
马尾松苗期抗旱性鉴定方法分析
2022-08-04 02:46:28尹明善蒙素讯桂云兰环江毛南族自治县华山林场广西环江54700环江毛南族自治县华山林业投资有限责任公司广西环江54700梧州市中森林业勘察设计有限公司广西环江54700
安徽农业科学 2022年14期
尹明善,蒙素讯,桂云兰 (.环江毛南族自治县华山林场,广西环江 54700;.环江毛南族自治县华山林业投资有限责任公司,广西环江 54700;.梧州市中森林业勘察设计有限公司,广西环江 54700)
水分是影响森林结构和物种分布的主导环境因子之一。在植物生长季节,严重水分亏损会抑制耐旱性差的植物生长发育,甚至造成严重不可逆的伤害导致衰亡,而耐旱能力强的物种在物种间的竞争中处于有利地位,从而得以大量繁殖。植物抗旱性研究一直是热点领域,国内外学者对植物抗旱性鉴定方法、评价体系及抗旱机制进行了大量研究,特别是植物应对干旱胁迫的响应机制。目前,植物抗旱性鉴定方法主要有田间鉴定法、干旱棚或人工模拟气候箱法、盆栽干旱法、大气干旱法、高渗溶液法等,以田间鉴定法、干旱棚和盆栽干旱法在作物抗旱选育中应用较普遍,在水稻、小麦、玉米、棉花等作物抗旱品种选育中应用较广。木本植物的抗旱性研究更倾向于干旱胁迫下苗木形态特征变换和生理生化响应观察,以及少数树种、种源、家系间抗性差异分析,缺乏适用马尾松(Lamb)抗旱性鉴定方法的探讨。抗旱品种的选育有利于根据不同环境特征提供相应的造林材料,因地制宜地发展林业产业。笔者通过对比不同抗旱性鉴定方法评价马尾松苗期抗旱性,筛选适用于松树苗期抗旱性鉴定的条件,为马尾松抗旱材料的批量筛选提供参考。
1 材料与方法
采用3个不同种源的半年生马尾松轻基质苗PM1、PM2、PM3作为试验材料。无纺布杯规格为高8.0 cm,直径3.5 cm,基质材料椰糠、黄心土、泥炭土按6∶3∶1配比。
干旱池基质选择。采用配对法设计,以过孔径 2.0 cm筛的新黄心土为土培干旱池基质,过孔径2.8 mm筛的河沙为沙培干旱池基质,基质厚0.2 m。选取生长一致的PM2苗木分别在土培基质和沙培基质内,按10.0 cm×10.0 cm的株行距定植40株,外围设置2行保护行,重复3次。待苗木正常生长14 d后开始干旱胁迫试验,胁迫前统一浇透水,采用L99-TWS-1土壤温湿度记录仪,每24 h记录干旱池基质湿度和苗木基质湿度1次,每次随机测量9个位点,同时观察记录植株萎蔫情况。根据试验结果选择最佳基质建立土壤湿度与含水量函数。土壤湿度用L99-TWS-1土壤温湿度记录仪测量,平行3次,同时测量位点土样土壤含水量。土壤含水量、饱和含水量参照张甘霖等的方法检测,用曲线估计对土壤湿度与含水量进行曲线拟合。
土壤干旱胁迫试验。
(1)极端干旱胁迫。采用随机区组试验设计,分别选取生长一致的PM1、PM2和PM3健康苗木,以30株为小区,按10.0 cm×10.0 cm株行距定植在干旱池内,外围设置2行保护行,重复4次。干旱池基质采用黄心土,厚度20 cm,待苗木正常生长14 d后统一浇水3 d,然后停水开始干旱胁迫,隔天记录1次植株萎蔫情况。萎蔫度的评价与计算参照徐超等的方法,当50%以上苗木达到永久萎蔫时进行复水,待苗木生长恢复稳定后统计成活率,评价苗木抗旱性。
(2)反复干旱胁迫。采用随机区组试验设计,设置轻度[土壤湿度为(32±3)%]、中度[土壤湿度为(24±3)%]、重度[土壤湿度为(16±3)%]和对照[土壤湿度为(40±3)%]共4个干旱胁迫处理,每处理选取生长一致的PM1、PM2和PM3健康苗木各30株,随机区组排列,重复3次,外围设置2行保护行。每天17:00观测一次土壤湿度,当土壤湿度达到处理下限时复水浇透。试验前测量苗高,当重度胁迫处理完成3个周期时停止试验,测量苗高和死亡率。
