
库布齐沙漠植被恢复对风沙土壤碳通量与碳储量的影响①
2022-07-22 10:52包玉海李雨薇王成龙
土壤 2022年3期
王 博,包玉海*,刘 静,李雨薇,王成龙
库布齐沙漠植被恢复对风沙土壤碳通量与碳储量的影响①
王 博1,包玉海1*,刘 静2,李雨薇1,王成龙2
(1 内蒙古师范大学地理科学学院,呼和浩特 010022;2 内蒙古农业大学沙漠治理学院,呼和浩特 010018)
为阐明库布齐沙漠植被恢复过程中土壤碳通量的时空动态特征及主控因子,明确土壤有机碳含量和储量的变化趋势,本研究以流动沙地、半固定沙地、藻结皮固定沙地和地衣苔藓混合结皮固定沙地为研究对象,运用静态暗箱–气相色谱法对风沙土壤碳通量及水热因子进行观测,并对土壤有机碳含量和密度进行测定和计算。结果表明,生长季内风沙土壤碳通量变异较大,季节动态与土壤温度基本一致,且随植被恢复碳通量呈递增趋势:混合结皮固定沙地(210.28 mg/(m2·h))>藻结皮固定沙地(177.45 mg/(m2·h))>半固定沙地(117.34 mg/(m2·h))>流动沙地(65.61 mg/(m2·h));土壤碳通量与各层土壤温度均显著正相关,除流动沙地土壤碳通量与深层土壤含水量显著负相关外,其余样地碳通量均与表层土壤含水量显著负相关;风沙土壤有机碳含量和密度随植被恢复而递增:混合结皮固定沙地 (1.32 g/kg,0.94 kg/m2) > 藻结皮固定沙地 (1.03 g/kg,0.74 kg/m2) > 半固定沙地 (0.45 g/kg,0.36 kg/m2) > 流动沙地 (0.27 g/kg,0.24 kg/m2),且均具有明显的随土层加深有机碳含量和密度逐渐降低的趋势;荒漠土壤表面CO2年通量与0 ~ 10 cm和10 ~ 20 cm土层有机碳密度显著正相关(< 0.05)。综上表明,人工建植促进植被恢复和沙地固定,增加风沙土壤碳排放通量,显著提高风沙土壤碳贮存能力和潜力,改变荒漠生态系统碳循环格局。
人工固沙区;碳通量;土壤有机碳;植被恢复;水热因子
土壤是陆地生态系统中储量最大的碳库,其储量规模约为大气碳库的2倍,是植被碳库的3倍[1]。土壤碳通量是土壤表面CO2通量,是土壤向大气释放CO2的主要途径[2],而土壤有机碳是进入土壤的生物残体等有机物质的输入与以土壤微生物分解作用为主的有机物质的损失之间的平衡,是直接衡量和评价土壤碳贮存能力的指标[3]。因此,作为土壤碳库输出和输入的重要通道,土壤碳通量和有机碳含量的任何细微变化,都将直接改变土壤圈碳储量及大气圈CO2浓度,进而影响生态系统碳循环过程和全球碳平衡[4]。开展陆地生态系统土壤碳通量和储量的研究,尤其在全球气候变化背景下,探讨不同土地利用方式下土壤碳库的变化特征,可为生态治理工作如人工林建设、天然林保护、退耕还林还草、荒漠化防治等提供科学依据,明确其价值性,保障其合理性。
目前,针对森林、草地、湿地等自然生态系统及城市绿地、农田等人工生态系统土壤碳动态的研究已较多开展,且主要集中于土壤呼吸速率、生态化学计量特征、碳氮分配格局、碳矿化及周转特性等[5-8]。荒漠生态系统作为陆地生态系统的重要组成部分,具有植被单一、覆盖度低、侵蚀严重的特点。研究表明,荒漠区土壤碳储量占整个土壤圈碳储量的9.5%,在全球碳循环过程中占有十分重要的地位[9]。因此,研究荒漠土壤碳动态特征,特别是土壤表面碳通量及地下碳储量,对于准确评估荒漠区碳收支及制定科学合理化的管护、利用措施具有十分重要的意义。
