
雅砻江中游支流力丘河鱼类资源现状
2022-07-28 08:26:32邓嘉俊王小东同琳钿徐亮亮向朋孙光肖能文宋昭彬
四川动物 2022年4期
邓嘉俊,王小东,同琳钿,徐亮亮,向朋,孙光,肖能文,宋昭彬,3*
(1.四川大学生命科学学院,生物资源与生态环境教育部重点实验室,成都 610065;2.中国环境科学研究院,环境基准与风险评估国家重点实验室,北京 100012;3.四川大学生命科学学院,长江上游鱼类资源与环境四川省野外科学观测研究站,成都 610065)
雅砻江是金沙江最大的支流,全长1 570 km,流域面积13.6×10km。流域内水力资源丰富,水能理论蕴藏量为3 000×10kW。迄今为止,雅砻江下游锦屏一级、锦屏二级、官地、二滩和桐子林5座水电站以及中游两河口、杨房沟2座水电站已投产发电。雅砻江干流大规模的水电开发必将对该流域的水生生物资源,特别是鱼类资源产生深远的影响(陈云华,2021)。
历史上,雅砻江的鱼类资源非常丰富。在20世纪80年代开展的鱼类资源调查中,邓其祥(1985)实际采集到72种鱼类,而吴江和吴明森(1986)结合历史资料及现场考察,认为雅砻江存在92种鱼类。随后,在1983—1993年对雅砻江中、下游的系统调查中,邓其祥(1996)共采集到94种鱼类。然而,2007—2008年对雅砻江中、下游开展的调查发现,鱼类种类数量下降到75种(杨青瑞等,2011)。除了种类数量下降外,水电开发还对鱼类种类组成产生了巨大影响。二滩水电站修建前雅砻江下游的优势类群为裂腹鱼类、高原鳅类等适应流水生境的种类,随着水坝的修建和水库的形成,河段内的急流生境转变为静水缓流生境,原有的优势类群缩至库尾和支流等狭窄生境,裂腹鱼类的捕获量由71.9%下降至16.0%,取而代之的是外来鱼类及小杂鱼等(蒋红等,2007)。支流生境保护是一种较为有效的生态补偿措施(高婷等,2012;Pracheil,2013;黄光明等,2015)。通过对支流流水生境的保护,干流受水电开发影响的鱼类可在支流得到一定的保护。随着两河口、杨房沟等水电站的建成运行,雅砻江中游河段亟需划定合适的栖息地保护干流受影响的鱼类。