(3)梯度持续干旱胁迫。试验设计与“反复干旱胁迫”相同。当干旱池湿度达到各胁迫下限水平时,适当浇水,维持土壤湿度在干旱胁迫处理值上下限之间。试验前测量苗高,当重度胁迫达到30 d时停止试验,测量苗高和死亡率。
离体针叶失水率测定。每个家系选取生长一致的苗木10株,分别剪取苗木相同部位的针叶混合均匀,并立即称取5 g针叶测定鲜重(fresh weight,FW),重复3次,取平均值。将称重后的针叶置于垫有干燥滤纸的培养皿中,放置在温度为22 ℃、空气湿度为75%的环境下自然失水24 h,称得离体针叶失水重()。然后置于烘箱中,80 ℃烘至恒重,称量针叶干重(dry weight,DW)。失水率(water losing rate,WLR)按以下公式计算:
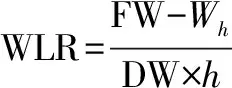
(3)
采用Excel 2010、SPSS 22、Origin Pro 9.1 进行试验数据统计分析及图表绘制。家系苗高生长量计算公式:苗高生长量=干旱胁迫结束后的苗高-干旱胁迫前的苗高。抗旱系数计算公式:抗旱系数=胁迫处理苗高生长量/对照处理苗高生长量。
2 结果与分析
干旱池基质水分均一性。从图1可见,充分浇透水后,土培和沙培干旱池基质最大湿度分别为43.6%、33.8% 。干旱池基质湿度均随干旱胁迫时间延长快速下降,第27天时,土培和沙培干旱池基质湿度均为最低,分别为6.0%、2.9%,随后趋于平衡。胁迫期间,土培干旱池基质湿度均明显高于沙培干旱池,湿度极差为37.6%,高出沙培(30.9%)6.7百分点。在持续干旱胁迫15 d前(土培和沙培干旱池基质湿度分别大于26.2%、13.8%),土培干旱池基质湿度变异系数小于沙培,但检验结果为差异不显著(Sig.=0.086);干旱胁迫15 d后,土培干旱池基质湿度变异系数大于河沙,检验结果为差异显著(Sig.=0.026);2种基质在整个干旱过程中湿度变异系数间差异显著(=0.151,Sig.=0.026),说明在整个干旱胁迫过程中2种基质不同位点湿度均一性差别不大。

图1 干旱池基质湿度(a)及其变异系数(b)变化趋势Fig.1 Change trend of humidity(a) and its coefficient of variation(b) in dry pool
干旱池内苗木轻基质湿度均一性。干旱池内苗木营养杯轻基质水分环境构成根系水分吸收小环境,而苗木根系周边水分环境差异往往是导致土壤干旱胁迫试验失败的主要因素之一。由表1可知,当干旱胁迫达到21 d时,土培干旱池黄心土基质和试验苗营养杯轻基质湿度分别为10.6%、9.6%,相差1.0百分点,变异系数相差0.51;沙培干旱池河沙和试验苗营养杯轻基质湿度分别为4.5%、20.6%,两者湿度、湿度标准差和变异系数的相差很大。当干旱胁迫达到30 d时,土培干旱池试验苗营养杯基质湿度为4.4%,较干旱池黄心土基质的湿度低2.2百分点,变异系数小0.04;沙培干旱池试验苗营养杯基质湿度为8.7%,变异系数为0.339,分别是河沙基质湿度和变异系数的3.0倍、2.7倍,是土培干旱池试验苗营养杯基质湿度的2.0倍、变异系数的3.8倍。可见,以黄心土作为干旱池基质,可以确保干旱池基质与苗木营养杯轻基质之间水分流动性,有效调节试验苗木间根系水分环境的一致性,而以河沙作为干旱池基质则难以达到该效果。
试验苗萎蔫度。马尾松苗木受到干旱胁迫后,会出现针叶失水萎蔫、植株脱水黄化、茎表皮微皱缩、植株顶端稍萎蔫下垂等不同程度的干旱伤害表型。土培干旱池内苗木在持续干旱18 d后开始出现萎蔫,而沙培干旱池内苗木在24 d出现萎蔫现象(图2)。萎蔫现象出现后,土培干旱池内苗木萎蔫度呈快速上升趋势,而沙培干旱池苗木萎蔫度则呈先缓慢上升再急速上升趋势,达到相同萎蔫度所需时间更长。该结果与干旱池基质和苗木营养杯基质湿度变化结论一致。可见,萎蔫度指标受苗木根系周围水分环境差异干扰较大,若采用萎蔫度作为抗旱评价指标,以黄心土作为干旱池基质有利于提高试验结果的可靠性。