我国对于干旱、半干旱区的沙化土地,及大沙漠边缘和绿洲延伸区,已开展了广泛的荒漠化治理工作。通过人工建植促进沙地固定、植被恢复、改善区域微生境,可形成群落自然演替的稳定生态系统[10]。在这一过程中,随着植被覆盖度的增加,植被–土壤的互馈作用会影响土壤理化性状、根系分布、微生物拓殖及土壤动物活动,也能够促进地表生物结皮发育并趋向成熟演替[11-12]。库布齐沙漠是全球唯一被整体治理的沙漠,已修复绿化面积达6 253 km2,基本实现了人进沙退。本研究以库布齐沙漠东段人工固沙区土壤为研究对象,以沙生植被恢复过程为样地划分依据,旨在揭示该过程中:①风沙土壤碳通量时空动态特征及其环境主控因子;②风沙土壤有机碳含量及密度的动态变化特征;③风沙土壤碳通量及碳储量的协同关系。
1 材料与方法
1.1 研究区概况
研究区位于内蒙古自治区鄂尔多斯市准格尔旗境内,属库布齐东段典型沙漠地貌类型。研究区为温带大陆性气候,具有明显的水热同期特征,春冬季节干旱多风,夏秋季节炎热且降水集中。年均气温6.1 ~ 7.1 ℃,年均降水量240 ~ 360 mm,年均蒸发量2 560 mm,年均日照时数3 138 h,年均无霜期130 ~ 140 d,年均风速3.3 m/s。研究区内土壤以风沙土为主,土壤颗粒组成为黏粉粒(<0.05 mm)占比2.61%,极细沙(0.05 ~ 0.1 mm)占比3.92%,沙粒(0.1 ~ 1 mm)占比92.94%,主要植物种为沙柳()、柠条()、杨柴()、油蒿()、猪毛菜()、沙竹()、沙米()等。
1.2 样地设置
在库布齐沙漠东段人工固沙区,依据植被恢复程度和地表生物结皮发育特征,划分样地类型,分别为:①流动沙地,为裸露流动沙地,地表基本无植被覆盖,仅生长少量一年生草本植物,风蚀强烈;②半固定沙地,经人工飞播20 a后地表逐渐生长先锋灌木油蒿及一年生草本,地表出现少量物理结皮附着;③藻结皮固定沙地,于半固定沙地网格状扦插沙柳枝条,形成生物活体沙障,提高地表植被盖度,促进沙地固定,植被恢复期为8 a,地表已形成黑色斑状藻结皮(叶绿素a含量为0.31 μg/g,伪枝藻素含量为0.28 μg/g);④混合结皮固定沙地,行带式扦插沙柳枝条后形成稳定的沙柳群落,自然稀疏后形成冠幅较大的团簇状“沙柳岛”,岛间空地生长大量油蒿,植被盖度很大,凋落物层较厚,地表已形成连片灰绿色的地衣苔藓混合结皮 (叶绿素a含量为1.74 μg/g,伪枝藻素含量为6.32 μg/g)。样地基本情况见表1。
1.3 气体样品测定
于2017年植物生长季(5—10月)进行土壤CO2气体样品采集。在每种样地内选择地势相对平坦的地块,分别布设3块2 m × 2 m气体样方,拔除样方内所有草本植物,并尽量保证生物土壤结皮的完整性。使用静态暗箱进行土壤CO2采集,静态暗箱由圆筒形顶箱(直径:320 mm,高:600 mm)和基座组成,顶箱顶壁安装风扇,搅拌箱内气体使混合均匀,基座嵌入气体样方土体内,嵌入深度为15 cm。每次采样时提前2 min将顶箱扣入基座,并向基座凹槽注水进行密封,防止采样过程中箱体内外发生气体交换。每月对4种样地分别采样2次,时间间隔15 d左右。每次采样时间均固定在9:00—12:00 am,以降低系统误差。自顶箱扣入基座开始计,分别在0、15、30 min各取气体样品3次,每次重复采集气体3袋,每袋气体50 ml。采样工具为医用带三通阀的注射器,气体存储于铝箔气袋。

表1 样地基本信息
气体样品带回实验室低温保存,使用气相色谱仪 (Agilent 4890D,USA) 测定气体样品中CO2浓度,确保7 d内完成测定。
1.4 土壤水热因子测定
研究区内气象资料由小型自动气象站记录(HOBO,USA)。土壤水热因子为动态测定,与气体采样同步进行,使用水分速测仪(TRIME PICO,Germany)分层测定各样地土壤温度和含水量,土层划分为0 ~ 10、10 ~ 20、20 ~ 40和40 ~ 60 cm。
1.5 土壤有机碳测定
