
干旱胁迫对中红杨光合生产和水分利用的影响1)
2022-08-04 01:47王怡霖安雄韬张芸香郭晋平
东北林业大学学报 2022年7期
王怡霖 安雄韬 张芸香 郭晋平
(太原工业学院,太原,030008)(山西省国有林场和种苗管理总站)(山西农业大学)
随着全球气候的改变,干旱频率、持续时间及严重程度不断增加[1],水分短缺已成为黄土高原干旱半干旱区限制植物生长发育与优良品种推广栽培的关键因素[2]。植物光合作用与种类、种源和品种等具有密切关系[3],对环境因子的响应也高度敏感,如轻度的干旱胁迫就会使植物的光合速率下降[4],因此光合作用是鉴定植物抗旱能力的重要指标[5]。植物水分利用效率(WUE)可以反映植物的耗水性[6],通常适度干旱可以提高植物水分利用效率[7],从而提高节水能力以有效降低对土壤水分的消耗[8]。植物水分利用效率也可用于评价水分利用与固碳能力的关系,植物水分利用效率越高,植物的耐旱生产力越强,越有利于维持生物量积累[9]。因此,以光合生产与水分利用策略为研究切入点,阐明杨树的抗旱机理对正确选择耐旱树种和科学制定苗期水分管理措施具有重要意义。
中红杨(Populus×euramericana‘Zhonghong’)为黑杨派杨树2025(P.deltoidescv. ‘Lux’(I-69/55)×P.deltoidscv. ‘Shan Hai Guan’)的芽变品种,其整株叶片从发芽至落叶由鲜红色至紫红色再至红绿色,最后变为杏红色,呈现“三季四变”的季相变化,是观赏性极佳的速生丰产型彩叶类绿化树种。目前,关于中红杨的抗旱研究多集中在生化指标[10-11]、光合生理[12-13]、色素含量[14],而对其干旱下光合生产的节水策略还未见报道。因此,本文选用黄土高原干旱半干旱区绿化树种中常见的毛白杨(Populustomentosa)与其进行比较研究,探讨中红杨和毛白杨光合生产与水分利用的抗旱策略,为黄土半干旱地区进行抗旱杨树品种的选育、适应性推广提供理论参考。
1 研究区概况
研究地位于山西省晋中市太谷县山西农业大学林学院试验基地(112°28′~113°1′E,37°12′~37°32′N),海拔767~900 m,年均气温9.8 ℃,年均降水量458 mm,供试土壤采自试验基地(20~40 cm),褐土壤,土壤有机质质量分数1.79 g/kg,全氮质量分数2.4 g/kg,全磷质量分数1.5 g/kg,土壤pH值7.22,最大田间持水量26.94%。
2 材料与方法
试验材料为1年生毛白杨(Populustomentosa)和中红杨(Populus×euramericana‘Zhonghong’)扦插苗,取自山西绿美园林绿化工程有限公司。
试验在基地的透光遮雨棚下进行,采用称质量法人工控制土壤含水量,设置3个水平的干旱梯度。1)正常供水CK:土壤相对含水量75%~80%(即盆栽土壤含水量占最大田间持水量的百分数;2)中度干旱MS:土壤相对含水量40%~45%;3)重度干旱SS:土壤相对含水量25%~30%。采取随机区组设计,每个品种每次处理17株,于2021年3月底扦插入试验用盆(高20 cm、直径30 cm),每盆1株,缓苗至6月30日开始干旱胁迫,分别于干旱胁迫第15、30、45天进行光合和生长指标的测定。
2.1 光合参数测定
每品种每处理分别选取生长势基本一致的6株幼苗,对植株中上部第3叶片标记。于上午10:00—11:30,采用Li-6400便携式光合仪(Li-6400XT,Li-COR,Lincoln,NE,USA)测定净光合速率(Pn)、蒸腾速率(Tr),计算水分利用效率(WUE=Pn/Tr)。测定时设定光合有效辐射强度(PAR)为1 000 μmol·m-2·s-1作为测定光强,并使用开放气路,流速为500 μmol·s-1,叶温26 ℃,相对湿度45%,外界CO2摩尔分数(Ca)为390 μmol·mol-1[15]。
