
澜沧江两条支流捕食线虫真菌水陆分布差异及关联性
2022-08-03 01:55朱娅佼杨晓燕
生态学报 2022年12期
张 欣,邓 巍,朱娅佼,李 娜,*,肖 文,3,4,杨晓燕,3
1 大理大学东喜玛拉雅研究院, 大理 671003 2 云南省高校洱海流域保护与可持续发展研究重点实验室, 大理 671003 3 大理大学三江并流区域生物多样性保护与利用云南省创新团队, 大理 671003 4 中国三江并流区域生物多样性协同创新中心, 大理 671003
阐释生态系统中生物多样性的分布格局和维持机制一直是生态学研究的核心问题,而微生物是地球上各生态系统物质循环的枢纽,是生物地球化学循环的关键类群[1—3],对微生物多样性分布格局的认识成为了完善整个生物多样性空间分布格局研究的关键。近年来,越来越多的研究证明了微生物多样性存在明显的地理分布差异[4—7],而造成这种差异的机制尚不清楚。为明晰微生物多样性地理分布差异的形成机制,有研究者提出,微生物多样性分布格局的研究可能应该基于流域开展[8]。
流域由水生生态系统和陆地生态系统共同组成[9],要阐释微生物在其中的空间分布格局就必须同时考虑水生生态系统和陆地生态系统的相互关系。然而,一直以来,陆地都被认为是微生物的大本营,许多研究仅基于陆地开展[10—11]。另一些研究关注到了水生生态系统中的微生物[12—14],但之前的研究也只对比过水生和陆生类群的差异[15—16],水生和陆生物种多样性被严格区分,两者之间的相互关系被忽略。因此,对微生物在水陆生态系统中相互关系的长期忽视是造成微生物多样性空间分布格局产生机制尚不明晰的原因之一。要厘清造成微生物多样性分布差异的问题,就必须把水生和陆生生态系统联系起来,同时,考虑到微生物超强的扩散能力和巨大的多样性,相关研究还需要在环境异质性强的区域,进行系统的水陆对称样点布设,结合纯培养和分子手段,开展定性和定量的测定。
捕食线虫真菌是一类生态功能独特、营养类型多样、分离纯化鉴定较为容易的真核微生物,现已知的物种仅106种[17—25],广泛分布于全球各种水陆生境中。按照捕食器官类型,捕食线虫真菌分为产黏性菌网的Arthrobotrys属,产黏球、黏性分枝和非收缩环的Dactylellina属,以及产收缩环的Drechslerella属[17]。鉴于其分布广泛、水陆兼有、分离纯化和鉴定又较为容易的特性[26—27],以捕食线虫真菌开展微生物水陆分布格局研究具有独特的优势,厘清其水陆关系对于明晰整个微生物类群的水陆分布模式有重要的借鉴意义。因此,本研究选择高环境异质性的澜沧江支流为研究区域,在沘江和黑惠江两支流范围内采集水流汇入点及其上下游水陆对称样点的土壤样品,结合传统形态学和分子生物学方法分离鉴定捕食线虫真菌,探究捕食线虫真菌物种多样性在澜沧江支流水、陆间的分布差异及相互关系,厘清流域生态系统中环境过滤作用对捕食线虫真菌扩散的影响,回答水陆间是否存在扩散限制及其在流域生态系统中的物种多样性格局形成作用问题。
1 研究方法
1.1 研究区域概况
澜沧江流域地势由北向南呈阶梯状下降,主体地貌特征表现为高山峡谷相间,随山脉南延,山川间距由上游向下逐渐展宽[28]。流域内支流众多,河流流量大,地势起伏,河流落差大[29],生物资源十分丰富[30]。本研究基于澜沧江的两条支流沘江和黑惠江开展,根据实际地形情况,在研究区域内每间隔50 km设置一个样点,规避城市等强人为干扰区、高山深谷等不可到达的区域,最终于沘江和黑惠江两条支流上设置12个采样点,具体样点布设如图1。
