
氨基酸态氮对油茶幼苗生长和吸收氮素的影响
2022-07-20 09:39:52李刘泽木王湘南何之龙陈隆升刘彩霞陈永忠李安亮
经济林研究 2022年2期
王 瑞,李刘泽木,张 震,唐 炜,王湘南,何之龙,陈隆升,刘彩霞,陈永忠,李安亮
(1. 湖南省林业科学院,湖南 长沙 410004;2. 国家油茶工程技术研究中心,湖南 长沙 410004)
油茶Camellia oleifera是原产于中国的木本食用油料树种,是世界四大木本食用油料树种之一,其主要产品茶油被联合国粮农组织推荐为“高级保健食用油”[1]。
氮是植物生长必不可少的营养元素之一,植物生长代谢过程中需要大量的氮[2]。土壤中氮的形态主要为无机氮和有机氮,氨基酸态氮是一种非常重要的有机氮,不仅为植物提供氮素养分,还参与能量代谢,对植物生长发育至关重要[3-11]。许多植物能直接吸收利用氨基酸态氮,但不同植物对氨基酸态氮的吸收能力不同,并且不同氨基酸对植物的效应也不同[10]。近年来,随着油茶产业的发展,有关油茶的科学研究取得了突出进展,关于油茶施肥方面有大量的研究报道[12-20],且多集中在无机氮方面。潘晓杰等[21]经研究发现,施用N、P、K、B、Zn 这5 种肥料对油茶幼林生长有明显的促进作用,且N、P、K 对油茶生长的作用由强到弱依次为N、P、K。张慧[22]经研究发现,每株施尿素390 g、钙镁磷肥390 g、氯化钾450 g并灌水29.5 g 的配比最有利于油茶产量的提升。陈铭等[23]经研究发现,不同形态氮素对油茶生长有显著的影响,其中铵态氮处理的油茶生长量、根冠比以及地下部分生物量达到最大,铵硝混合处理时总鲜质量达到最高。目前关于有机氮对油茶影响的研究报道不多。李安亮等[24]的研究结果表明,甘氨酸、赖氨酸和丙氨酸促进油茶嫁接苗高、地径和生物量增长的效果最好。在前期研究基础上,本试验中以1 年生油茶新品种幼苗为试材,选择了甘氨酸态氮、赖氨酸态氮和丙氨酸态氮3种有机氮,研究不同氨基酸态氮及硝态氮对油茶幼苗生长和氮素吸收的影响,以期为提高油茶苗木对氮肥的吸收利用效率提供参考。
1 材料与方法
1.1 试验材料
所用材料为油茶新品种‘朝霞’芽苗砧嫁接苗。容器规格为高12 cm、上口径8 cm。栽培基质为黄心土、珍珠岩和泥炭的混合物(体积比3∶1∶1),pH 为5.88,铵态氮质量浓度0.92 mg/L,硝态氮质量浓度2.34 mg/L。2014 年5 月嫁接后栽植于容器杯中,2015 年7 月选取生长一致且无病虫害的苗木进行施肥试验。
1.2 试验设计
有机氮选择甘氨酸态氮、赖氨酸态氮、丙氨酸态氮3 种氨基酸态氮肥,处理编号分别为Gly-N、Lys-N、Ala-N,无机氮选择硝酸钾(处理编号NO3-N),以无氮处理(CK)为对照。除CK 外,各处理的供氮浓度相同,均为3 mmol/L,使用霍兰格营养液供应其他必需大量元素和微量元素。自2015 年7 月,采用液体浇灌施肥的方式,每株苗每次施肥300 mL,每隔1 周施肥1 次,共8 次。每个处理30 株,3 次重复。试验在国家油茶工程技术研究中心温室中进行,温室光照度6 000 ~8 000 lx,温度20.0 ~25.0 ℃,湿度80%~85%。除施肥外,正常管护。
1.3 指标测定
