
基于Maxent模型探讨河流对紫茎泽兰入侵的影响
2022-08-02 07:35瞿佳利谌利民周国兵
杂草学报 2022年1期
杨 洪, 易 文, 瞿佳利, 秦 霞, 谌利民, 周国兵, 陈 艳
(1.绵阳师范学院生态安全与保护四川省重点实验室,四川绵阳 621000; 2.绵阳师范学院生命科学与技术学院,四川绵阳 621000;3.四川省唐家河国家级自然保护区管理处,四川广元 628109; 4.四川省会理第一中学,四川会理 615100)
紫茎泽兰(AgeratinaadenophoraR. King & H. Robinson),又名破坏草、败马草等,原产于墨西哥、乌拉圭、牙买加、哥斯达黎加和古巴一带。最初作为观赏性植物先后被引进到欧洲、澳洲和亚洲,后逸为野生植物[1],目前已经扩散至30多个国家,造成了严重的经济损失和生态破坏,是《外来有害生物的防治和国际生防公约》中四大恶性杂草之一[2]。20世纪40年代紫茎泽兰从缅甸、越南传入我国云南省南部,经过20年的潜伏期后在我国中亚热带湿润气候地区以12 km/年的速度迅速蔓延,而在我国北亚热带地区的扩散速度相对缓慢,但也达到6.8 km/年[3-6],目前已经广泛分布在我国云南、贵州、四川、广西、重庆、湖北、台湾、西藏等省份,并仍然持续向东北蔓延,严重威胁着这些地区的农林业和畜牧业发展,给我国西南地区带来了巨大的生态灾难[1,7-9]。紫茎泽兰可通过根状茎和种子繁殖,种子在其扩散过程中起重要作用。紫茎泽兰的种子产量很高,生长年限为3~5年植株产量达26万~28万粒/m2,种子小而轻,千粒重仅0.04 g[8],同时紫茎泽兰种子具有冠毛,可凭借风力、水力、动物、车载以及无性繁殖等方式扩散[10-12],实地调查和模型模拟均显示河流和道路为紫茎泽兰的扩散提供了良好的通道。调查显示,在滇中地区公路沿线,紫茎泽兰的丰度、植株大小及种子产量在距公路5m处分布时达到峰值[13],在云南省省道公路里程数与紫茎泽兰入侵面积呈显著正相关[14];在金沙江流域,紫茎泽兰入侵的严重程度随距河流及道路的距离增大而减弱[15]。Wang等利用紫茎泽兰在四川、重庆、湖北3个省份部分县(市、区)紫茎泽兰的首次出现时间,构建模型重演其入侵历程,结果显示,多种因素的联合导致紫茎泽兰快速扩散,而河流在其中起重要作用[4,12]。尽管这些研究证实道路和河流为紫茎泽兰的扩散提供了通道和载体,但其周边地区是否同时为紫茎泽兰的定植、繁殖提供适宜的生境还不明晰。