力丘河发源于四川省甘孜藏族自治州康定市塔公镇西部,流经康定市新都桥、瓦泽乡、呷巴乡、甲根坝乡、朋布西乡、沙德乡,在康定市普沙绒乡境内注入雅砻江,全长206 km,自然落差1 880 m,流域面积 5 920 km,年均流量 148 m·s。力丘河中上游海拔较高,瓦泽乡以上河段流经较为平坦的高原,河道曲折、河床宽阔,中游多为宽谷,下游多峡谷分布,海拔落差较大,河口海拔约2 400 m。色物绒沟是力丘河最大的支流,虽然沟口位于力丘河下游,但其大部分河段位于海拔3 300 m以上的区域。塔拉沟为力丘河第二大支流,沟口位于力丘河中游。力丘河是雅砻江中游唯一河流连通性保存较好、尚无水坝修建的较大支流。良好的河流连通性是支流得以发挥鱼类多样性保护作用的必要条件(杨青瑞等,2015),在连通性较好的河段,鱼类可以在不同栖息地之间自由迁移,从而满足不同生活史阶段的生境需要。姜跃良等(2015)认为力丘河、色物绒沟等具备良好的生境质量,适合作为高原鱼类栖息地保护水域。
然而,姜跃良等(2015)的评价主要考虑河流底质、栖境复杂性、堤岸稳定性等生境尺度的指标,而缺乏河流内水生生物多样性的数据支撑。此外,海拔对高原鱼类的分布往往也具有决定性作用,许多生物地理学研究认为同一流域内不同海拔的河段中鱼类种类组成及分布范围差异显著(Súarez,2011;Singh&Agarwal,2013)。为了掌握力丘河的鱼类种类组成、分布特征及资源现状,2019—2020年对力丘河流域进行了系统的鱼类调查,分析了鱼类多样性、空间结构及其与海拔的关系,并探讨了将其作为雅砻江中游干流水电开发后鱼类栖息地保护的价值。
1 材料与方法
1.1 样本采集
本次调查在力丘河共设置11个采样点,其中,干流设置9个采样点,第一大支流色物绒沟和第二大支流塔拉沟各设置1个采样点(表1,图1),基本覆盖了力丘河干流及支流不同的河段及生境。
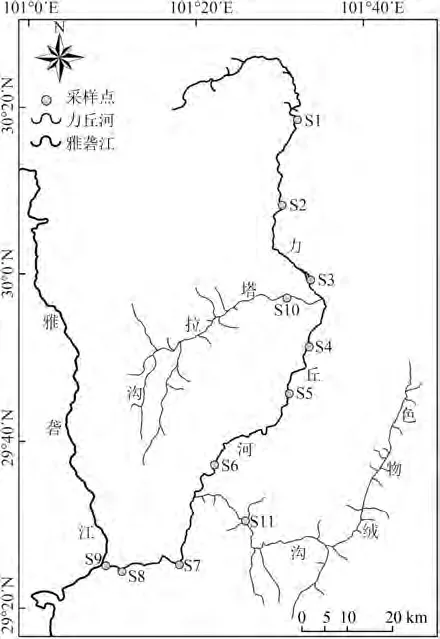
图1 雅砻江中游支流力丘河鱼类资源调查采样点Fig.1 Sampling sites of fish investigation in the Liqiu River,a tributary of the Middle Yalong River
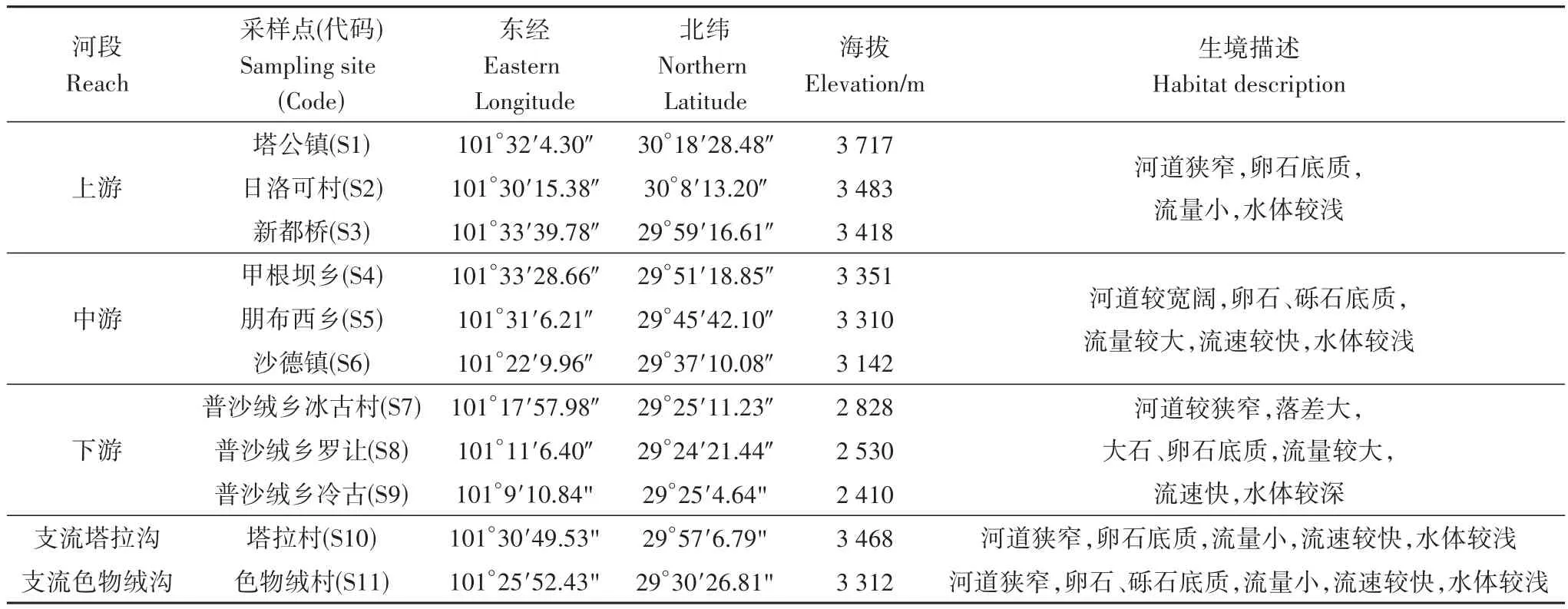