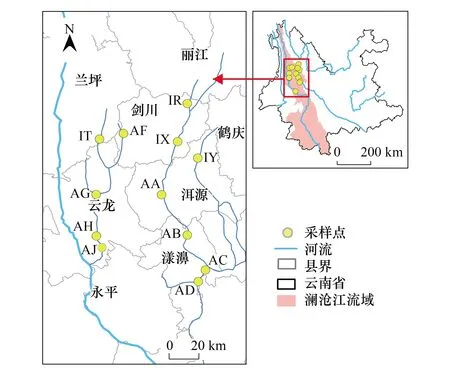
图1 研究区域和采样点布设Fig.1 The study area and sample points distribution 图中IT、IR、IX、IY、AA、AB、AC、AD、AF、AH、AJ为采样点编号,采样点编号根据空间位置和采样顺序设置
1.2 样品采集
为了保证采样点的物候一致和水陆界限分明,于2014年3—4月份进行了样品采集。采样点尽量选择在水流交汇处,以水流汇入点为原点,采集原点上下游各10 m、20 m处对称水(底泥)、陆(土壤)样品共10份(图2),每份样品按五点采样法采集0—10 cm深度的混合土样(或底泥)约50 g置于一次性自封袋中常温保存,并用油性记号笔标记样点编号、采样时间、海拔、经纬度、水或陆等样品信息,采集好的样品于一周内带回实验室并立即开展捕食线虫真菌的分离、纯化工作。所有样品采集由中科院昆明动物所专职采样人员协助完成。

图2 每个采样点的采样示意图Fig.2 Sampling design of each sample point
1.3 菌株的分离纯化
1.3.1培养基的制备
玉米培养基(Cornmal Agar Medium,CMA)、土豆培养基(Potato Dextrose Agar Medium,PDA)以及燕麦培养基均参照文献[31]配制。
1.3.2诱饵线虫的培养及线虫悬浮液的制备
将全齿复活线虫(Panagrellusredivivus)接入燕麦培养基,放恒温培养箱(26.5±1)℃培养一周,直到观察到线虫爬壁即可取出备用或放4℃冰箱保存。采用贝尔曼氏漏斗法[17]制备线虫悬浮液。
1.3.3捕食线虫真菌的分离纯化
将土壤样本0.5—1 g均匀地撒在90 mm的 CMA上(在样品之间留有空隙),加入线虫悬液2 mL(约5000条)线虫以诱导捕食线虫真菌生长,每份土样设3个重复。室温下培养4周,体视显微镜下进行镜检。
观察到捕食线虫真菌孢子后,用无菌牙签挑取单个真菌孢子到直径为60 mm的CMA培养基上,用保鲜膜封口,置于(26.5±1)℃恒温培养箱中培养1周。
1.4 捕食线虫真菌的鉴定
1.4.1形态学鉴定
采用插片法制作捕食线虫真菌临时装片,用奥林巴斯BX53微分干涉显微镜拍取分生孢子、孢子梗、厚垣孢子和捕食器官等形态特征。参照《Nematode-trapping fungi》 和《中国真菌志-三十三卷》进行形态学鉴定。
1.4.2分子生物学鉴定
将分生孢子转接至60 mm的PDA上进行菌丝富集培养,待菌丝富集后,刮取菌丝,用CTAB法提取菌株的DNA,外送至生物测序公司进行DNA测序。通过ITS(internal transcribed spacer region of the ribosomal RNA gene,核糖体RNA上的非转录间隔区)及RPB2(RNA polymerase II core subunit,RNA聚合酶II核心亚基编码基因)序列同源性比对进行分子生物学鉴定。
1.5 数据分析与处理
(1)计算水、陆及全部样品中的物种检出率,计算公式为:
总检出率(OF)=(检出捕食线虫真菌的样品数/总样品数)×100%
水检出率(AOF)=(检出捕食线虫真菌的水体底泥数/总水体底泥数)×100%
陆检出率(TOF)=(检出捕食线虫真菌的陆地土壤数/总陆地土壤数)×100%
物种检出率(SOF)=(检出某物种的样品数/总样品数)×100%