物种分布模型(species distribution models,简称SDM)是研究物种适生性的重要工具[16-17],其中最大熵模型(maximum entropy model,简称Maxent模型) 是目前应用较多的一种模型[18-20],该模型通过计算最大熵的概率分布估计目标物种适生区的分布,能够在较小样本量的条件下获得较高的预测精度,目前已广泛应用于入侵生物的潜在分布区预测[18,21]、濒危物种适生区预测[22-23]以及全球气候变化对物种分布的影响[24-25]等领域。Maxent模型在紫茎泽兰中已有应用,如洪宇辰等预测了紫茎泽兰在四川省的分布,提出四川省中部平原地区应为重点防治区域[26];李丽鹤对紫茎泽兰在云南省的分布进行了预测,证实未来气候条件下紫茎泽兰入侵高风险区分布中心将向高纬度、高海拔迁移,提出应加强对怒江河谷南段、元江河谷、洱海流域、滇池流域等的防控措施[27];张颖模拟了紫茎泽兰在全国的潜在分布,并指出影响其扩散的关键环境变量是年平均气温和潜在蒸腾强度[28]。笔者利用Maxent模型预测紫茎泽兰在我国的适生分布区及关键影响因子,并利用ArcGIS生成主要河流和道路的缓冲区,通过比较不同缓冲区内紫茎泽兰的适生区面积差异,探讨河流和道路缓冲区对紫茎泽兰扩散和定植的作用,以期为紫茎泽兰的防治提供参考。
1 材料与方法
1.1 数据来源
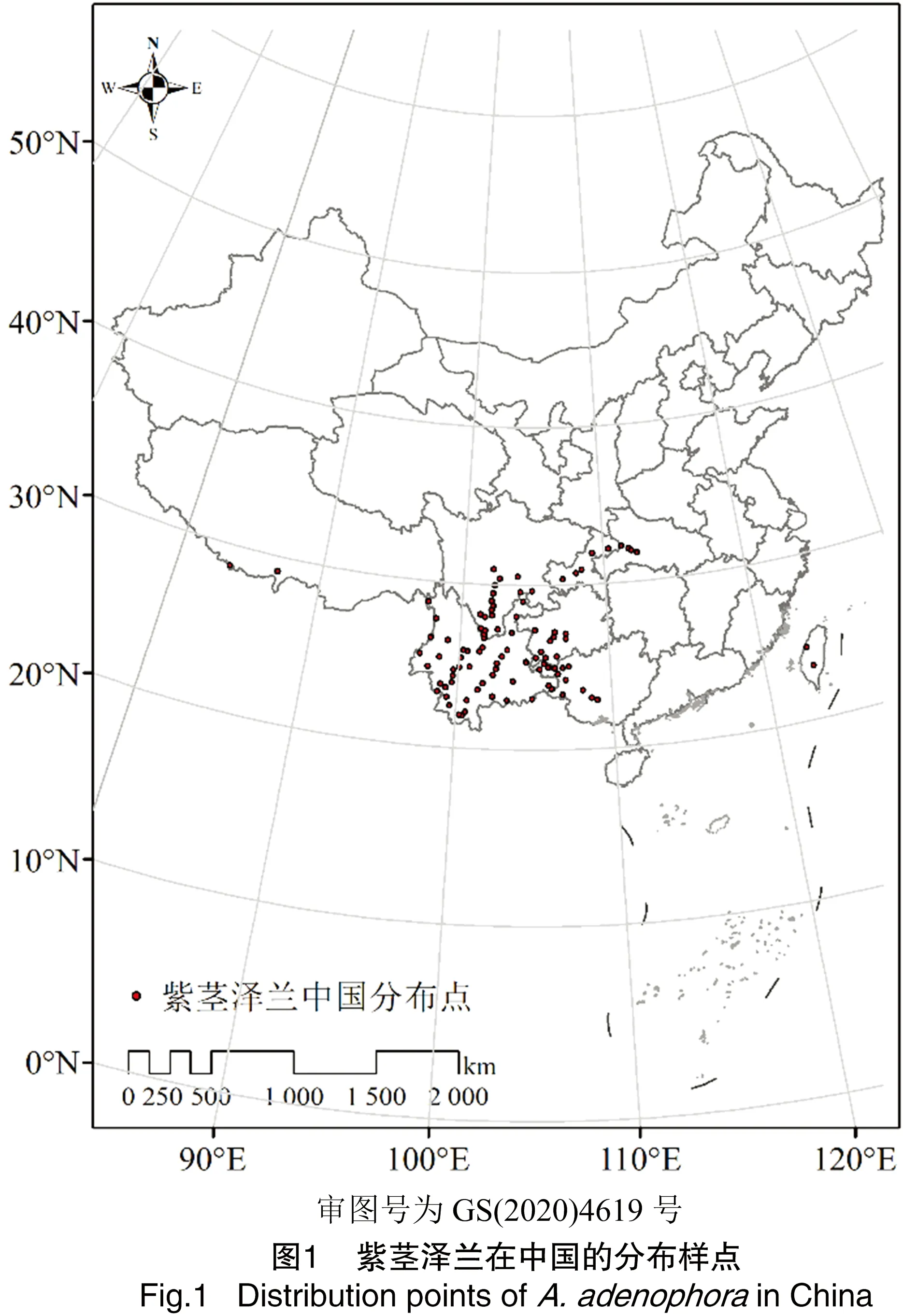
1.1.1 紫茎泽兰分布数据 紫茎泽兰分布的数据主要采取3种方式收集:(1)对中国数字植物标本馆(https://www.cvh.ac.cn/)和全球生物多样性信息平台(https://www.gbif.org/)进行检索。(2)查询中国知网、万方等有关紫茎泽兰的文献(报道)和政府网站的新闻消息,通过“91卫图助手”进行坐标拾取和校准。(3)实地调查获取四川、云南等地紫茎泽兰的分布数据。将获取的所有分布数据导入ArcGIS 10.5,去除国界外分布点和重复信息,保留紫茎泽兰分布样点101个(图1)。