1.3.1 生长量
当年11 月,每处理随机选取10 株,统计单株叶片数量,使用卷尺测量苗高,使用数显游标卡尺测量地径,3 次重复。同时,每株苗选取3 片叶,避开叶脉,使用SPAD 502 Plus 型叶绿素仪(柯尼卡公司,日本)测定SPAD 值,每片叶读取5 次数据,取平均值。
1.3.2 氮素吸收利用指标
当年11 月,每个处理随机选取10 株,每株采集3 片叶,用液氮带回实验室后,放超低温冰箱保存,待测。谷氨酸合酶(GOGAT)、谷氨酰胺合成酶(GS)活性测定参考Miflin 等[25]、杜旭华等[26-27]的方法,硝态氮和铵态氮含量的测定采用分光光度法[28-29]。
1.4 数据分析
使用Excel 2007 和SPSS 19.0 软件进行统计分析。采用单因素方差分析和LSD 比较处理间差异的显著性(P<0.05)。使用GraphPad Prism 8.0软件作图。
2 结果与分析
2.1 氮源对油茶幼苗生长的影响
不同氮源处理下油茶幼苗的生长情况见表1。由表1 可知,各氮源处理间油茶苗高均不存在显著差异,但均显著高于对照。与无氮处理(CK)相比,施用氨基酸态氮和硝态氮均促进了油茶幼苗的高生长,各处理按照苗高由高到低排序依次为赖氨酸态氮处理、甘氨酸态氮处理、丙氨酸态氮处理、硝态氮处理、无氮处理,说明氮素可以显著促进油茶苗的高生长,施用氨基酸态氮对油茶苗高生长的促进效果优于施用硝态氮,且各施用氨基酸态氮处理间不存在显著差异。在所有处理中:施用赖氨酸态氮处理的幼苗高生长最佳,苗高为13.72 cm,比对照提高了41.88%;其次为施用甘氨酸态氮处理,苗高为13.41 cm。
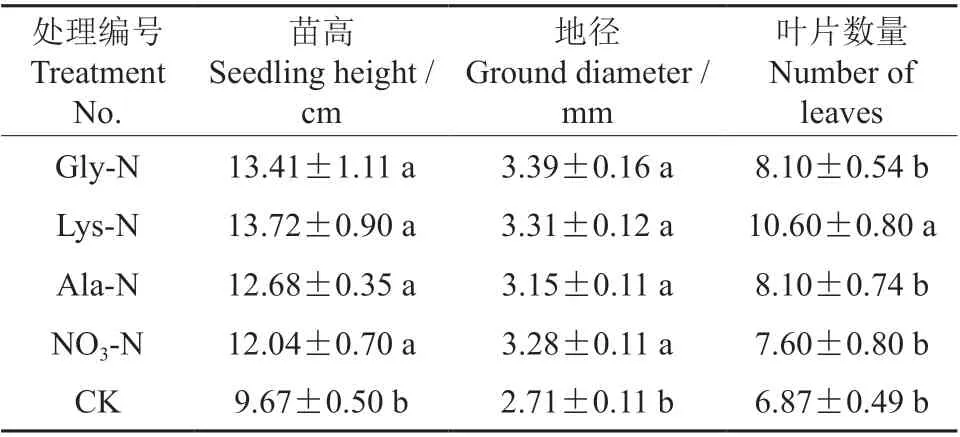
表1 不同氮源处理下油茶幼苗的生长情况†Table 1 The growth of C. oleifera seedlings treated by various nitrogen
在地径方面,各氮源处理间地径均不存在显著差异,均显著高于对照。与无氮处理(CK)相比,施用氨基酸态氮和硝态氮均促进了幼苗地径的生长,各处理按照地径由高到低排序依次为甘氨酸态氮处理、赖氨酸态氮处理、硝态氮处理、丙氨酸态氮处理、无氮处理,说明施用氨基酸态氮和硝态氮可以显著促进幼苗地径的生长。其中:施用甘氨酸态氮处理的苗木地径生长最佳,为3.39 mm,比对照提高了25.09%;其次为施用赖氨酸态氮处理,地径为3.31 cm。
在叶片数量方面,各氮源处理的叶片数量均高于对照(6.87),施用各种形态的氮素均促进了苗木叶片数量的增加,各处理按照叶片数量由高到低排序依次为赖氨酸态氮处理、甘氨酸态氮(丙氨酸态氮)处理、硝态氮处理、无氮处理。其中,施用赖氨酸态氮处理的叶片数量最高,为10.60,显著高于其余处理。说明施用氮素可以提高苗木的叶片数量,施用赖氨酸态氮可以显著促进苗木叶片数量的增加。
2.2 氮源对油茶幼苗叶片SPAD 值的影响