于2017年8月进行样地土壤调查。在每种样地内随机挖取3个土壤剖面,确定土壤发生层次后,分别在剖面深0 ~ 10、10 ~ 20、20 ~ 40和40 ~ 60 cm处混合取样200 g,装入无纺布袋,带回实验室后剔除植物根系与砾石,自然风干后采用重铬酸钾–浓硫酸氧化外加热法测定有机碳含量。土壤容重采用环刀法测定,环刀体积100 cm3,每层重复测定3次。
1.6 数据处理
土壤碳通量的计算是通过气体浓度随时间的变化,计算单位面积土壤的气体交换量,通量计算公式如下:
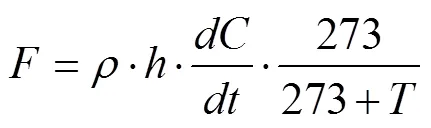
式中:为被测气体通量(CO2,mg/(m2·h));为标准状态下气体密度(kg/m3);为静态暗箱高度(m);/为箱内气体浓度变化斜率;为采样时箱内平均温度(℃)。
生长季土壤表面CO2年通量(C,g/(m2·a))采用累加法计算,即实测各月平均土壤碳通量通过以当月天数为步长乘积后累加计算。
土壤有机碳密度是指单位面积一定深度土层中土壤有机碳的储量,土壤剖面中第层的有机碳密度计算公式如下:

式中:SOC为第层土壤有机碳密度(kg/m2),C为第土层有机碳含量(g/kg),D为第土层容重(g/cm3),H为第土层厚度(cm),G为第土层砾石体积分数(%)。土壤剖面总有机碳密度为各土层有机碳密度累加得到。
试验数据处理及作图采用Excel及SigmaPlot 14.0软件进行,并采用SPSS 20.0软件进行统计分析。选取最小显著极差法(LSD)进行不同样地间土壤碳通量、水热因子、土壤有机碳密度等指标的差异显著性检验(=0.05),采用双因素方差分析、Pearson检验对变量进行相关性分析。图表中数据为平均值±标准误。
2 结果与分析
2.1 风沙土壤碳通量及水热因子变化特征
研究区中各样地间0 ~ 60 cm土壤温度无显著差异,生长季内平均土壤温度分别为25.32、24.42、25.19和25.11 ℃。风沙土壤温度的季节变化呈明显的单峰曲线(图1)。流动沙地和藻结皮固定沙地土壤最高温均出现在6月下旬,半固定沙地和混合结皮固定沙地最高温出现在7月下旬,各样地土壤最低温均出现在10月下旬。
流动沙地、半固定沙地、藻结皮固定沙地和混合结皮固定沙地间0 ~ 60 cm土壤含水量无显著差异,生长季内平均土壤含水量分别为81.6、73.2、74.6 和79.0 g/kg (表2)。风沙土壤含水量季节变化明显,各样地均为6、7月较低,5、10月较高,动态规律恰与土壤温度相反。

(A. 流动沙地;B. 半固定沙地;C. 藻结皮固定沙地;D. 混合结皮固定沙地)
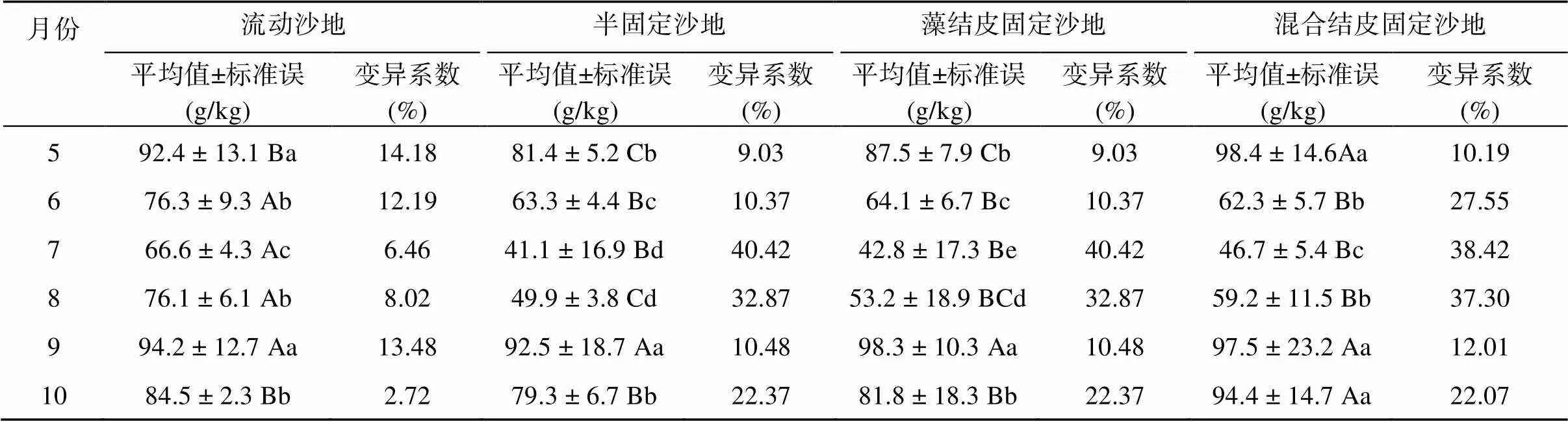
表2 不同植被恢复阶段0 ~ 60 cm平均土壤含水量变化
注:表中大写字母不同表示同一月份不同样地间差异达<0.05显著水平,小写字母不同表示同一样地各月份间差异达<0.05显著水平。
在生长季内,荒漠土壤碳通量时空变化明显,在植被恢复和生物结皮发育的不同阶段,及不同季节间土壤碳通量差异显著(<0.05),且动态规律与土壤温度基本一致,呈单峰曲线格局(图1)。流动沙地、半固定沙地、藻结皮固定沙地和混合结皮固定沙地土壤平均碳通量分别为65.61、117.34、177.45和210.28 mg/(m2·h),其中,流动沙地土壤碳通量最大值出现在7月上旬,半固定沙地为7月下旬,藻结皮固定沙地和混合结皮固定沙地均为6月上旬,各样地通量最低值均为10月下旬。
双因素方差分析可知(表3),植被恢复、季节变化及二者的交互作用对风沙土壤碳通量的影响均达到极显著水平(<0.01),且植被恢复对土壤碳通量的影响大于季节变化。对于土壤温度和含水量,仅季节变化能够产生极显著影响(<0.01),植被恢复和交互作用对二者无显著影响。表明植被恢复和生物结皮发育能够改变风沙土壤碳通量格局,但对土壤水热再分配影响不大。