1.1.2 环境变量与地理信息数据 本研究共选择年平均气温、月平均日较差、等温性、温度季节性变化、最暖月最高气温、最冷月最低气温、温度年变化范围、最湿季平均气温、最干季平均气温、最暖季平均气温、最冷季平均气温、年降水量、最湿月降水量、最干月降水量、降水季节变异系数、最湿季降水量、最干季降水量、最暖季降水量、最冷季降水量、各月平均气温、各月平均降水量、海拔等共44个环境变量。环境变量数据(1970—2000年)通过全球气候数据库(worldclim global climate and weather data version 2.1,https://www.worldclim.org/)下载分辨率为30″(地理空间分辨率约为1 km)的全球气候数据,再与中国行政区划数据插值得到。行政区划(1 ∶4 000 000)、水系分布、主要公路等数据来自于中国国家基础地理信息系统(http:∥nfgis.nsdi.gov.cn)。
1.2 模型模拟
1.2 .1 主导环境变量筛选 Maxent模型是目前具有最佳预测能力和精度的生态位模型[18,29],将上述101个紫茎泽兰分布点数据和44个环境变量导入模型,选取75%的紫茎泽兰分布数据作为训练集建立预测模型,25%的紫茎泽兰分布数据作为测试集验证模型,重复10次,其余参数为默认值[30]。
通过刀切法结果(jackknife test)和环境变量的综合得分评价环境变量对模型的相对贡献,筛选关键环境变量,其中综合得分通过赋予贡献率(percent contribution)和置换重要性(permutation importance)各50%的权重计算得到[31-32];运用 R 3.60 计算关键环境变量两两间的Pearson相关系数[33-35],当环境变量之间的相关性系数绝对值≥0.8,则认为二者具有很强的相关性[36],对于相关性>0.8的环境变量,结合综合得分保留相对贡献较大的变量。
1.2.2 紫茎泽兰适生区模拟 依据筛选出的主导环境变量再次运行模型,各参数设置不变,采用受试者工作特征曲线(receiver operating characteristic curve,ROC曲线)下面积值即AUC值(area under the curve)来评价模型预测准确度[37]。AUC值的取值范围为0~1,值越接近1表明环境变量与分布模型之间的相关性越大,预测精度越高,当AUC值≥0.9时,预测效果达到极佳[35,38]。