使用光合仪的配件6400-02红蓝LED为光源,设定并记录PAR为1 800、1 600、1 400、1 200、1 000、800、600、400、300、200、150、100、60、40、20、0 μmol·m-2·s-1共16个光强点下的Pn。重复测定3次。采用非直角双曲线模型对不同干旱胁迫下净光合速率的光响应曲线参数进行拟合[16],并计算最大净光合速率(Amax)、暗呼吸速率(Rday)、光补偿点(LCP)和光饱和点(LSP)。
2.2 株高和地径测定
分别于干旱0、15、30、45 d测量株高(h)和地径(d),株高生长量=测定时株高-干旱0 d时株高,地径生长量=测定时地径-干旱0 d时地径。
2.3 数据处理与分析
采用单因素方差分析检验不同胁迫程度和干旱时长对各性状的影响。采用多重比较(P<0.05)检验不同处理间差异的显著性。采用双因素方差分析检验胁迫程度与干旱时长的交互影响。
3 结果与分析
3.1 干旱时长与胁迫程度对瞬时光合参数的影响
3.1.1干旱时长与胁迫程度对净光合速率(Pn)的影响
毛白杨和中红杨随干旱时长递增而下降,Pn由大到小表现为15、30、45 d;随胁迫程度的递增而下降,Pn由大到小表现为CK、MS、SS。干旱15 d下MS即引起毛白杨的Pn显著下降(P<0.05),干旱30 d才引起中红杨的Pn显著下降(P<0.05),干旱45 d毛白杨和中红杨的Pn在3个处理间均差异明显(P<0.05);相同干旱时长和胁迫程度下,毛白杨的Pn均比中红杨的Pn高;相较CK,干旱15、30和45 d,毛白杨Pn的降幅在MS下为15.14%、10.29%和36.62%,在SS下为15.45%、17.54%和56.74%,中红杨Pn的降幅在MS下为7.22%、19.66%和32.93%,在SS下为9.59%、26.72%和45.68%。两个品种对比,相同处理下毛白杨的Pn较高;相同胁迫程度下,中红杨的Pn在干旱30 d有较大降幅,在干旱15和45 d有较小降幅(表1)。
3.1.2干旱时长与胁迫程度对蒸腾速率(Tr)的影响
相同胁迫程度下,毛白杨随干旱时间Tr由大到小表现为30、15、45 d;中红杨随干旱时间Tr在MS下由大到小表现为30、15、45 d,在CK和SS下随干旱时间Tr由大到小表现为15、30、45 d。相同干旱时长下,毛白杨随胁迫程度Tr由大到小表现为CK、MS、SS;干旱15 d中红杨随胁迫程度Tr由大到小表现为CK、SS、MS,干旱30 d中红杨随胁迫程度Tr由大到小表现为MS、SS、CK,干旱45 d中红杨随胁迫程度Tr由大到小表现为CK、MS、SS。干旱15 d时,MS和SS相较正常供水,毛白杨的Tr降幅为10.52%和17.62%,中红杨的Tr降幅为5.77%、3.05%;干旱45 d时,MS和SS相较正常供水,毛白杨的Tr降幅为17.14%和24.70%,中红杨的Tr降幅为9.78%、12.15%;干旱30 d,MS和SS相较正常供水,毛白杨的Tr降幅为5.04%和10.65%,中红杨的Tr增幅为14.34%和3.08%。表明:干旱时长和胁迫程度可一定程度提高毛白杨和中红杨的Tr(表2)。

表1 不同干旱时长和胁迫程度时的净光合速率(Pn) μmol·m-2·s-1

表2 不同干旱时长和胁迫程度时的蒸腾速率(Tr) mmol·m-2·s-1
3.1.3干旱时长与胁迫程度对水分利用效率(WUE)的影响