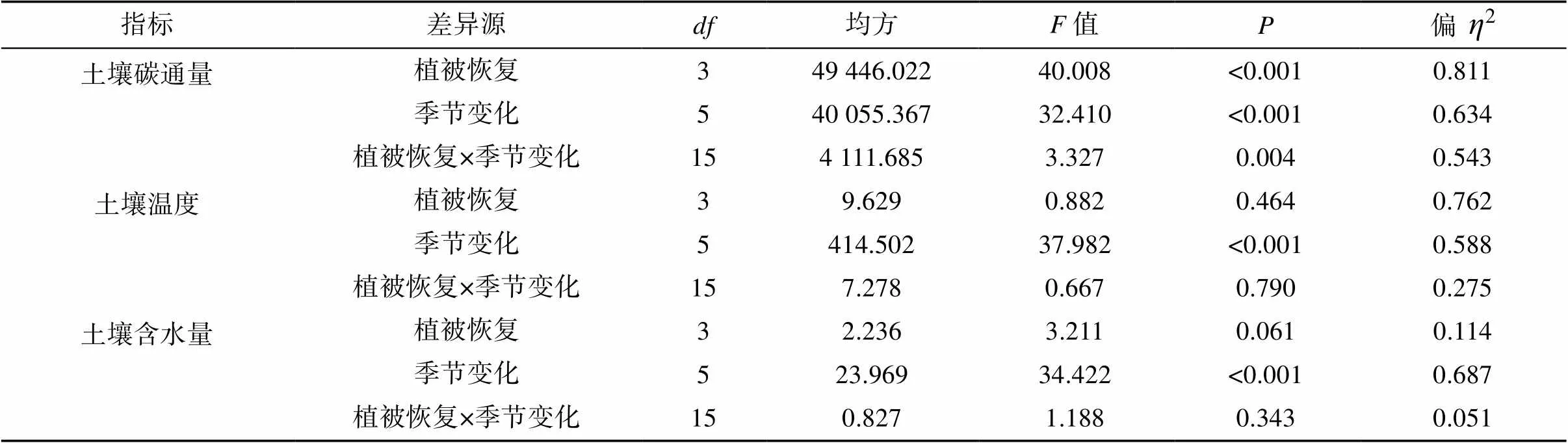
表3 植被恢复、季节变化及其交互作用对土壤碳通量、温度和含水量的影响
2.2 水热因子对土壤碳通量的影响
相关性分析表明(表4),土壤温度能够显著影响风沙土壤碳通量,各样地土壤碳通量均与各土层土壤温度显著正相关(<0.05)。土壤含水量对碳通量的影响因样地而异,流动沙地土壤碳通量与深层土壤含水量(20 ~ 60 cm)显著负相关,半固定沙地土壤碳通量与浅层土壤含水量(0 ~ 20 cm)负相关,而藻结皮固定沙地和混合结皮固定沙地土壤碳通量均与表层土壤含水量(0 ~ 10 cm)相关。表明土壤温度是影响风沙土壤碳通量的主控因子,植被恢复和生物结皮发育能够改变风沙土壤碳通量对水分条件的响应格局。
2.3 风沙土壤有机碳密度变化特征
植被恢复的不同阶段及土壤不同深度间,土壤有机碳含量和容重均具有显著差异性(<0.05)。在0 ~ 60 cm土层内,土壤有机碳含量随生物结皮演替逐渐增大,随土层加深而减小(图2)。流动沙地、半固定沙地、藻结皮固定沙地和混合结皮固定沙地土壤有机碳含量的变化范围是0.11 ~ 0.34、0.18 ~ 0.52、0.22 ~ 0.73和0.75 ~ 2.89 g/kg,其中,流动沙地土壤有机碳含量最大,分别是半固定沙地、藻结皮固定沙地和混合结皮固定沙地的5.11倍、2.77倍和1.14倍。在各土层中,0 ~ 10 cm土层土壤有机碳含量最大,分别是10 ~ 20、20 ~ 40和40 ~ 60 cm的3.67倍、2.44倍和2.07倍。土壤容重的空间变化恰与土壤有机碳相反,表现为随生物结皮演替逐渐减小,随土层加深而增大。
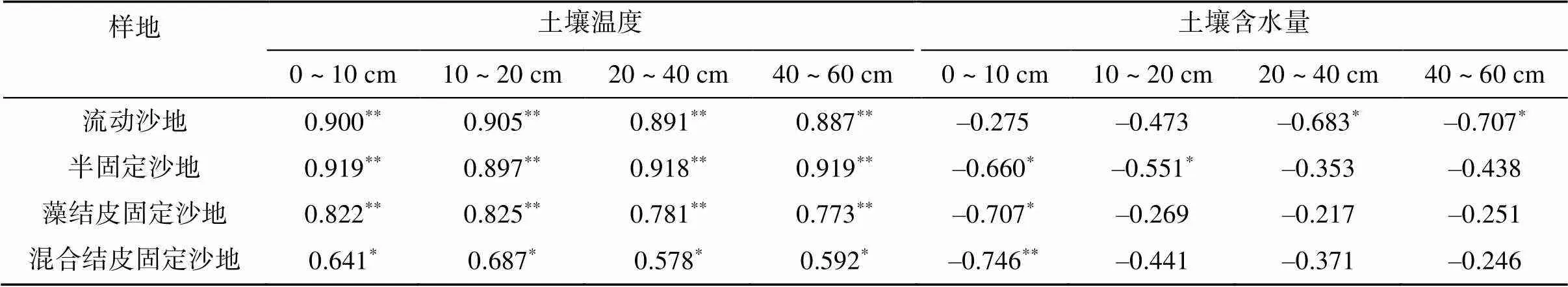
表4 风沙土壤碳通量与土壤水热因子相关系数
注:*、**分别表示相关性达<0.05和<0.01显著水平,下同。
不同植被恢复阶段风沙土壤总有机碳密度差异显著(<0.05),流动沙地、半固定沙地、藻结皮固定沙地和混合结皮固定沙地土壤总有机碳密度分别为0.243、0.362、0.736和0.940 kg/m2,具有随植被恢复而明显增加的趋势(表5)。
植被恢复、土层深度变化对风沙土壤有机碳含量、容重及土壤有机碳密度的影响均达到极显著水平(<0.01),而二者的交互作用仅对土壤有机碳含量产生显著影响(表6)。且对土壤有机碳密度而言,植被恢复所产生的影响大于土层深度变化,表明植被恢复和生物结皮发育能够显著改变风沙土壤碳储量,促进土壤碳贮存。
2.4 生长季内风沙土壤表面CO2通量与有机碳密度相关性
生长季内,风沙土壤表面CO2通量随植被恢复和生物结皮演替而逐渐增大,流动沙地、半固定沙地、藻结皮固定沙地和混合结皮固定沙变化能够产生极显著影响(<0.01),植被恢复和交互地土壤表面CO2年累计通量分别为269.28、567.19、759.02和911.40 g/(m2·a)。相关分析表明(表7),处于植被恢复过程中的风沙土壤,其表面CO2年通量与0 ~ 20 cm土层有机碳密度呈显著正相关(<0.05),与0 ~ 40 cm土层有机碳含量呈显著正相关(<0.05),与各土层容重呈显著负相关(<0.05)。
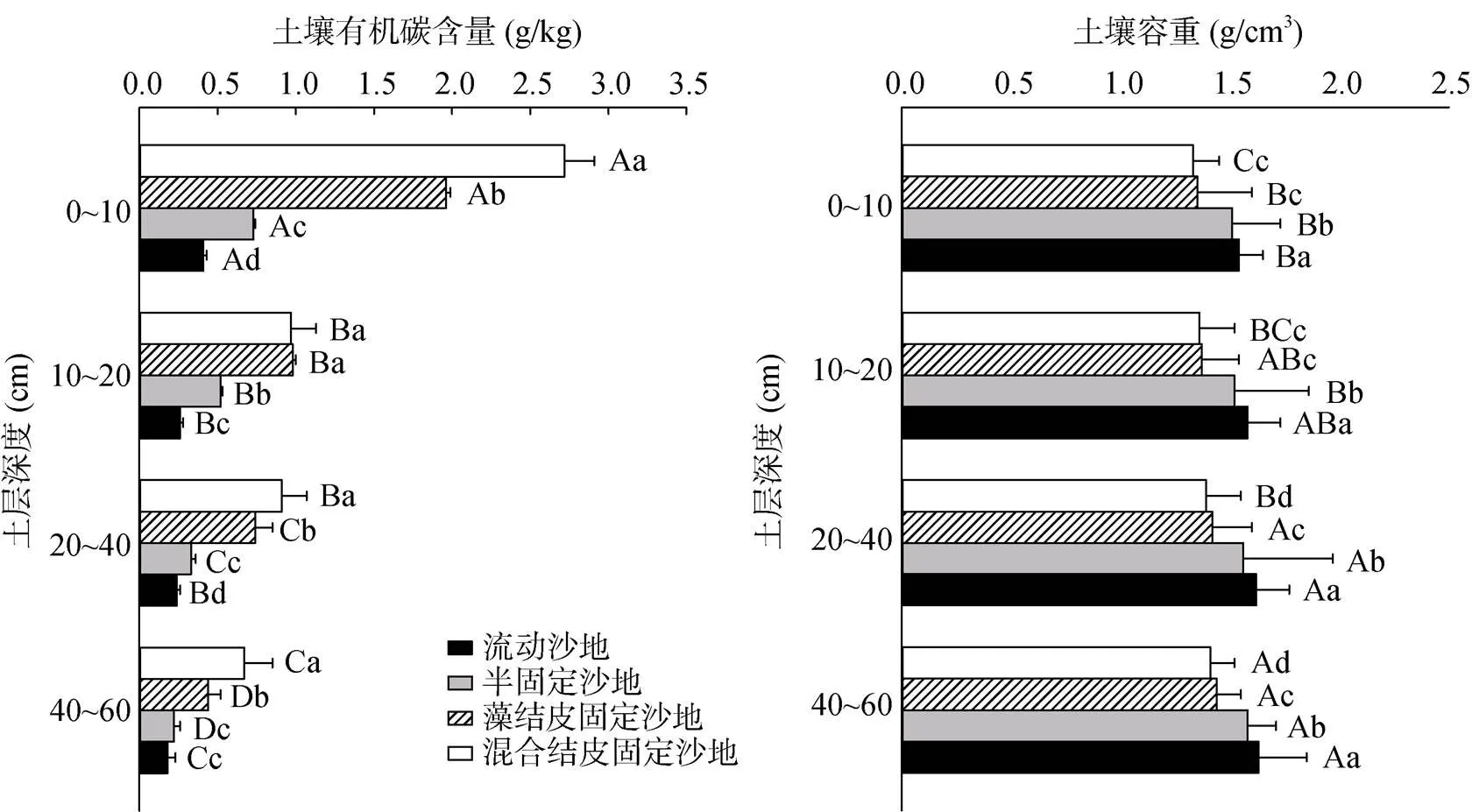
(图中大写字母不同表示同一样地不同土层间差异达P<0.05著水平,小写字母不同表示同一土层不同样地间差异达P<0.05著水平,下同)

表5 不同植被恢复阶段土壤有机碳密度变化

表6 植被恢复、土层深度及其交互作用对土壤容重、有机碳含量和有机碳密度的影响
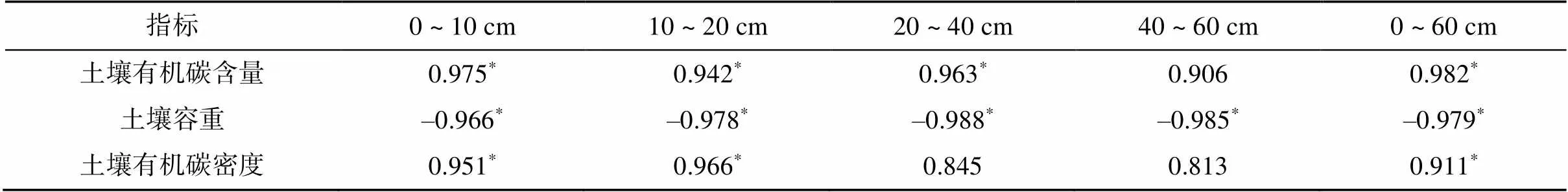
表7 风沙土壤表面CO2通量与土壤容重、有机碳含量和有机碳密度相关系数