Maxent模型输出结果为紫茎泽兰在中国范围内的存在概率(0≤P≤1),数据格式为ASCⅡ格式,运用ArcGIS进行格式转换及重分类操作,将结果分为4个不同等级的综合评价等级:(1)P≤ 0.1为非适生区;(2) 0.1
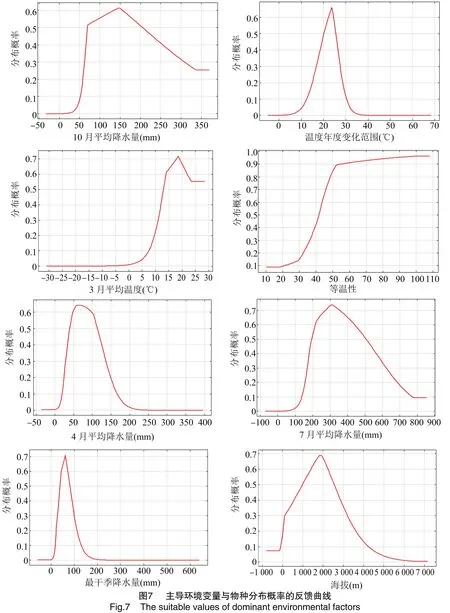
Maxent模型可以体现主导环境变量和物种分布概率之间的关系,通过环境变量与物种分布概率之间的反馈曲线来反映不同阈值下的环境变量取值范围。按照本研究分布概率等级划分,以P=0.3为临界值,分析紫茎泽兰适生生境的主导环境变量的最适范围。
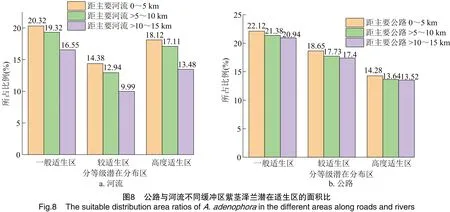
1.2.3 河流道路缓冲区分析 利用ArcGIS软件,分别对全国4级水系数据和4级公路数据与紫茎泽兰预测结果执行相交分析,获得结果再进行缓冲区分析[40],统计距河流或道路近距离0~5 km、中距离>5~10 km、远距离>10~15 km缓冲区范围内紫茎泽兰不同适宜等级的面积,分析距河流和道路距离对紫茎泽兰分布的影响。
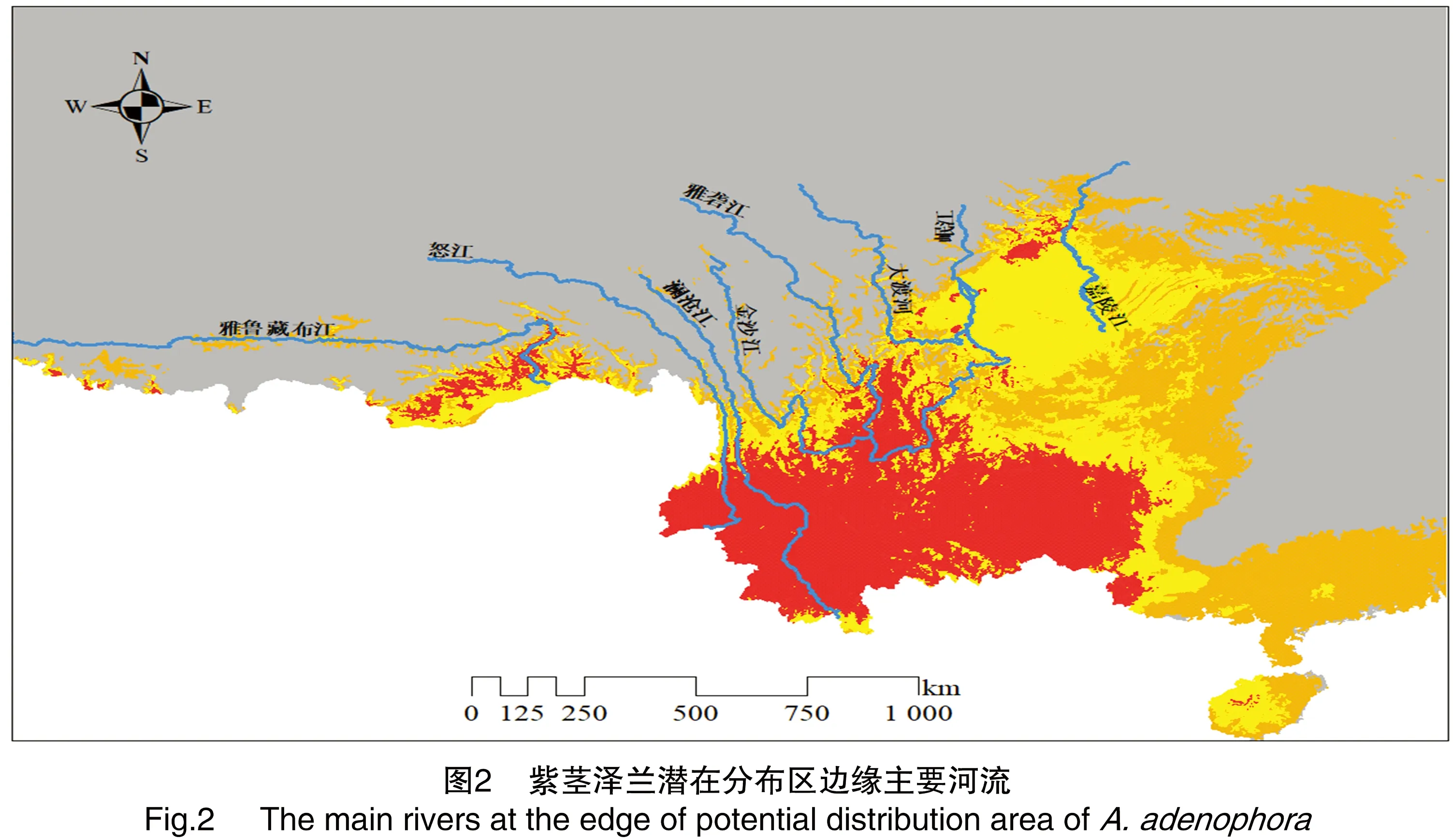
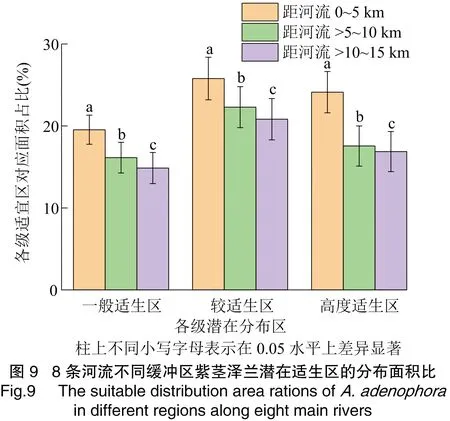
为检验河流3个缓冲区(分别距河流0~5、>5~10、>10~15 km)紫茎泽兰生境适宜性的差异,选择扩散前沿的雅鲁藏布江、怒江、澜沧江、金沙江、雅砻江、大渡河、岷江、嘉陵江等8条主要河流(图2),对每条河流进行上述缓冲分析、相交分析、面积融合、面积统计处理,分别统计每条河流3个缓冲区域紫茎泽兰各适生等级的面积,将各等级适生区面积转化为对应缓冲区范围入侵总面积的百分比。在R软件中对河流不同缓冲区内每个适生等级的面积比进行配对样品t检验。
2 结果与分析
2.1 主导环境变量筛选
结合初始模型刀切法结果(图3、表1),AUC值得分(图4)及相关性分析结果,筛选出8个主导环境变量(表2)。根据筛选后变量重新构建模型,运行模型后得到其贡献率和置换重要性,结果见表2。从模型结果来看,10月平均降水量、等温性和3月平均气温对紫茎泽兰分布的贡献最大,主导环境变量与紫茎泽兰的分布呈正相关。在8个主导环境变量中与降水相关的变量累积贡献率达68.2%,包括4、7、10月平均降水量及最干季降水量,说明降水明显影响了紫茎泽兰的分布。