表1 雅砻江中游支流力丘河鱼类资源调查采样点信息Table 1 Sampling sites of fish investigation in the Liqiu River,a tributary of the Middle Yalong River
调查时间为2019年9月和2020年6月、11月。采用地笼(长10 m、宽0.30 m、高0.25 m,网目50 mm)捕捞样本,每个采样点放置4个,所有地笼均放置过夜,采样时间约12 h,且连续采样2~3 d。对所采集的样本进行常规生物学测定和统计。在满足鱼类标本和组织样品等采集的前提下,剩余的渔获样本完成物种鉴定和测量后放回采集水域。采集的鱼类标本用8%~10%甲醛或95%乙醇保存。依据《四川鱼类志》(丁瑞华,1994)、《中国动物志硬骨鱼纲鲤形目(中卷)》(陈宜瑜等,1998)、《中国动物志硬骨鱼纲鲇形目》(褚新洛等,1999)、《中国动物志硬骨鱼纲鲤形目(下卷)》(乐佩琦等,2000)等进行种类鉴定。采用便携式水质检测仪测定水温、电导率和总溶解固体等常规水质指标,使用LS1206B流速仪测量河流流速及水深,使用手持测距仪测量河道宽度,使用图新地球4(LocaSpace Viewer)记录各采样点海拔。
1.2 数据分析
1.2.1 群落优势种 采用Pinkas相对重要性指数(index of relative importance,IRI)(Pinkas,1971)划定力丘河流域鱼类群落的优势种,IRI()10,式中,为某一种类的尾数占总尾数的百分比,为某一种类的质量占总质量的百分比,为某一种类出现的频率。IRI≥1 000为优势种,1 000>IRI≥100为常见种,100>IRI≥10为一般种,IRI<10为偶见种(代梦梦等,2019)。
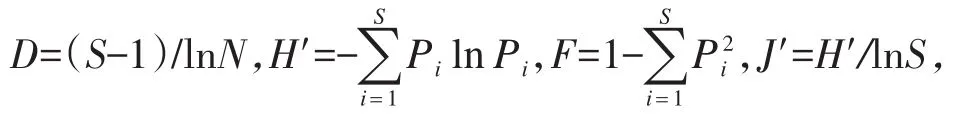
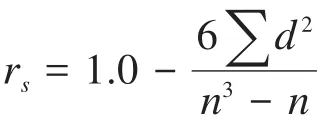
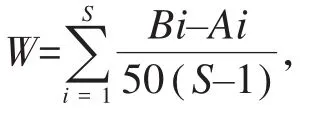