(2)对水、陆及总体捕食线虫真菌物种多样性指数进行计算,比较水、陆间物种多样性差异,物种多样性指数(Shannon-Wiener index,H′)计算公式为:
式中,H′ 为 物种多样性指数;Pi为第i个种所占百分数;S为采样点的物种数
(3)应用Adobe Photoshop 2019绘制沘江和黑惠江研究区域内捕食线虫真菌的物种分布图。
(4)应用RStudio1.3.1093中的“VennDiagram”包绘制水陆间物种组成韦恩图,比较各对称水、陆采样位点间的物种组成差异。
(5)应用Execl和SPASS对数据进行整理及分析。
2 结果与分析
2.1 捕食线虫真菌总检出情况
本研究共分离到捕食线虫真菌2属13种88株,总检出率为47.5%,多样性指数为2.07,其中:陆地土壤中共分离到2属11种45株,检出率为41.67%,多样性指数为1.99;水体底泥分离出1属10种43株,检出率为53.33%,多样性指数为1.93。各物种在水、陆的检出情况详见图3。

图3 捕食线虫真菌水、陆物种检出率比较Fig.3 Comparsion of species′ occurrence frequency of nematode-trapping fungi between terrestrial and aquaticA.为Arthrobotrys属的缩写,Dac.为Dactylellina属的缩写; A. oligospora: 少孢节丛孢; A.conoides: 圆锥节丛孢; A.eudermata: 厚皮单顶孢; A.oudemansii: 秀丽单顶孢; A.longiphora: 长梗单顶孢; A.rutgeriense: 狂带单顶孢; A.cystosporia: 囊孢单顶孢; Dac. Leptospora: 长孢隔指孢; A.globospora: 球状单顶孢; A.superba: 强力节丛孢; A.musiformis: 弯孢节丛孢; A.xiangyunensis: 祥云单顶孢; A.thaumasia: 奇妙单顶孢
2.2 各物种水陆采样点检出情况
研究区域内分离到的捕食线虫真菌中,有3个物种在水、陆检出的样点数相同,分别为Arthrobotrysoligospora、Arthrobotrysoudemansii和Arthrobotryslongiphora。Arthrobotrysglobospora和Arthrobotryssuperba只在水体底泥中检出,为研究区域内水体底泥中特有物种;Arthrobotrysrutgeriense、Arthrobotryscystosporia和Dactylellinaleptospora只在陆地土壤中检出,为研究区域内陆地土壤中特有物种。Arthrobotrysxiangyunensis和Arthrobotrysmusiformis的水体底泥中样点检出数较陆地土壤多,Arthrobotrysconoides、Arthrobotrysthaumasia和Arthrobotryseudermata的陆地土壤样点检出数较水体底泥中多。各个物种在水、陆12个样点中检出的样点数见表1。

表1 捕食线虫真菌检出样点数Table 1 The number of detected sample points of nematode-trapping fungi
2.3 两支流各采样点对称水陆位点间物种组成差异
研究区域内,黑惠江比沘江拥有更高的物种多样性(图4),且两支流捕食线虫真菌在对称水、陆位点间的物种组成均存在较大的差异:

图4 澜沧江两条支流捕食线虫真菌物种分布 Fig.4 Species distribution of nematode-trapping fungi in two tributaries of the Lancang River
沘江共分离出1属9种捕食线虫真菌,其中陆地土壤中分离出1属8种,水体底泥中分离出1属6种,水陆间物种组成相似性指数为0.57。而对沘江各样点对称水、陆间物种组成分析发现:AJ样点水、陆间物种组成完全不同,且陆地土壤中物种多样性高于水体底泥;AG样点水体底泥中物种是陆地土壤中物种的子集;AH和AF两样点的对称水陆间物种组成存在差异的同时也存在一定的相似性;IT样点陆地土壤中物种是水体底泥中物种的子集(图5)。
黑惠江共分离出捕食线虫真菌2属13种,其中陆地土壤中分离出2属10种,水体底泥中分离到1属9种,水陆间物种组成相似性指数为0.63。其中AA点和IR点分别只在陆地土壤和水体底泥中有物种检出;AC和AD两样点的对称水陆间物种组成完全不同,且物种丰富度前者陆地土壤大于水体底泥,后者水体底泥大于陆地土壤;IY、IX和AB三个样点的对称水陆间物种组成存在差异的同时也存在一定的相似性(图5)。
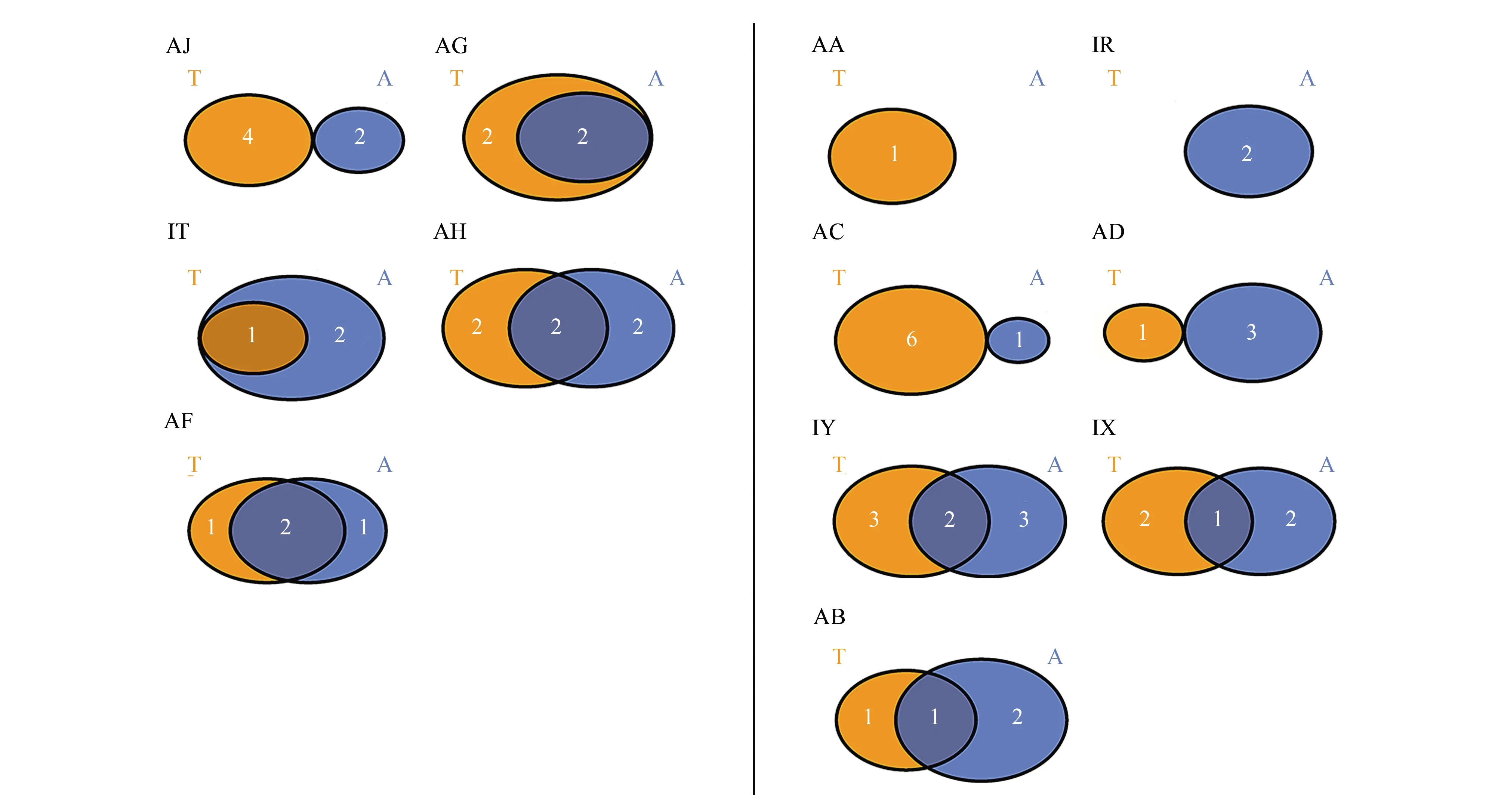
图5 每个采样点水陆间捕食线虫真菌物种组成Fig.5 Species composition of nematode-trapping fungi between terrestrial and aquatic of each sample pointT: 陆地土壤 Terrestrial soil; A: 水体底泥 Aquatic sediment;黄色部分为陆地中检出的物种数,蓝色部分为水中检出的物种数,灰色部分为水陆共有的物种数
3 讨论
3.1 澜沧江支流捕食线虫真菌分布存在水陆差异
研究结果显示,捕食线虫真菌在澜沧江支流的物种多样性分布存在水陆差异:首先,研究区域内水体底泥中捕食线虫真菌物种检出率高于陆地土壤,而陆地土壤中物种多样性则略高于水体底泥;其次,在澜沧江支流分离出的捕食线虫真菌绝大多数为产黏性菌网的Arthrobotrys属真菌,占总物种数的92.31%,但这类捕食线虫真菌在水、陆环境间的物种分布存在一定的差异;第三,捕食线虫真菌群落结构在澜沧江两条支流的水、陆间均存在不同程度的差异。此前有研究者提出,水生生态系统较陆生生态系统有持续的环境梯度和水流均一性[32],水流的均一性使得捕食线虫真菌在水中更容易被检出可能是其在水体底泥中检出率较高的原因,而水陆生境异质性则决定了物种在水、陆间的多样性分布差异。产黏性菌网的Arthrobotrys属真菌以营养要求简单、腐生能力强、生长迅速等特点广泛分布于各种生态环境中[33—35],但这类捕食线虫真菌在水、陆环境间的物种分布存在一定的差异,表明捕食线虫真菌的环境适应能力也存在属内差异,这与范喜杰等[36]对大理温泉中捕食线虫真菌的研究结论一致。此外,在分析沘江和黑惠江各自的物种组成时发现,捕食线虫真菌在澜沧江两条支流间呈现不同的分布模式:两支流的整体群落结构存在差异;两支流水、陆间捕食线虫真菌群落相似性不同,拥有较高物种多样性的黑惠江水、陆间物种相似性更高;同一个物种在不同的样点甚至不同的流域会有不同的水、陆分布特点。这表明流域间及流域内的水生和陆生环境间均存在明显的环境过滤,物种在流域内也同样存在生境选择和生境适应性,而捕食线虫真菌是一个小类群,两生境中物种多样性越高,物种重叠的概率可能就越大。
3.2 捕食线虫真菌水陆分布联系紧密