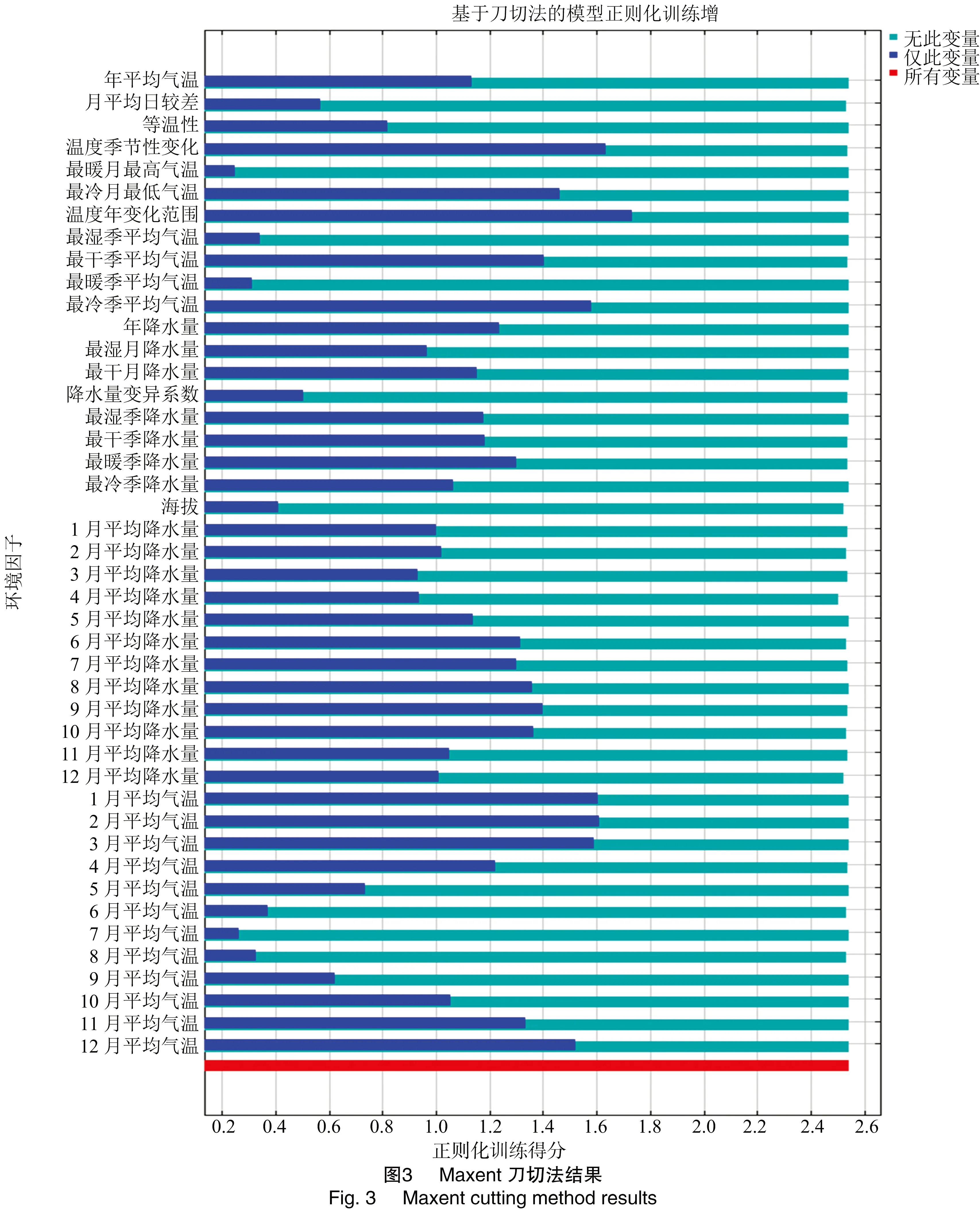
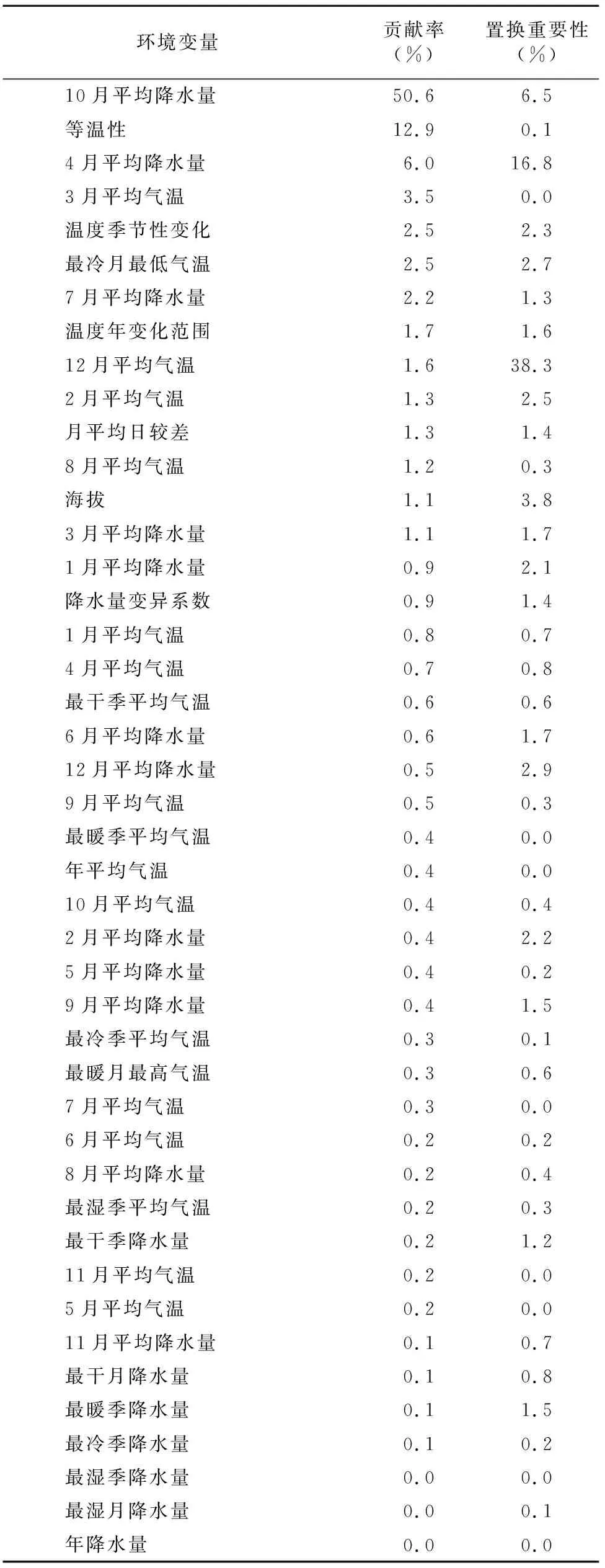
表1 环境变量对紫茎泽兰分布重要性Table 1 Importance of environmental variables on the distribution of Ageratina adenophora
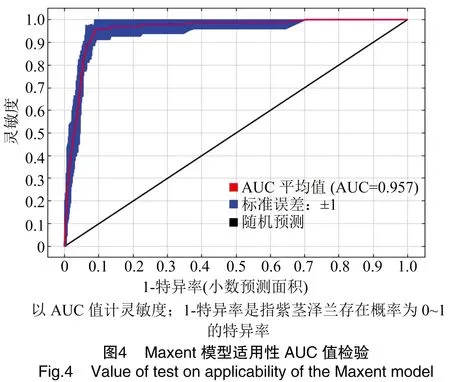
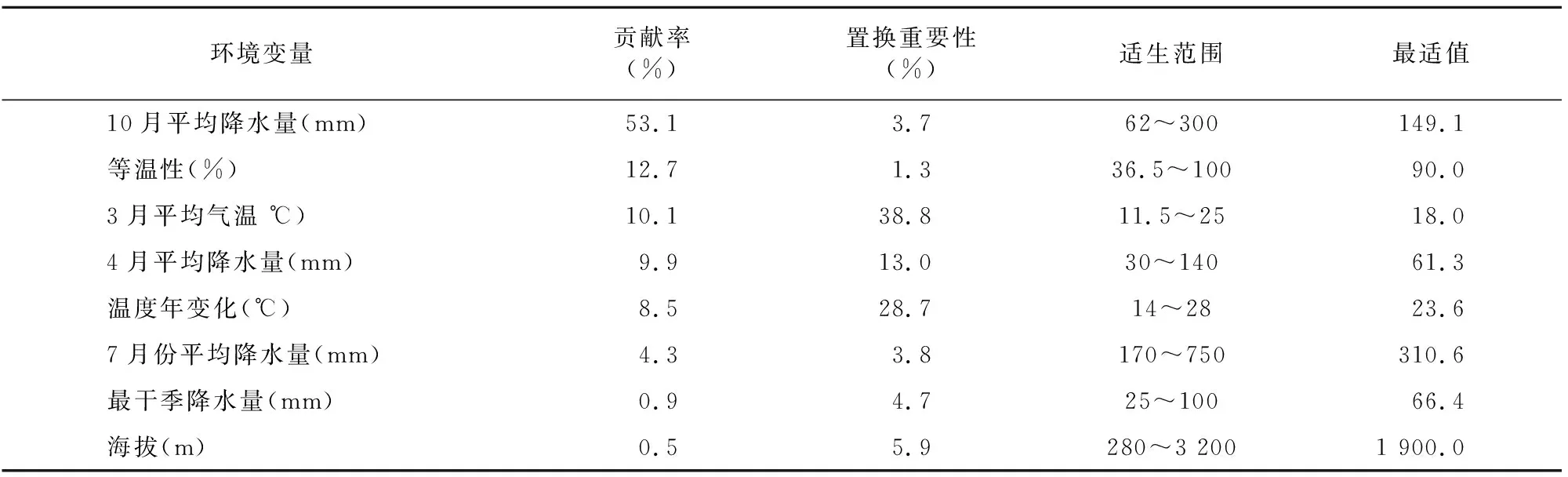
表2 主导环境变量贡献率与适生范围Table 2 Contribution rate and suitable range of leading environmental factors
2.2 紫茎泽兰在中国的地理分布预测结果
模型平均AUC值为0.959±0.014,表明模型预测可靠性高,可以应用于紫茎泽兰适生区划。紫茎泽兰在中国潜在分布区4种适宜等级(非适生区、一般适生区、较适生区、高度适生区)分布情况见图5。
紫茎泽兰在当前气候条件下在中国的潜在分布范围主要集中在中国西南部和东南地区,跨越较广泛, 在80.05°~121.50°E、18.22°~33.88°N区间都有分布,涉及省份有云南、贵州、四川、广西、西藏、广东、福建、海南、台湾、重庆、湖北、陕西、甘肃。
在目前的气候模型条件下,紫茎泽兰预测潜在分布区总面积达到139.42×104km2,占国土总面积的14.52%,其中高度适生区、较适生区、一般适生区面积分别为40.67×104、37.48×104、61.27×104km2。高度适生区主要分布在云南全境、广西西部、贵州黔西南州、四川凉山州、西藏东南部;其中,云南、四川、贵州、广西的高度适生区面积占比很大,达到全部高度适生区面积的95.94%(图6-a);较适生区主要集中在四川、云南、贵州、广西、西藏、和重庆,其中云南、四川、贵州和广西的较适生区面积达到全国较适生区面积的75.21%(图6-b)。紫茎泽兰潜在适生区在各省的分布见表3。
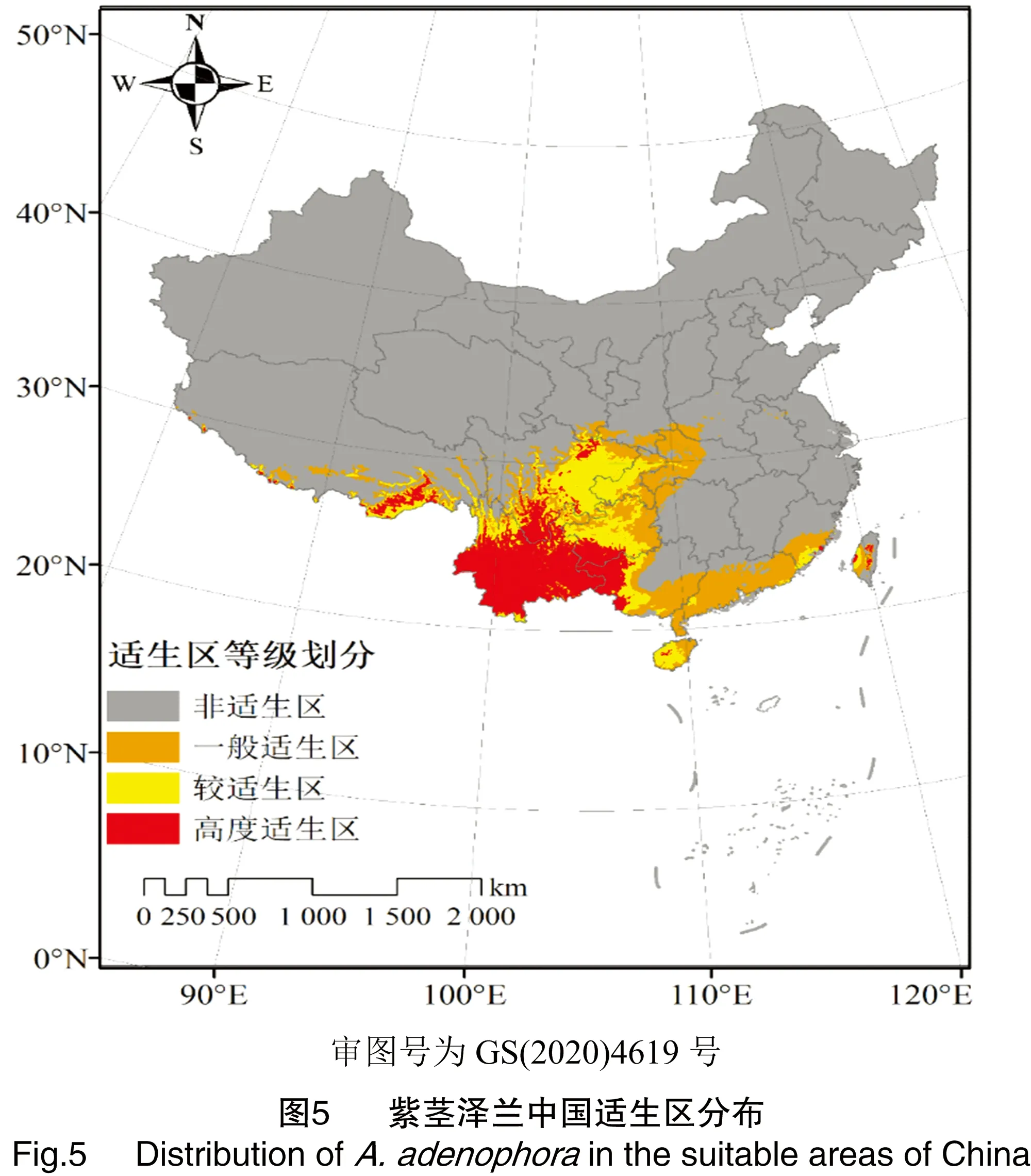
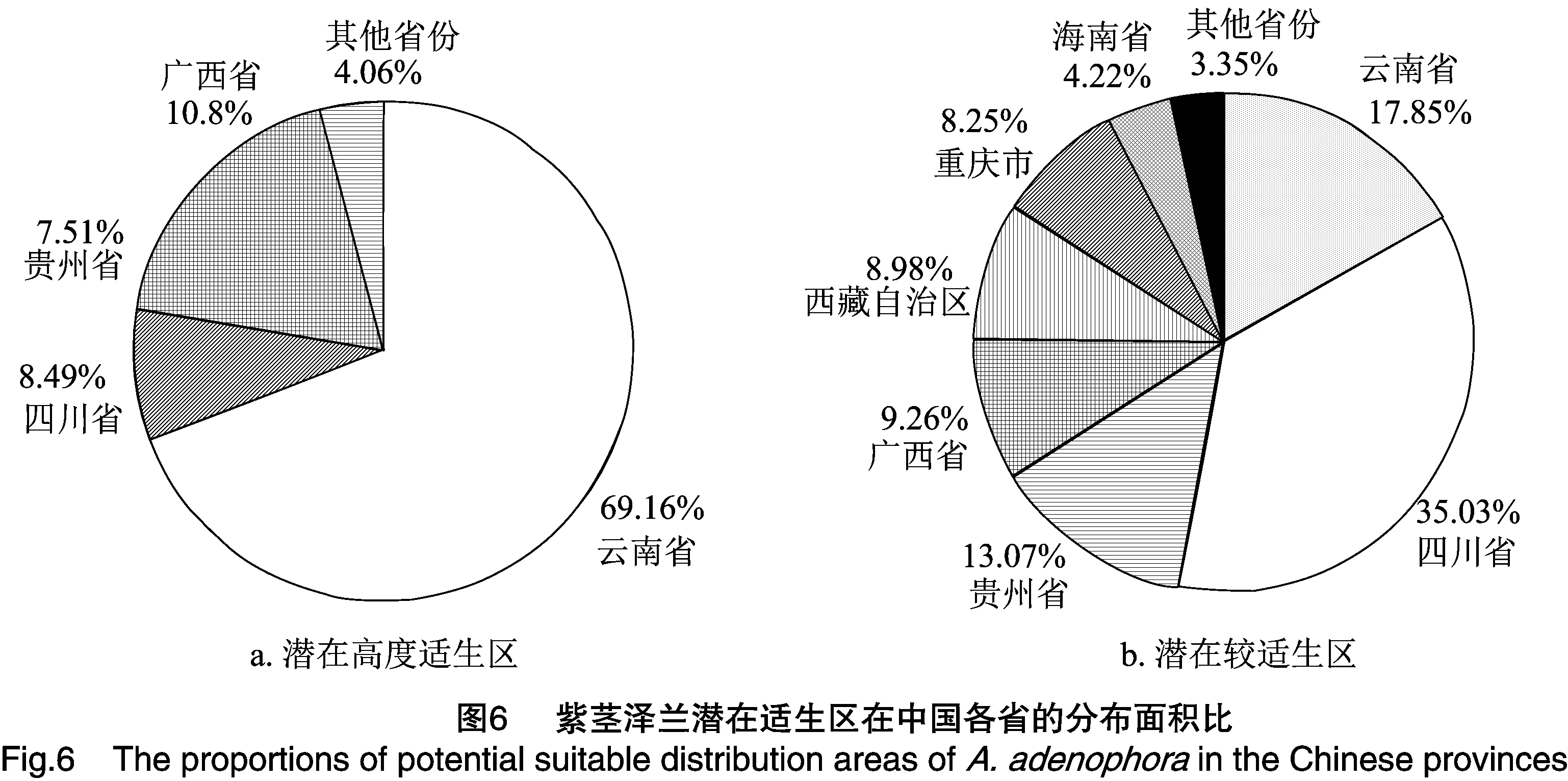
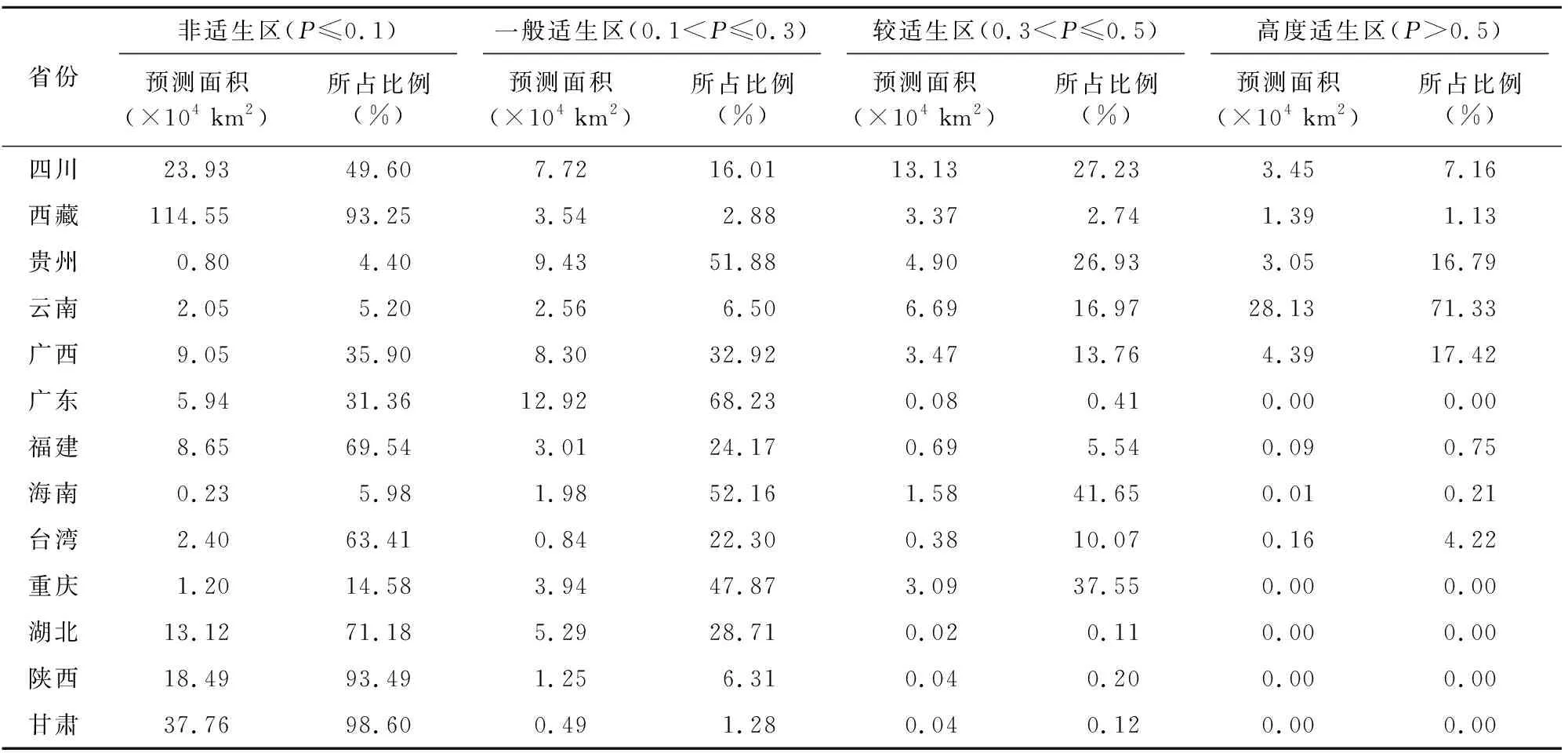