1.2.4 群落结构多元统计分析 运用Bray-Curtis相似性指数计算不同河段间的鱼类群落结构差异性(Clarke&Warwick,1994b),得到鱼类群落相似性矩阵;对相似性矩阵进行等级聚类分析,并绘制聚类分析图;利用非度量多维标度(non-metric multidimensional scaling,NMDS)分析不同河段鱼类群落间的结构特征,并绘制平面图,可信度由胁强系数衡量(Clarke,1993)。
运用相似性分析检验对鱼类群落的时空变化进行显着性检验,对差异明显的2个群落,运用相似性百分比分析方法(similarity percentages,SIMPER)得出造成群落时空差异的鱼类种类。
以上群落结构特征均使用PRIMER 5进行计算。
所有数据均使用SPSS进行统计分析。
2 结果
2.1 鱼类种类组成

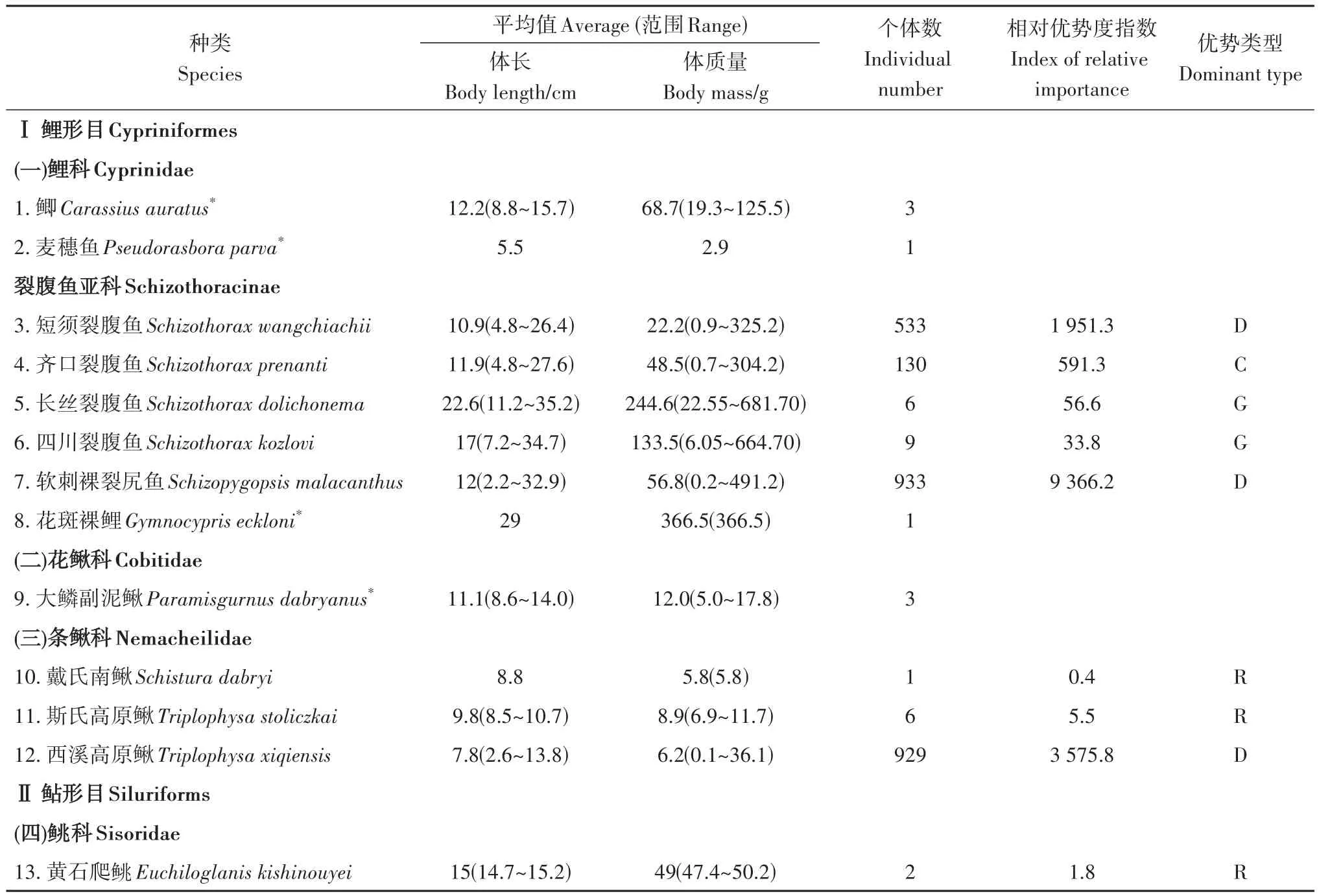
表2 雅砻江中游支流力丘河的鱼类组成Table 2 Composition of fish in the Liqiu River,a tributary of the Middle Yalong River
2.2 鱼类多样性
力丘河的鱼类多样性整体较低(表3):干流的Margalef种类丰富度指数、Shannon-Wiener多样性指数和Simpson优势度指数均高于支流,而直流色物绒沟的Pielou均匀度指数高于干流;从干流的上游到中游、下游,Margalef种类丰富度指数、Shannon-Wiener多样性指数和Simpson优势度指数逐渐上升,而Pielou均匀度指数则逐渐下降。

表3 雅砻江中游支流力丘河的鱼类多样性Table 3 Fish diversity in the Liqiu River,a tributary of the Middle Yalong River
Spearman秩相关性系数表明,物种数、Margalef种类丰富度指数和海拔呈极显著负相关,Pielou均匀度指数和河道宽度呈显著负相关,群落多样性与其他环境因子间无显著相关性(表4)。
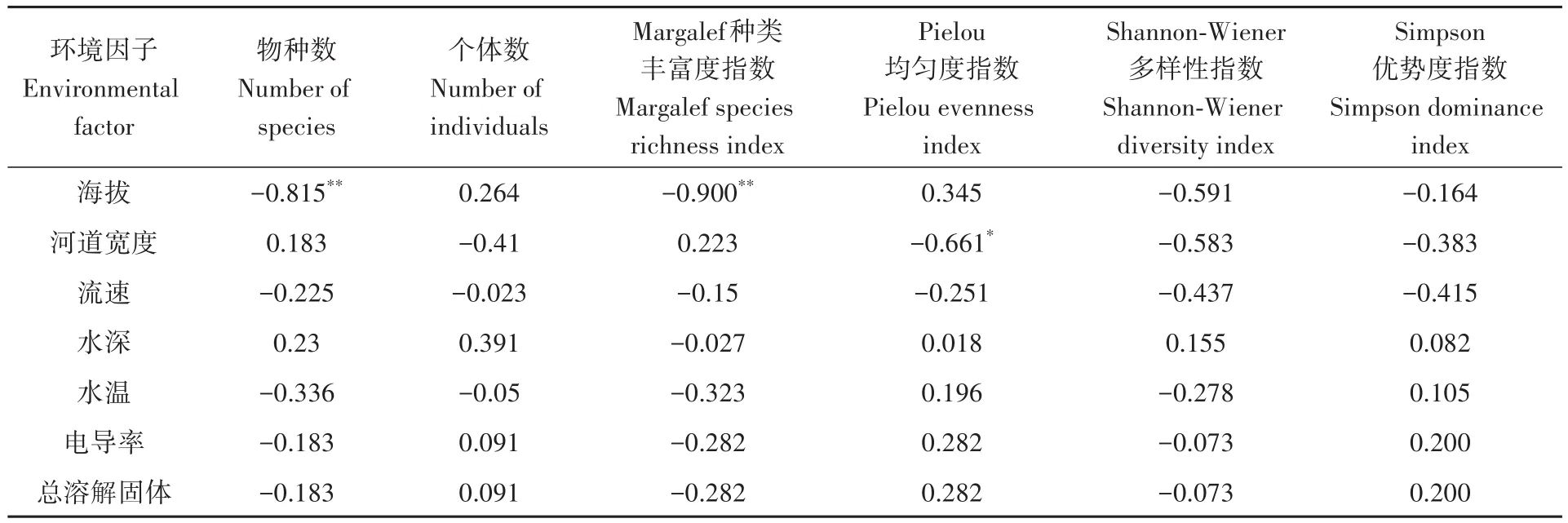
表4 雅砻江中游支流力丘河鱼类多样性与环境因子的Spearman秩相关性系数Table 4 Spearman’s rank correlation coefficient between fish diversity and environmental factor in the Liqiu River,a tributary of the Middle Yalong River