3 讨论
3.1 风沙土壤碳通量季节变化及对水热因子的响应
本研究表明,在生长季内,从流动沙地到混合结皮固定沙地,风沙土壤碳通量表现出明显的季节变化,均呈现出与土壤温度相一致的单峰曲线变化,且相关性检验也表明二者之间存在显著的正相关关系,说明土壤温度是影响风沙土壤碳通量的主控因子,学者对西北干旱荒漠区的研究也得到相同结论[13]。土壤温度对土壤碳通量的直接影响,主要源于碳通量各组分对温度变化的敏感性。本研究所测得的土壤碳通量主要是由异养呼吸(土壤微生物呼吸)和自养呼吸(植物根系呼吸)组成。土壤温度能够改变土壤微生物的群落组成结构,还可改变微生物菌群数量,在一定范围内温度升高可以促进微生物增殖[14];也会显著影响微生物活性,随着温度升高,越来越多的分子达到或超过自身活化能,反应速度加快,碳排放量也随之提高[15]。植物现存根生物量对土壤温度变化的响应非常敏感[16],而活根可以进行自养呼吸,死根则是异养呼吸的基质,土壤温度升高增加根生物量累积,必然导致土壤碳通量增大。
干旱、半干旱荒漠区土壤含水量对土壤碳通量的影响较为复杂。本研究中,土壤含水量对土壤碳通量的影响相对较弱。在生长季内,仅流动沙地的深层和固定沙地的表层土壤含水量与土壤碳通量显著负相关。其原因可能是在水分匮乏的荒漠区,当土壤含水量较低时,植物冠层开始承受水分压力,分配到根系生长的可溶性碳水化合物比例增加,导致了较高的根系呼吸,增加土壤对CO2的排放量[17]。与本研究相反,学者对梭梭林的研究发现土壤碳通量与水分间存在显著正相关关系[18],而李永涛等[19]则表明二者间无相关性,这种复杂情况的出现,可能是因为只有当土壤含水量达到土壤生物(根系或微生物)萎蔫点或超过了田间持水量的情况下,土壤碳通量才会明显地受到土壤水分的影响,如果水分的变化没有超出上下限,并不足以影响土壤微生物或根系的生命力,则很难明显地测出水分对土壤碳通量的影响,此时土壤水分的影响很可能被其他因子的影响所掩盖[20]。
3.2 风沙土壤有机碳含量及密度对植被恢复的响应
本研究中,风沙土壤有机碳含量和密度在水平空间和垂直空间均呈现规律性变化。以植被正向恢复过程为水平轴线,土壤有机碳含量和密度呈现明显的递增趋势,从流动沙地到演替终点的混合结皮固定沙地,前者增长了4.89倍,后者增长了3.92倍,表明人工建植促进沙地固定可以有效增加风沙土壤碳贮存能力。一般来说,植被–土壤的协同作用、相互影响的反馈关系,是土壤性状改变的驱动力。随着地表植被种群和数量的增加,凋落物和根系生物量不断积累,有机残体大量分解周转返还于土壤,增加其有机碳含量[21]。同时,地表微生境在植被生长作用下的改变,为土壤微生物拓殖和生物结皮发育创造有利条件,前者自身的死亡分解和代谢分泌物,及后者通过隐花植物胶结形成的松、稳态腐殖质,也是土壤有机碳的直接来源[22]。而且,随着地表被草本、灌木及生物结皮的覆盖,可有效减弱风沙流活动,从而降低风蚀引起的土壤碳库流失。
以土层变化为垂直轴线,人工固沙区土壤有机碳随深度增加不断减小,表层含量(0 ~ 10 cm)为深层土壤(40 ~ 60 cm)的3.84倍,具有十分明显的养分表聚效应。这是因为大气降尘和有机残体输入都是发生在地表和浅层根际环境,土壤有机碳也主要累积于此,而研究区地处半干旱区,降水稀少,水分入渗困难,淋溶作用不强,导致外源输入的有机碳很难向深层土体移动,形成了养分贫富分明的垂直格局。
3.3 风沙土壤表面CO2通量与有机碳密度协同关系