表3 紫茎泽兰在各省潜在分布面积及其占各省面积的比例Table 3 Potential distribution area of A. adenophora in each province and its proportion in the area of each province
2.3 关键环境变量的最适范围
环境变量与物种分布概率的反馈曲线(图7)显示,10月平均降水量的适宜范围在62~300 mm之间,最适值为149.1 mm,在62~149.1 mm之间的分布概率与降水量呈正相关,149.1~300 mm之间呈负相关。温度年变化范围的适宜范围在 14~28 ℃之间,最适值为23.6 ℃,其中在14~23.6 ℃ 之间,分布概率与温度年变化范围呈正相关,在23.6~28 ℃之间,分布概率与温度年变化范围呈负相关;3月平均气温在大于11.5 ℃时适宜,但在11.5~18 ℃之间,分布概率与平均气温呈正相关,18~25 ℃呈负相关,超过25 ℃无明显变化;其余环境变量与分布概率的关系与上述情况类似(表1),但是变化幅度和取值范围存在差异。在分布范围内的环境变量变化对紫茎泽兰的分布情况产生一定的影响,对取值在适宜范围之外时的环境变量对紫茎泽兰的分布概率影响逐渐降低。
2.4 河流与公路对紫茎泽兰分布的影响
河流、道路不同缓冲区紫茎泽兰各等级适生区的占比见图8,随着距河流、公路距离的增加,紫茎泽兰高度适生区、较适宜区、一般适生区面积所在比例均逐步降低。距河流距离每增加5 km,一般适生区、较适生区和高度适生区的面积比分别减少(1.88±0.62)、(2.20±0.53)、(2.32±0.93)百分点;而距公路的距离每增加5 km,对应适生区的面积比分别减少(0.59±0.11)、(0.63±0.21)、(0.38±0.18)百分点,河流缓冲区的适生面积比降低更快,其适应性的变化远大于道路缓冲区。对紫茎泽兰扩散前沿8条主要河流不同缓冲区紫茎泽兰适宜生境的统计也显示,随着距河流距离的增加,紫茎泽兰各等级适宜生境面积比显著降低(图9)。
3 讨论与结论