2.3 丰度/生物量比较曲线
力丘河的丰度/生物量比较曲线如图2,其中横坐标为丰度优势度及生物量优势度由高至低排序的种类序号,纵坐标为丰度及生物量优势度的累计百分比。丰度优势度曲线与生物量优势度曲线交叉(>0),说明鱼类群落正处于中等程度干扰。
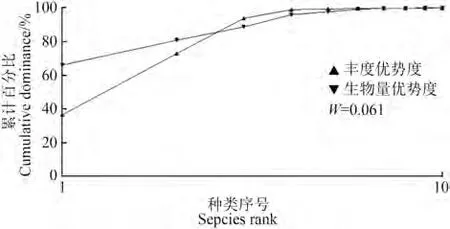
图2 雅砻江中游支流力丘河鱼类丰度/生物量曲线Fig.2 Abundance/biomass curve of fish in the Liqiu River,a tributary of the Middle Yalong River
2.4 群落空间结构
聚类分析和NMDS分析均将不同采样点的群落分为3个组群:组群Ⅰ包括采样点S1、S2、S3、S4、S10和 S11,组群Ⅱ包括采样点 S5、S6和 S7,组群Ⅲ包括采样点S8和S9(图3,图4)。相似性分析检验(ANOSIM)显示不同组群间差异极显著(Global R=0.965,<0.001)。结合采样点地理位置可知,组群Ⅰ、组群Ⅱ、组群Ⅲ分别代表来自上游、中游、下游3个河段的鱼类群落。虽然色物绒沟是力丘河下游的支流,但色物绒沟(采样点S11)的鱼类与力丘河上游(采样点S1~S4)的鱼类一起被划分入组群Ⅰ。

图3 雅砻江中游支流力丘河鱼类群落11个采样点的非度量多维标度分析Fig.3 Non-metric multi-dimensional scaling based on 11 sampling sites in the Liqiu River,a tributary of the Middle Yalong River
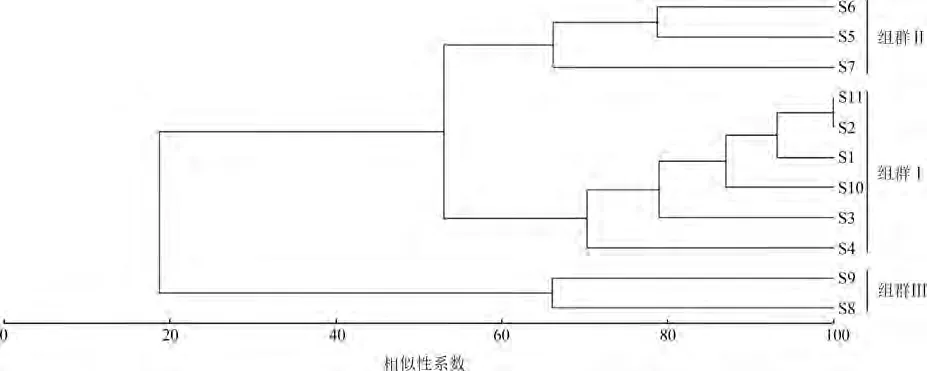
图4 雅砻江中游支流力丘河11个采样点的聚类分析Fig.4 Hierarchical cluster analysis based on 11 sampling sites in the Liqiu River,a tributary of the Middle Yalong River
