复阶段,乔木密度仍处于较低水平,冠层受损较为严重。虽然此时落叶松占据着绝对的优势地位,但群落内产生的林隙、林窗已经为非优势树种生长提供了有利条件[30-31],一定程度上为改变群落物种组成格局奠定了基础。随着群落逐渐恢复,在19~30 a恢复阶段,白桦的密度和重要值显著增加,群落内还出现了樟子松、山杨等其他树种。这些伴生树种无论在密度上还是重要值上都体现了极强的竞争力。在恢复19 a和恢复30 a两个时期,乔木层的树高、胸径和落叶松的树高、胸径之间的大小关系发生了变化。主要原因,一是林木整体的竞争生长增加平均值,二是更新幼苗的大量生长稀释平均值。火烧程度较重将使白桦等阔叶树种的更新更为频繁,从而改变群落结构[32-34]的结论基本一致。因此,无论群落恢复过程如何变化,在本研究的时间尺度内,火后恢复的30 a时间里,落叶松虽然能够在乔木层保持重要值大于50%的优势地位,但实际上优势度已经受到了白桦、山杨、樟子松等重要值不断增长的伴生树种的影响,与对照群落相比,树种组成变化巨大,未来森林演替很可能朝着混交林的演替方向发展。

1.灌木10 a;2.草本10 a;3.灌木19 a;4.草本19 a;5.灌木30 a;6.草本30 a;7.灌木对照;8.草本对照。
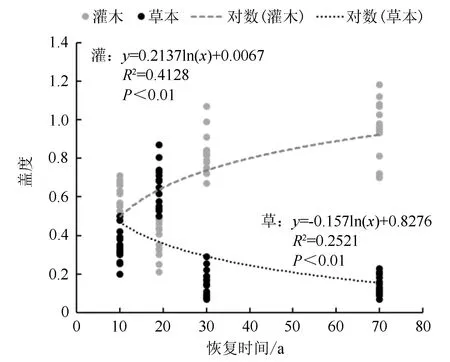
图2 中度火烧迹地不同恢复时间灌木层与草本层盖度变化
林下物种组成与多样性不仅是反映群落生态水平的重要指标,也与生态系统的稳定性密切相关[35]。火烧对于群落林下物种组成结构的影响显著,火后近几年内,群落内由于林冠稀疏,光照条件增强,使物种种类增加,多样性也随之增加[36-37]。因此,对于更长时间尺度的研究可以更好地揭示火干扰对群落的影响机制。火烧后,灌木层多样性整体呈现由大到小的顺序为:恢复19 a、恢复10 a、恢复30 a、对照,在恢复19 a时,多样性明显高于其它三个时期,并不意味着恢复19 a时群落就是稳定的[38]。乔木层平均胸径、平均树高与灌木层3种多样指数间存在显著负相关性(P<0.05),而与灌木层优势灌木重要值间存在显著正相关性(P<0.05),因此,灌木层的稳定与乔木层的物种组成特征间存在密切关系。特别是在恢复19 a时,乔木层密度明显提升的同时,灌木层多样性也明显提升,主要是因为灌木与小乔木之间竞争激烈。类似的研究表明,灌木层多样性在火后的10~22 a里波动较大,处于一个较为活跃的恢复期,群落内物种间竞争剧烈,物种数量、多样性与未火烧样地都有一定差距[20,39-41]。根据实地调查和相关研究发现,漠河处于冻土地带,受此影响越桔、笃斯越桔和杜香是常见的林下灌木,它们的株高低矮,能够在生态较为优良的林下环境成片覆盖地表,占据灌木层的主要生存空间[42]。当乔木层与灌木层间竞争加剧时,优势灌木重要值下降,而当林下环境趋于稳定后,优势灌木重要值上升。因此,可以将越桔、笃斯越桔和杜香等3种灌木作为对火后群落恢复水平具有指示性的参考物种。

表5 轻度火烧迹地恢复时间与群落特征的相关性
草本层多样性由大到小的顺序表现为:对照、恢复30 a、恢复19 a、恢复10 a。在恢复10 a时,多样性处于一个极低水平,之后升高至较为稳定的水平,这一结果以往研究发现有所不同[7]。由于在恢复10 a阶段,草本层小叶章的重要值过高,导致了极低多样性水平的出现。而随着上层乔木的更新恢复,群落冠层逐渐郁闭,有利于喜阴植物获得更多的生存资源,从而提升了多样性指数[19]。
林下物种盖度是直接反映群落恢复水平的指标,从灌木层和草本层的盖度曲线就可以直观地发现火后群落发生的显著变化。灌木层盖度随时间增加而增长,逐渐趋于稳定;草本层盖度随时间增加而降低,逐渐趋于稳定。与类似的研究比较,在整体趋势上草本层的表现基本一致,但灌木层的表现却完全相反,并未表现出逐渐降低的趋势[19]。主要原因是杜鹃-兴安落叶松群落中,兴安杜鹃通常植株较大[43],与乔木层间的竞争更为激烈,使得群落在逐渐郁闭的过程中林下光照条件改变,灌木生长受到影响。在恢复19 a时,灌木层和草本层的实际值都明显偏于拟合曲线的一侧。这一现象进一步表明了在恢复19 a时,前后群落内部种间资源和空间竞争的激烈状态。随着恢复时间增加,群落趋于稳定,最终达到一个以越桔、笃斯越桔和杜香等灌木为主的林下生态环境。综上,中度林火干扰对漠河地区落叶松群落物种组成及多样性变化影响显著。(1)乔木层树种组成变化巨大,虽然优势树种未变,但伴生树种的重要值持续上升。(2)林下灌木层和草本层受乔木层影响,出现种间竞争加剧的情况,盖度和多样性均产生相应变化。(3)整体来看,经过30 a的恢复时间群落林下灌木层和草本层已接近未火烧样地水平,但乔木层树种组成比例已产生显著差异。
可以推测,中度林火干扰对乔木层树种组成结构变化这种影响绝不是止步于恢复期前10 a对乔木层组成结构的破坏,更表现在之后的恢复期内对群落次生演替进程的影响。群落物种间竞争关系在恢复19 a前后明显加剧,物种组成与重要值明显波动。尽管之后群落趋于稳定,但在所研究的时间范围内,群落的演替已经与火烧前差异显著,原有的伴生树种比重大幅增加使群落向混交方向发展。因此,从保护生境及其生态系统的角度,对漠河地区火烧迹地进行适度人工辅助恢复,有助于兴安落叶松顶极群落的演替。
猜你喜欢
兰台内外(2021年33期)2021-12-17
青年文学家(2021年16期)2021-08-05
现代计算机(2021年7期)2021-05-12
少儿美术(2020年8期)2020-11-06
中国新闻周刊(2020年6期)2020-03-08
军民两用技术与产品(2018年11期)2018-07-16
河南科技(2018年16期)2018-07-13
绿色科技(2014年4期)2015-01-06
小说月刊(2014年1期)2014-04-23