微生物在流域内分布存在水陆差异的同时其在水陆间也应该是相互联系的,而之前的研究把水生和陆生物种相互割裂开分析,仅有少量的研究做了水生和陆生物种多样性的比较[15—16],但仍缺少水陆关系研究。在对澜沧江支流各采样点的对称水陆样点间捕食线虫真菌物种组成差异分析中发现,捕食线虫真菌在对称水陆样点间物种组成存在明显差异的同时还存在相互联系。尽管捕食线虫真菌是一类水、陆广泛分布的真菌类群,本研究分离得到的物种全部为已知种,且除A.xiangyunensis只在温泉底泥中报道过以外[18],其余物种均在水、陆生境中报道过[17, 26, 37],但本研究区域内不仅有在水体底泥、陆地土壤中均有检出的物种,还有只在于水体底泥或陆地土壤中检出的物种;既有对称水、陆间群落组成存在较大差异的样点,也有水、陆间群落组成相似性较高的样点,表明在流域生态系统内,水、陆生境间存在一定的扩散限制,水流既积极作用于捕食线虫真菌的扩散,同时也形成地理屏障限制了物种的分布。此前有研究指出,水生微生物可能是由于陆生环境中的微生物因河流涨水后长时间的浸泡与冲刷作用而扩散到水中,然后随水流扩散至水生或陆生环境[38—39];长期适应水环境而存在于水生境中的微生物也可能会随水流扩散至陆生环境,最后因水流减少或者水流渗透作用而停留在河岸带[40]。在本研究中,这两种可能性或许都存在,而捕食线虫真菌各物种的扩散能力、环境适应能力和定植能力的差异导致其在水陆环境的分布呈现一定的差异性,表明流域内水生生态系统可通过水文循环驱动流域内环境及生物相互联系,并通过径流和地下水运动与陆地生态系统紧密相连,使得微生物能够在流域内水、陆间横向扩散,从而形成在水陆间的空间分布格局。
3.3 陆地可能是流域内捕食线虫真菌多样性的源
从整体来看,即便有水流的均一性作用,陆地土壤的检出样点数和捕食线虫真菌物种多样性仍高于水体底泥,且有Dactylellina属的检出。这说明和其他微生物一样,陆地土壤也是捕食线虫真菌繁衍的大本营,是其主要的演化场所,因此,陆地土壤可能是捕食线虫真菌在流域内多样性的源。在流域生态系统中,微生物物种多样性分布在水陆间既存在差异又相互联系,水陆间的扩散限制和水流的连通性均对微生物在流域内的扩散和分布极其重要,是流域内微生物物种多样性分布格局产生和维持的重要机制。
4 结论
本研究明晰了捕食线虫真菌在支流流域内水、陆分布差异及其相互关系,发现水、陆不同生境对捕食线虫真菌空间分布格局塑造的作用是不一致的:陆地是捕食线虫真菌的主要演化场所,水流是流域内捕食线虫真菌的重要传播因子。此前大多数流域生态学的研究没有充分考虑流域的水陆耦合作用,建议未来在开展微生物的流域分布格局研究时应考虑各级支流微生物的分布差异,同时兼顾水、陆环境异质性及河流对微生物的扩散和阻隔作用。
致谢:中国科学院昆明动物所陈锦辉老师在样品采集中给予帮助,大理大学创新团队(ZKLX2019103)给予支持,特此致谢。
猜你喜欢
湖北植保(2022年4期)2022-08-23
国画家(2021年4期)2021-10-13
科学与财富(2021年36期)2021-05-10
土壤(2021年1期)2021-03-23
中国农村水利水电(2020年4期)2020-06-12
皮革制作与环保科技(2020年14期)2020-03-17
中国资源综合利用(2017年3期)2018-01-22
福建农业学报(2016年6期)2016-11-01
浙江大学学报(工学版)(2016年9期)2016-06-05
现代企业(2015年6期)2015-02-28