SIMPER分析显示,组群Ⅰ、Ⅱ、Ⅲ组内的鱼类群落的相似性指数分别达到80.95%、70.38%和66.07%。组群Ⅰ采样点数多、空间跨度广,组内鱼类群落具有最高的相似性系数,各采样点采集的样本几乎仅由西溪高原鳅和软刺裸裂尻鱼2种优势种类组成。组群Ⅰ与组群Ⅱ的平均差异性为47.03%,对差异贡献较大的鱼类有短须裂腹鱼、齐口裂腹鱼和西溪高原鳅;组群Ⅱ与组群Ⅲ的平均差异性为58.87%,对差异贡献较大的鱼类有西溪高原鳅、齐口裂腹鱼和软刺裸裂尻鱼;组群Ⅰ与组群Ⅲ的平均差异性为92.51%,对差异贡献较大的鱼类有西溪高原鳅、齐口裂腹鱼和软刺裸裂尻鱼。
3 讨论
3.1 力丘河鱼类资源现状
虽然本次的渔获量较大(共计2 557尾),但力丘河的鱼类资源并不丰富,主要表现在以下几方面:种类丰富度较低:相比于历史记载雅砻江中游分布的19种鱼类(吴江,吴明森,1986),力丘河的鱼类种类组成较为简单,仅9种,张春光等(2019)在力丘河沙德镇发现的新种花斑裂腹鱼在本次调查中未被采集到,可能是由于其数量较少;种类分布极不均匀:仅软刺裸裂尻鱼、西溪高原鳅、短须裂腹鱼和齐口裂腹鱼分布范围较广,个体数占比达98.7%;各河段鱼类的Margalef种类丰富度指数、Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数均较低,且多样性从河口至上游逐渐降低。
丰度/生物量比较曲线显示力丘河鱼类群落受中等程度干扰。Spearman秩相关性系数表明,鱼类种类数和Margalef种类丰富度指数和海拔呈显著负相关。海拔是限制长江上游鱼类分布的主要环境因素(Fu,2004;石睿杰等,2018)。在金沙江和雅砻江上游的高海拔区域,鱼类群落普遍具有低物种多样性和高地方性特有种占比的特点(Liu,2020),低水温、低溶氧等高海拔河流生境中严苛的环境条件限制着鱼类的生长和繁殖,仅有裂腹鱼类、高原鳅类等少数对高海拔生境适应能力较强的物种能够在其中生存(赵亚辉等,2008)。力丘河是典型的横断山区高原河流,鱼类多样性较低、资源较不丰富可能是较高海拔所致。