随着经济贸易和交通的全球一体化,物种跨越原有生物地理障碍的扩散情况日趋严重,外来入侵物种在世界各地都呈现出快速膨胀的趋势,给入侵地带来巨大的生态危害和经济损失[41-42],目前生物入侵已成为全球生物多样性丧失的重要诱因[43],对入侵地生态系统有长久的影响,因而入侵生物丰度和分布的变化是生态系统管理不可或缺的信息[10]。Maxent模型预测结果显示,紫茎泽兰在中国的潜在分布区总面积高达139.42×104km2,在80.05°~ 121.50°E、18.22°~33.88°N都有分布,其中云南、四川、贵州、广西的高度适生区面积占比达95.94%,这与鲁萍等的研究结果[9,44-46]一致,在紫茎泽兰的防治工作中,需特别加强对这些地区的监测和管理。
入侵生物扩散和定植动态为生物入侵防治策略的制定提供了科学依据,但入侵生物的扩散和定植的影响因素与机制仍是生物入侵研究的巨大挑战。紫茎泽兰可通过根状茎和种子扩散,且种子小而轻,具冠毛,这些生物学特性增强了紫茎泽兰种群的扩张能力[1,7],而道路和河流为紫茎泽兰的扩散提供了良好的通道[4,12,14-15]。笔者对道路、河流缓冲区内紫茎泽兰适应生境面积的统计显示,随着距道路、河流距离的增加,紫茎泽兰适生区的比例逐步降低(图5)。而随着距河流距离的变化高于随距公路距离的变化,距离每增加5 km,河流公路紫茎泽兰高度适生区占比分别降低(2.32±0.93)、(0.38±0.18)百分点,较适生区分别降低(2.20±0.53)、(0.63±0.21)百分点,一般适生区分别降低(1.88±0.62)、(0.59±0.11)百分点;利用紫茎泽兰种群扩散前沿8条河流的检验显示,紫茎泽兰适生区比例随河流距离的增加而降低且已达到显著水平。这与Wang等的研究结果[10,12]一致,在紫茎泽兰的扩散中河流较道路起到更重要的作用,说明尽管河流和道路均为紫茎泽兰提供扩散通道,但河流可能更有利于紫茎泽兰的定植。
生境的可入侵性直接影响着入侵物种的定植,结构复杂、生物多样性高的生态系统对生物入侵有较高的抗性[47]。道路建设和维护均会破坏道路两侧的生态系统,而洪灾等自然灾害也会破坏河流两侧的生态系统,道路和河流两侧的裸地和受干扰的生态系统为紫茎泽兰的定植提供了可入侵生境。对道路、河流缓冲区的比较显示,紫茎泽兰适生区的降低在河流缓冲区更为明显,说明河流缓冲区除了提供可入侵生境外,可能还为紫茎泽兰的存活和繁殖提供其他有利条件。