相同时长下,毛白杨和中红杨的WUE由大到小表现为CK、MS、SS,除干旱15 d时SS提高了毛白杨的WUE。相同程度下,毛白杨和中红杨的WUE在MS下由大到小表现为15、45、30 d,在SS下由大到小表现为15、30、45 d。干旱45 d,毛白杨和中红杨的WUE在各胁迫程度间差异显著(P<0.05)。表明胁迫时长可一定程度提高毛白杨和中红杨的WUE(表3)。

表3 不同干旱时长和胁迫程度时的水分利用效率(WUE) μmol·mmol-1
3.2 干旱时长与胁迫程度对光响应曲线参数的影响
相同胁迫程度下,毛白杨和中红杨的Amax和Rday由大到小均表现为30、15、45 d,除SS下中红杨的Amax由大到小表现为15、30、45 d,Rday由大到小表现为30、45、15 d。干旱15和30 d,相较CK,毛白杨和中红杨的Amax和Rday在MS下变化均不显著,在SS下的变化均显著(P<0.05);干旱45 d,毛白杨和中红杨的Amax在各胁迫处理间差异均显著(P<0.05),毛白杨和中红杨的Rday在各胁迫处理间差异均不显著。表明:干旱30 d可提高毛白杨和中红杨的Amax和Rday;SS对毛白杨的Amax和中红杨的Rday影响大(表4~表6)。

表4 干旱15 d不同胁迫程度时的光合-光响应曲线参数 μmol·m-2·s-1
相同干旱时长下,毛白杨和中红杨的LCP和LSP总体均随胁迫程度的递增而下降,除LCP、LSP在MS下均有不同程度提高,表明MS能一定程度提高毛白杨和中红杨对弱光和强光的利用能力。同等处理下较中红杨,毛白杨LCP低、LSP高,表明毛白杨有效利用光照的范围宽(表4~表6)。
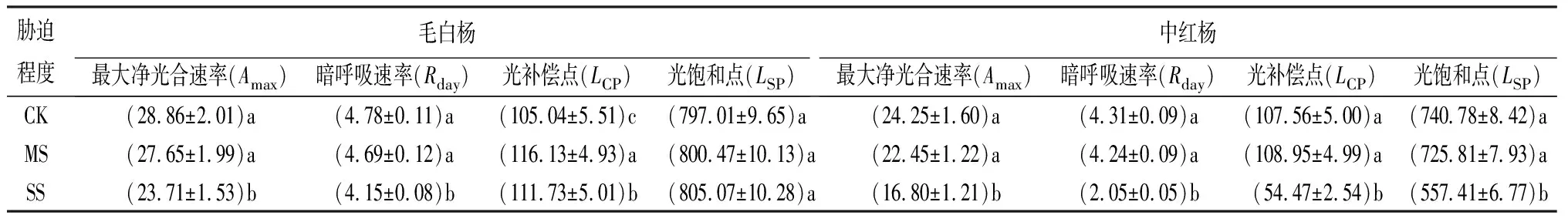
表5 干旱30 d不同胁迫程度时的光合-光响应曲线参数 μmol·m-2·s-1
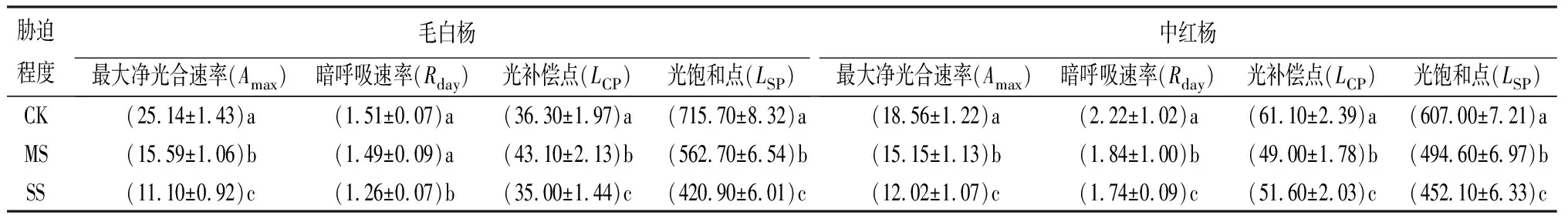
表6 干旱45 d不同胁迫程度时的光合-光响应曲线参数 μmol·m-2·s-1
3.3 干旱时长与胁迫程度对株高生长量的影响
干旱时间的延长和胁迫程度的递增引起毛白杨和中红杨的株高生长量降低,干旱前后差异显著(P<0.05)。相同胁迫时长和程度下,毛白杨比中红杨有更高的株高生长量且降幅更小,如干旱15、30、45 d,MS相较CK,毛白杨株高生长量降幅37%、50%、57%,中红杨株高生长量降幅61%、64%、64%;SS相较CK,毛白杨株高生长量降幅65%、71%、75%,中红杨株高生长量降幅70%、72%、73%。两个品种对比,干旱胁迫对中红杨的株高生长量影响大(表7)。

表7 不同干旱时长和胁迫程度时的株高生长量 cm
3.4 干旱时长与胁迫程度对光合生理和植株生长的交互影响
干旱时长和胁迫程度对毛白杨和中红杨的Pn、WUE、株高生长量交互影响均显著(P<0.05),对中红杨的Tr交互影响显著(P<0.05),对毛白杨Tr的交互影响不显著。两个品种对比,干旱时长与胁迫程度对毛白杨的Pn影响大;干旱时长对毛白杨的Tr影响大,胁迫程度对中红杨的Tr影响大;干旱时长较胁迫程度对杨树的WUE影响大;胁迫程度较干旱时长对杨树的株高生长量影响大(表8)。

表8 干旱时长与胁迫程度影响光合生理和植株生长的显著性水平
4 讨论与结论