丰富的鱼类多样性是实施支流保护的必要前提(高婷等,2012),支流生境保护有效的案例多见于鱼类多样性较丰富的平原地区(李浩林等,2014;林俊强等,2014;何滔,2016)。高原地区鱼类种类组成较为单一、多样性较低的现象可能限制支流生境保护策略的有效性。
3.2 海拔对力丘河鱼类分布的影响
力丘河的鱼类群落分为3组具有明显差异的空间组群,海拔是塑造鱼类群落空间结构的主要环境因子,原因如下:
第一,虽然塔拉沟和色物绒沟的汇口分别位于力丘河的中游和下游,但这2条支流的鱼类却和上游的鱼类共同构成了组群Ⅰ,可能是由于这些鱼类所处生境的海拔相近(3 300 m以上)。此外,组群Ⅰ还具有最低的物种丰富度和最高的相似性系数,表明只有对高海拔适应能力最强的少数种类才能在这些采样点所覆盖的区域生存。
第二,海拔对力丘河鱼类群落的影响还表现在优势种类在不同海拔区域的分布方面。SIMPER分析显示,组群Ⅰ、组群Ⅱ和组群Ⅲ3个群落之间差异显著的原因主要是软刺裸裂尻鱼和裂腹鱼属鱼类的相对数量差异,其中软刺裸裂尻鱼主要分布于力丘河上游等高海拔河段,而裂腹鱼属的齐口裂腹鱼和短须裂腹鱼则主要分布于海拔相对较低的中下游河段。虽然裸裂尻鱼属和裂腹鱼属均隶属于裂腹鱼亚科Schizothoracinae,但2个属的鱼类对海拔的适应能力及范围存在一定的差异(He,2010;Qi,2012):无鳞无须的软刺裸裂尻鱼属于高度特化的裂腹鱼类,其适应海拔为1 500~5 000 m,而具细鳞、吻部有2对须的裂腹鱼属则属于裂腹鱼类中较原始的类群,其对高海拔生境的适应能力稍弱(1 250~2 500 m)(王金林等,2018)。这2类裂腹鱼类在力丘河的分布特点反映了其对海拔的不同适应能力。
第三,力丘河中下游海拔落差很可能对干流鱼类形成阻隔作用,从而使其难以上溯到力丘河中、上游栖息。在本研究中,鱼类多样性最高的组群Ⅲ仅分布于力丘河下游普沙绒乡罗让至河口,该河段的海拔与干流相似,鱼类可能来自干流并栖息在该河段,但无法进一步上溯至力丘河的中上游河段,从而与组群Ⅱ的鱼类群落组成形成显著差异。
可见,高海拔不仅限制了力丘河的鱼类多样性,还塑造了力丘河鱼类群落的空间结构,并可能阻碍了雅砻江中游干流的鱼类向力丘河中上游的迁移。
3.3 力丘河作为鱼类栖息地的保护价值