Maxent模型的模拟结果显示,紫茎泽兰的分布概率随8个主导环境变量(10月平均降水量、等温性、3月平均温度、4月平均降水量、温度年变化范围、7月平均降水量、最干季降水量和海拔)的变化而改变,其中10月降水量、温度年变化范围和3月平均温度是最为重要的影响变量,有研究证明温度和降水极大地影响紫茎泽兰的分布,紫茎泽兰主要分布在年平均气温10~20 ℃、年降水量800~2 000 mm的地区[5,48]。紫茎泽兰种子活力高,在湿度较高的条件下能够迅速发芽生长,实现入侵[49],高温和低温都会抑制紫茎泽兰种子萌发的与幼苗的生长,从而限制了紫茎泽兰的传播[50],其中3月的最高温度对紫茎泽兰的传播限制最大[51];而冬季相对较高的温度有助于紫茎泽兰的越冬,并提高其竞争能力[52]。河流和湖泊都可以调节周围地区的局部小气候,夏季,河流水量丰富,蒸发旺盛,使局部地区降雨增多,同时蒸发时吸热,因此可以降低气温。在冬季,陆地降温快,而河流湖泊降温慢,可以提高局部温度[53-55],因此河流在紫茎泽兰早期的扩散中,可能不仅仅为种子传播提供通道,同时还为种子萌发、幼苗生长提供有利条件,促进紫茎泽兰的快速入侵。
运用Maxent模型预测紫茎泽兰在中国的潜在分布虽然具有操作简便、样本量需求小、预测精度高等优点,但也不可避免地有其他类似生态位预测模型一样的缺点,模型模拟的情况是一种理想状态下的物种分布概率,不考虑天敌分布等实际情况[56],所以模拟出的分布区域大于实际分布区域。另外,Maxent预测模型是一种理想的生态位模型,仅考虑了非生物因素的影响。而影响物种分布的因素有很多,包括生物因素和非生物因素等,任何给定物种的分布模式都是其生物特征和生态条件随时间相互作用的结果[57-58]。在后续研究中,须要更加关注生物变量与非生物变量的交互作用,探讨其如何影响紫茎泽兰的分布和种群动态,进一步为紫茎泽兰的科学防治提供参考。
猜你喜欢
导航定位学报(2022年4期)2022-08-15
小天使·三年级语数英综合(2022年4期)2022-04-28
小太阳画报(2019年4期)2019-06-11
散文诗(2018年20期)2018-05-06
汽车导报(2017年5期)2017-08-03
中国新通信(2017年3期)2017-03-11
中学生数理化·高二版(2016年4期)2016-05-14
少儿科学周刊·少年版(2015年11期)2015-12-17
时代人物(2014年10期)2015-01-28
环球时报(2012-01-12)2012-01-12