表1 干旱池基质与营养杯轻基质湿度Table 1 Humidity of dry pool substrate and nutrition cup light substrate
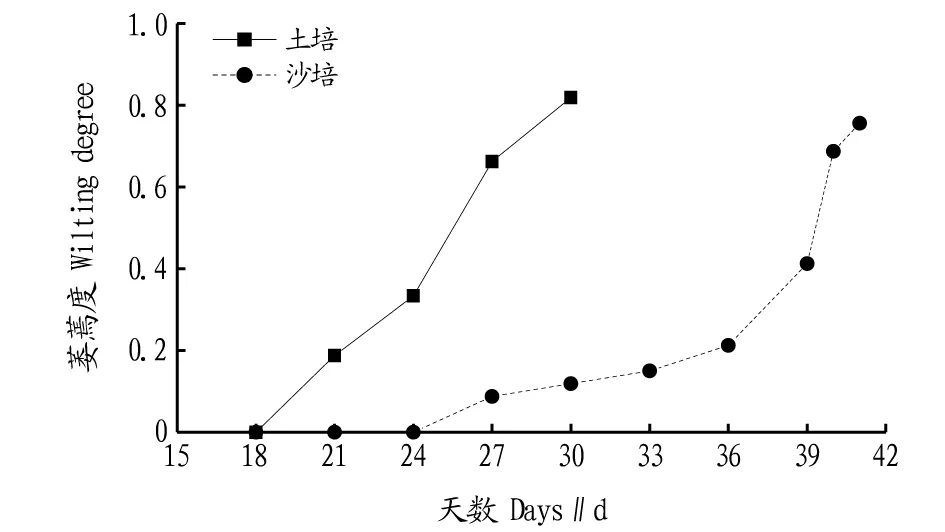
图2 不同干旱池基质苗木萎蔫度变化规律Fig.2 Variation of wilting degree of seedling in different dry pools
黄心土湿度与含水量曲线拟合。检测结果显示,试验用黄心土饱和含水量为38.0%。采用SPSS 22回归分析中曲线估计对黄心土湿度变化和含水量变化进行 11种曲线模型拟合,以三次曲线方程模型的相关指数最大,拟合曲线最佳。拟合方程如下:
=a+b+b+b
其中,为土壤含水量,为土壤湿度,拟合度=0988,回归方程显著性检验值=696526,显著性概率=0000<001,常数a=-6.375 925 616 344 82,参数b=1.894 351 901 411 99,b=-0.049 142 041 860 945 30,b=0.000 582 140 672 316 06。
极端干旱胁迫。PM1、PM2、PM3苗木在经过极端干旱胁迫复水后的枯死率分别为30.36%、12.50%、67.86%。单因素方程分析显示,3种苗木枯死率差异显著(=4.453,Sig.=0.045)。根据苗木枯死率评价,其抗旱性从大到小依次为PM2、PM1、PM3。
反复干旱胁迫。干旱胁迫程度不同,不同种源马尾松苗高生长量表现有所差异。由图3可知,随着土壤干旱胁迫的加重,PM1和PM3苗高生长量呈先增大后减小趋势,而PM2则先减小后增大趋势。在轻度和中度干旱胁迫下,PM1和PM3苗生长量均大于对照,而PM2苗高生长量低于对照,说明适度干旱有利于PM1和PM3苗高生长,充足的水分更有利于PM2苗高生长。在重度干旱胁迫下,各种源苗高生长量均小于对照,说明干旱胁迫程度超过某个临界值后,马尾松苗高生长均会受到抑制。抗旱系数是育种工作者评定品种表现较为通用的指标,抗旱系数越大,抗旱性越强。在轻、中度胁迫下,PM1、PM2的抗旱系数均小于PM3,在重度干旱胁迫条件下则相反。这表明在反复干旱胁迫下,不同胁迫条件获得的抗旱性评价结果不一致。在重度干旱胁迫下,3种苗木抗旱系数差异极显著(=35.435,Sig.=0.001),Duncan多重比较结果表明,除了PM1与PM2生长量差异不显著外,其余均差异显著。在重度干旱胁迫下,马尾松种源苗木的抗旱性强弱表现为PM2>PM1>PM3。

图3 反复干旱胁迫下马尾松苗高生长量(a)和抗旱系数(b)Fig.3 Seedling height growth (a) and drought resistance coefficient (b) of Pinus massoniana under repeatedly drought stress
梯度持续干旱胁迫。从图4可见,在不同程度的持续土壤干旱胁迫下,各种源马尾松苗高生长量均随着干旱胁迫程度的加重而下降,苗高生长量均低于对照,这说明持续控水不利于马尾松苗高的生长。通过对比反复干旱胁迫试验结果发现,在轻度、中度干旱胁迫下,复水方式不同,苗木抗旱性表现也有差异。在轻度、中度、重度干旱胁迫下,抗旱系数最大的种源苗木分别为PM1、PM3、PM2,即不同干旱胁迫梯度下各种源苗木耐旱性表现各不相同。在重度干旱胁迫下,各种源苗木耐旱性表现与极端干旱胁迫、反复干旱胁迫中的重度胁迫试验结果一致,且3种苗木抗旱系数差异显著(=5.456,Sig.=0.045),Duncan多重比较结果表明,除了PM1与PM2生长量差异不显著外,其余均差异显著。此外,在干旱胁迫截止期,PM3出现了11.90%的枯死率,而其他参试品种并未出现枯死现象,说明PM3的抗旱性表现最差。在重度干旱胁迫下,3种苗木的抗旱性强弱表现为PM2>PM1>PM3。