本研究中,风沙土壤生长季内表面CO2年通量随植被恢复和生物结皮演替而显著增大,从流动沙地到混合结皮固定沙地,土壤表面CO2年通量增大了3.38倍,这与高艳红等[23]的试验分析一致。研究结果表明,土壤表面CO2年通量与0 ~ 10 cm和10 ~ 20 cm土层有机碳密度显著正相关,范跃新等[24]也发现土壤碳输入的增加是土壤碳排放随植被演替进程增大的重要原因。这主要是因为土壤底物浓度是决定土壤碳排放的重要因素,土壤有机碳是微生物分解和植物生长的主要碳源,其有效性和含量能够直接影响微生物活性和根细胞代谢,二者的异养和自养呼吸则是土壤碳排放的来源主体[25]。同时,土壤有机碳也会在土壤微生物或动物的参与下不断分解和转化为无机碳并释放CO2,而在植被恢复和生物结皮演替过程中,土壤有机碳不断积累,微生物菌群数量也不断增加,导致有机碳分解产生的碳排放量逐渐增大[26]。
4 结论
在半干旱荒漠区,进行人工建植固沙后,随着植被恢复,微生境发生改变,地表生物结皮逐渐形成并演替,土壤碳通量格局也发生明显变化。在生长季内,植被恢复和生物结皮演替可有效增加荒漠土壤碳排放通量,且碳通量具有明显的季节动态特征,其变化趋势与土壤温度基本一致。土壤温度是影响发生土壤碳通量的主控因子,土壤含水量对碳通量的影响较弱。在植被恢复和生物结皮演替过程中,风沙土壤有机碳含量和总有机碳密度均逐渐增大,且具有明显的表聚现象,风沙土壤表面CO2年通量同样呈现增长趋势,且与土壤有机碳密度呈显著正相关关系。植被恢复能够改变荒漠生态系统碳循环格局,是风沙土壤碳汇形成的主要驱动力。
[1] Raich J W, Potter C S. Global patterns of carbon dioxide emissions from soils[J]. Global Biogeochemical Cycles, 1995, 9(1): 23–36.
[2] Hamdi S, Chevallier T, Ben Aïssa N, et al. Short-term temperature dependence of heterotrophic soil respiration after one-month of pre-incubation at different temperatures[J]. Soil Biology and Biochemistry, 2011, 43(9): 1752–1758.
[3] Anderson O R. Soil respiration, climate change and the role of microbial communities[J]. Protist, 2011, 162(5): 679–690.
[4] 吕富成, 王小丹. 凋落物对土壤呼吸的贡献研究进展[J]. 土壤, 2017, 49(2): 225–231.
[5] 刘宝, 王民煌, 余再鹏, 等. 中亚热带天然林改造成人工林后土壤呼吸的变化特征[J]. 林业科学, 2019, 55(4): 1–12.
[6] Sharma C M, Gairola S, Baduni N P, et al. Variation in carbon stocks on different slope aspects in seven major forest types of temperate region of Garhwal Himalaya, India[J]. Journal of Biosciences, 2011, 36(4): 701–708.
[7] 卫玮, 党坤良. 秦岭南坡林地土壤有机碳密度空间分异特征[J]. 林业科学, 2019, 55(5): 11–19.
[8] Hu Y L, Li J T, Zhao S Y, et al. Soil respiration response to precipitation reduction in a grassland and a Mongolian pine plantation in semi-arid northeast China[J]. Journal of Forestry Research, 2019, 30(5): 1925–1934.