植物对干旱胁迫的响应是一个非常复杂的过程,会因胁迫程度和胁迫时长呈现出明显的阶段性特点[17]。本研究中,连续干旱胁迫下两个杨树品种叶片光合与光响应参数变化规律相对一致,但各干旱时长、各胁迫程度下,两个杨树品种叶片光合能力受抑制程度存在明显差异,说明除干旱胁迫程度及胁迫时长外,品种也是影响杨树光合作用对水分亏缺反应的重要因素。可见,杨树的光合能力既受环境影响,更受遗传品种(基因型)制约,在相同的环境中,后者决定光合能力差异,这与张江涛等[13]的研究结果一致。光合作用是植株生长的生理基础,在不同干旱胁迫下的变幅可判断其抗旱性强弱[18],也可反映植株生长势[19]。本研究结果显示较中红杨,同等干旱时长和胁迫处理下,毛白杨具有高Pn和Amax,但两个指标均受干旱影响大而降幅大,说明毛白杨是一个具有高光合性能和干旱敏感型的品种,印证了干旱敏感型杨树比耐受型杨树有着更大的Pn降幅[20],这与其株高生长量高且降幅大也是一致的。
Rday在一定干旱胁迫范围内减小,说明叶片生理活性降低,使苗木呼吸减弱,可一定程度减少光合产物的消耗,有利于抵御干旱胁迫对苗木干物质积累的影响[21]。中红杨的Rday在SS下降幅大,说明其主要通过降低呼吸作用对光合产物的过多消耗,以适应干旱,从而有效缓解其株高生长量在SS下的降幅,这与李泽等[4]的研究结果一致。植物光合作用光饱和点和补偿点分别指示了植物叶片对强光和弱光的利用能力,代表了植物的需光特性和需光量[22]。干旱15、30、45 d时MS处理能一定程度提高毛白杨和中红杨LCP和LSP,但总体对弱光和强光的利用能力在干旱下减弱,这与桢楠幼树的表现相似[23],说明随着干旱胁迫程度和时长的递增,毛白杨和中红杨对强光和弱光表现出一定的可塑性和适应性,且毛白杨有效利用光照的范围较宽,对强光和弱光的利用效率都比中红杨高。
植物在水分亏缺时能以高效的水分利用方式来抵御干旱胁迫[8]。本研究中,相同干旱时长,随胁迫程度递增,毛白杨WUE有不同程度提高,这与前人研究基本一致[8]。中红杨持续下降,是由于8月份为杨树最旺盛的生长季,大气温度与平均日照辐射在全年相对最高,这些都会引起蒸腾速率的提高[24],而WUE是由Pn、Tr两个参数共同决定的[25],相同时间段其光合速率下降速度比蒸腾速率下降速度快会引起WUE的降低,反映了中红杨高蒸腾耗水和节水能力弱的特点。WUE能够反映植物耗水与其生产力之间的关系[26],毛白杨比中红杨有着更高的株高生长量和地径生长量,反映适度干旱下毛白杨既维持较高的光合作用水平和正常的生长发育,又防止了蒸腾作用引起的大量耗水,从而维持较高的水分利用效率,可以更有效的协调碳同化和水分消耗之间的关系,说明如果通过光合作用产生相同的干物质时,毛白杨消耗的水分较少,节水能力强于中红杨,这与南吉斌等[27]的研究结果相似。此外,相同干旱胁迫下两个杨树品种的WUE变化规律差异较大,很大程度上由基因遗传特性决定,支持了Masle et al.[28]的研究结论。
综上所述,两个杨树品种对比发现,干旱前后毛白杨光合速率和光合潜力高,有效利用光照的范围宽,对强光和弱光的利用效率高,这些均说明其光合性能显著强于中红杨,但干旱敏感度也显著高于中红杨,反映毛白杨通过降低Pn,牺牲碳固定,从而维持较高的WUE,采取了节水型抗旱策略,与幼龄柠条(Caraganakorshinskii)的表现相似[28]。反之,干旱胁迫下中红杨具有光合性能弱、蒸腾强且水分利用效率低的特点,导致其干物质生产能力相对弱,不适宜在干旱或缺水地区推广栽培。
猜你喜欢
灌溉排水学报(2022年9期)2022-09-28
农业工程学报(2022年8期)2022-08-08
作物学报(2022年10期)2022-07-21
江西农业(2022年8期)2022-05-13
绿色科技(2021年21期)2021-11-26
内蒙古林业调查设计(2021年3期)2021-07-06
安徽林业科技(2020年2期)2020-06-29
汽车与安全(2019年7期)2019-09-17
汽车与安全(2019年6期)2019-08-02
汽车与安全(2019年3期)2019-06-21