图4 持续干旱胁迫下马尾松苗高生长量(a)和抗旱系数(b)Fig.4 Seedling height growth (a) and drought resistance coefficient (b) of Pinus massoniana under continuous drought stress
通过检测各种源马尾松苗木离体针叶失水率发现,PM1、PM2、PM3离体针叶24 h的失水率分别为3.97%、3.47%、5.28%,单因素方差分析显示,差异极显著(=33.440,Sig.=0.001),Duncan多重比较结果表明,除了PM1与PM2离体针叶失水率差异不显著(Sig.=0.073)外,其余均差异极显著。该试验结果表明,PM2离体针叶保水能力最强,其次为PM1和PM3,各种源马尾松苗木离体针叶保水能力(即抗旱性)表现为PM2>PM1>PM3。
3 讨论与结论
干旱池或盆栽试验基质主要有土培、沙培和高渗溶液水培等材料。在作物抗旱品种选育中,以土培和沙培应用最为广泛。为了保存试验材料,该试验重点探索了采用营养杯苗作为试材的可靠性。试验结果表明,以黄心土作为干旱池基质,较河沙具有更高的饱和含水量,能有效调节试验苗营养杯基质湿度,确保试验苗木根系周围水分环境的均一性,提高试验结果的可靠性。
在作物抗旱品种选育中,反复干旱胁迫法应用较为普遍。2010年,姚艳荣等在玉米抗旱品种选育上进行了改进,提出采用一次干旱复水存活率鉴定玉米苗期抗旱性,可以更加实用和高效。离体叶片失水率则是国外最先应用于作物抗旱性鉴定的指标之一,该检测技术要求不高、设备简单,能够快速检测并反映植物叶组织保持水分的能力。通过比较不同试验评价结果发现,在反复干旱胁迫试验中,轻度和中度胁迫处理下,不同种源苗木抗旱性表现一致(轻度和中度胁迫下强抗旱性均为PM3,中抗旱性均为PM1,弱抗旱性均为PM2),而在重度胁迫处理下则出现差异(强、中、弱抗旱性表现较好的分别为PM2、PM1、PM3);在梯度持续干旱胁迫试验中,不同种源苗木抗旱性表现因胁迫处理而异;在所有试验中,极端干旱胁迫、离体针叶失水率苗木抗旱性表现与反复干旱胁迫试验、梯度持续干旱胁迫试验重度胁迫处理下的苗木抗旱性表现一致,即强、中、弱抗旱性表现较好的均为PM2、PM1、PM3。可见,在土壤干旱胁迫试验中,以重度胁迫处理下的评价结果为可靠。
在土壤干旱胁迫过程中,马尾松苗木开始出现萎蔫时,干旱池基质与苗木轻基质湿度分别为16.5% 和15.9%,苗木开始萎蔫的基质湿度临界值在15.9% 左右(拟合公式计算得土壤含水量为13.7%,相对含水量为36.0%),该相对含水量介于黑龙江省旱田土壤干旱指数分级标准中的严重干旱等级和永久凋萎点前期。可见,以黄心土作为干旱池基质开展马尾松苗期抗旱性评价,胁迫处理以土壤湿度可参照(16±3)%[土壤含水量为(14±2)%,土壤相对含水量为(36±6)%]。
猜你喜欢
吉林中医药(2021年12期)2021-03-27 17:55:48
农技服务(2020年1期)2020-12-17 08:15:01
山东农业科学(2019年11期)2019-12-24 01:11:27
现代农业科技(2019年1期)2019-07-11 01:36:25
安徽农业科学(2019年5期)2019-03-18 13:12:10
现代农业科技(2017年24期)2018-01-22 21:35:06
浙江农业学报(2017年1期)2017-05-17 06:13:36
西南农业学报(2016年4期)2016-05-17 05:42:13
山西农经(2015年7期)2015-07-10 15:00:57
中国民间疗法(2015年7期)2015-01-24 21:09:47