[9] Poulter B, Frank D, Ciais P, et al. Contribution of semi-arid ecosystems to interannual variability of the global carbon cycle[J]. Nature, 2014, 509(7502): 600–603.
[10] 周欣, 左小安, 赵学勇, 等. 半干旱沙地生境变化对植物地上生物量及其碳、氮储量的影响[J]. 草业学报, 2014, 23(6): 36–44.
[11] Cao C Y, Jiang D M, Teng X H, et al. Soil chemical and microbiological properties along a chronosequence ofLam. plantations in the Horqin sandy land of Northeast China[J]. Applied Soil Ecology, 2008, 40(1): 78–85.
[12] 张健, 刘国彬, 许明祥. 黄土丘陵区退耕地生物结皮影响下的土壤腐殖质分异特征[J]. 土壤, 2010, 42(4): 541–547.
[13] Song W M, Chen S P, Wu B, et al. Vegetation cover and rain timing co-regulate the responses of soil CO2efflux to rain increase in an arid desert ecosystem[J]. Soil Biology and Biochemistry, 2012, 49: 114–123.
[14] Biasi C, Rusalimova O, Meyer H, et al. Temperature-dependent shift from labile to recalcitrant carbon sources of Arctic heterotrophs[J]. Rapid Communications in Mass Spectrometry: RCM, 2005, 19(11): 1401–1408.
[15] Vose J M, Ryan M G. Seasonal respiration of foliage, fine roots, and woody tissues in relation to growth, tissue N, and photosynthesis[J]. Global Change Biology, 2002, 8(2): 182–193.
[16] Boone R D, Nadelhoffer K J, Canary J D, et al. Roots exert a strong influence on the temperature sensitivityof soil respiration[J]. Nature, 1998, 396(6711): 570–572.
[17] Casals P, Romanyà J, Cortina J, et al. CO2efflux from a Mediterranean semi-arid forest soil. I. Seasonality and effects of stoniness. Biogeochemistry[J]. Biogeochemistry, 2000, 48(3): 261–281.