良好的栖息地是鱼类资源维持和发展的根本,在很多干流受水电开发影响较为强烈的流域,自然连通性较为完好的支流往往可以作为维持干流鱼类多样性的重要栖息地(Marques,2018):一方面,这些支流可以为干流鱼类提供产卵场和索饵场(Nunes,2015;Azevedo-Santos,2020)及一些珍稀鱼类的庇护场所;另一方面,在干流河段形成水库后,支流的流水生境将成为干流中急流型鱼类的栖息地(高婷等,2012)。为了缓解雅砻江流域水电开发和水生态保护之间的矛盾,基于对栖息地质量的评估,姜跃良等(2015)建议对包括力丘河在内的一些雅砻江支流进行栖息地保护。本研究从鱼类多样性及群落空间格局的角度进一步探讨力丘河栖息地的保护价值。
力丘河中软刺裸裂尻鱼、西溪高原鳅、短须裂腹鱼等长江上游特有鱼类的资源量较丰富,因此该支流具有一定的鱼类多样性保护价值。同时,较大的海拔落差导致雅砻江干流的许多鱼类仅分布于普沙绒乡罗让以下的力丘河下游河段,无法进一步上溯至力丘河的中上游河段,从而与力丘河中游下段的鱼类群落(组群Ⅱ)组成形成显著差异;而力丘河上游至中游上段甲根坝乡(采样点 S1~S4)和塔拉沟(S10)、色物绒沟(S11)等支流几乎仅分布有软刺裸裂尻鱼和西溪高原鳅(组群Ⅰ)。因此,力丘河作为雅砻江中游干流水电开发后的鱼类栖息地的保护价值将一定程度降低,尚不足以有效缓解水电建设对雅砻江干流鱼类资源的影响,建议将保护范围扩大至干流的部分河段。
西华师范大学的杨坤博士参加了部分野外调查工作,在此特别感谢!
猜你喜欢
疯狂英语·新读写(2024年4期)2024-06-10 18:38:05
节能与环保(2022年3期)2022-11-24 13:25:57
水泵技术(2022年2期)2022-06-16 07:08:40
水泵技术(2021年4期)2021-11-20 06:28:50
现代苏州(2021年3期)2021-09-10 07:12:13
江苏教育研究(2020年28期)2020-11-23 07:30:10
高原山地气象研究(2020年1期)2020-06-24 08:20:10
网络安全和信息化(2017年3期)2017-03-10 07:45:51
水利科技与经济(2016年11期)2016-04-22 01:10:14
大众考古(2014年6期)2014-06-26 08:31:40