[18] 王新友, 马全林, 靳虎甲, 等. 石羊河下游人工梭梭林土壤呼吸变化特征及其与水热因子的关系[J]. 干旱区地理, 2019, 42(3): 570–580.
[19] 李永涛, 王霞, 王振猛, 等. 黄河三角洲不同林龄人工柽柳林生长季土壤微生物与土壤呼吸特征研究[J]. 中南林业科技大学学报, 2019, 39(2): 86–92.
[20] 陈全胜, 李凌浩, 韩兴国, 等. 水分对土壤呼吸的影响及机理[J]. 生态学报, 2003, 23(5): 972–978.
[21] von Lützow M, Kögel-Knabner I, Ekschmitt K, et al. SOM fractionation methods: Relevance to functional pools and to stabilization mechanisms[J]. Soil Biology and Biochemistry, 2007, 39(9): 2183–2207.
[22] 闫德仁, 薛英英, 赵春光. 沙漠地区生物土壤结皮层腐殖质特征[J]. 生态学杂志, 2007, 26(12): 2017–2020.
[23] 高艳红, 刘立超, 贾荣亮, 等. 沙坡头人工植被演替过程的土壤呼吸特征[J]. 生态学报, 2012, 32(8): 2474–2482.
[24] 范跃新, 杨玉盛, 郭剑芬, 等. 中亚热带常绿阔叶林不同演替阶段土壤呼吸及其温度敏感性的变化[J]. 植物生态学报, 2014, 38(11): 1155–1165.
[25] 邓翠, 吕茂奎, 曾敏, 等. 红壤侵蚀区植被恢复对土壤呼吸及其温度敏感性的影响[J]. 土壤学报, 2019, 56(1): 135–145.
[26] Xu X, Zhou Y, Ruan H H, et al. Temperature sensitivity increases with soil organic carbon recalcitrance along an elevational gradient in the Wuyi Mountains, China[J]. Soil Biology and Biochemistry, 2010, 42(10): 1811–1815.
Effects of Vegetation Restoration on Sandy Soil Carbon Flux and Carbon Storage in Hobq
WANG Bo1, BAO Yuhai1*, LIU Jing2, LI Yuwei1, WANG Chenglong2
(1 College of Geographical Science, Inner Mongolia Normal University, Hohhot 010022, China; 2 College of Desert Control Science and Engineering, Inner Mongolia Agricultural University, Hohhot 010018, China)
In order to investigate the spatio-temporal dynamic characteristics and the main controlling factors of soil CO2flux during the process of vegetation restoration in Hobq desert, and clarify the variation trend of soil organic carbon content and density, the mobile dune, semi-fixed sand, algal crust fixed sand and mixed crust fixed sand were taken as the object, and soil CO2flux, temperature and moisture were continuously measured using the static closed chamber-gas chromatography technique, and the content and density of soil organic carbon were measured and calculated. The results showed that in the growing season, the spatial and temporal variation of soil CO2flux was obvious and increased with vegetation restoration, CO2flux was in an order of mixed crust fixed sand (210.28 mg/(m2·h)) > algal crust fixed sand (177.45 mg/(m2·h)) > semi-fixed sand (117.34 mg/(m2·h)) > mobile dune (65.61 mg/(m2·h)). Soil CO2flux of the four sites were significantly positively correlated with soil temperature. CO2flux of mobile dune was significantly negatively correlated with deep soil moisture, and were significantly negatively correlated with surface soil moisture in other sites. Soil organic carbon content density also increased with vegetation restoration: mixed crust fixed sand (1.32 g/kg, 0.94 kg/m2) > algal crust fixed sand (1.03 g/kg, 0.74 kg/m2) > semi-fixed sand (0.45 g/kg, 0.36 kg/m2) > mobile dune (0.27 g/kg, 0.24 kg/m2). Both the content and density of organic carbon decreased with soil depth. Soil surface carbon flux was significantly positively correlated with organic carbon density at the depths of 0–10 cm and 10–20 cm. The above results demonstrated that artificial planting could promote vegetation restoration and sand fixation, which effectively improved carbon emission flux of sandy soil, moreover, significantly increased carbon storage capacity and potential, thus could alter carbon cycle pattern of desert ecosystems.
Sand-binding area; Carbon flux; Soil organic carbon; Vegetation restoration; Hydro-thermal factors
S157.2
A
10.13758/j.cnki.tr.2022.03.014
王博, 包玉海, 刘静, 等. 库布齐沙漠植被恢复对风沙土壤碳通量与碳储量的影响. 土壤, 2022, 54(3): 539–546.
蒙古国基础地理要素与土地利用/土地覆被调查项目(2017FY101301-4)资助。
( baoyuhai@imau.edu.cn)
王博(1989—),男,内蒙古呼和浩特人,博士,副研究员,研究方向为生态修复与林业碳汇。E-mail: wbbrave@163.com
猜你喜欢
农业灾害研究(2022年1期)2022-05-07
南方农业·上旬(2022年3期)2022-04-15
当代体育(2021年23期)2021-09-10
环境(2021年5期)2021-06-20
幼儿智力世界(2021年4期)2021-05-17
土木建筑与环境工程(2018年4期)2018-09-29
科普童话·百科探秘(2018年7期)2018-09-13
大气科学学报(2018年4期)2018-09-10
上海师范大学学报·自然科学版(2018年4期)2018-05-14
江苏农业科学(2017年21期)2017-12-13