不同氮源处理下油茶幼苗叶片的SPAD 值如图1 所示。由图1 可知,施用氨基酸态氮和硝态氮均显著提高了叶片的SPAD 值,各处理按照SPAD 值由大到小排序依次为赖氨酸态氮处理、丙氨酸态氮处理、甘氨酸态氮处理、硝态氮处理、无氮处理。其中,施用赖氨酸态氮处理的叶片SPAD 值最高,为67.90,比对照提高了50.99%。Evans 等[30]的研究结果表明,植物叶片叶绿素含量与氮含量关系密切,SPAD 值与叶绿素含量呈正相关关系,可以利用叶片SPAD 值预测植物的氮营养状况。说明施用赖氨酸态氮可以显著提高叶片的叶绿素含量。

图1 不同氮源处理下油茶幼苗叶片的SPAD 值Fig. 1 The SPAD values in leaves of C. oleifera seedlings treated by various nitrogen
2.3 氮源对油茶幼苗叶片氮代谢酶活性的影响
不同氮源处理下油茶幼苗叶片的酶活性如图2 所示。由图2 可知,施用氨基酸态氮、硝态氮均显著提高了叶片中GS 的活性。在所有处理中:施用赖氨酸态氮处理的叶片GS 活性最高,为65.31 μmol/(g·h),显著高于其他处理,比对照提高了70.03%;施用硝态氮的处理次之,叶片GS 活性为55.15 μmol/(g·h),显著高于对照;无氮处理的叶片GS 活性最低,为38.41 μmol/(g·h)。说明施用氨基酸态氮和硝态氮均可以加速氮代谢途径的运转,促进氨基酸的合成和转化,以施用赖氨酸处理为最佳。由图2 可知,各处理的叶片GOGAT活性间均不存在显著差异。其中:CK 的GOGAT活性最高,为183.97 μmol/(g·h);施用甘氨酸态氮处理的GOGAT 活性最低,为147.62 μmol/(g·h)。

图2 不同氮源处理下油茶幼苗叶片的酶活性Fig. 2 The enzyme activity in leaves of C. oleifera seedlings treated by various nitrogen
2.4 氮源对油茶幼苗叶片氮素累积的影响
不同氮源处理下油茶幼苗叶片中氮素的含量如图3 所示。由图3 可知,所有处理间叶片中硝态氮含量均不存在显著差异。其中:施用硝态氮处理的叶片中硝态氮质量分数最高(3.24 mg/g),比对照提高了16.55%,比施用氨基酸态氮处理提高了3.09%~19.56%;施用氨基酸态氮处理的叶片中硝态氮含量均低于施用硝态氮处理,说明施用氨基酸态氮可以抑制叶片对硝酸盐的吸收;施用赖氨酸态氮处理的叶片中硝态氮含量最低,低于施用硝态氮处理及对照,说明赖氨酸态氮抑制吸收硝态氮的作用最明显。由图3 可知,施用氨基酸态氮和硝态氮均可以显著提高叶片中铵态氮含量,其中施用丙氨酸态氮处理的油茶叶片中铵态氮质量分数为2.27 mg/g,显著高于其余处理,比对照(0.51 mg/g)提高了3.45 倍。
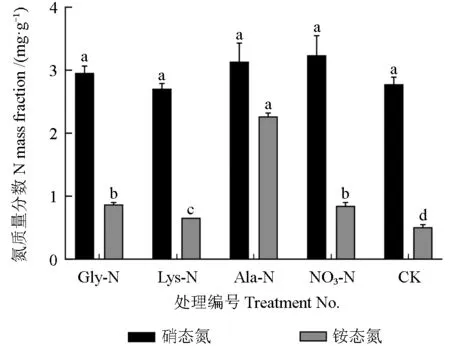
图3 不用氮源处理下油茶幼苗叶片中氮素的含量Fig. 3 The nitrogen content in leaves of C. oleifera seedlings treated by various nitrogen
3 结论与讨论
本研究结果表明,氨基酸态氮和硝态氮均可以促进油茶幼苗的生长及其对氮素的吸收,施用赖氨酸态氮可以显著提高油茶苗的高、地径、叶片数量、叶片SPAD 值和GS 活性,丙氨酸态氮比其他氮源更有利于提高油茶幼苗叶片铵态氮含量,氨基酸态氮对油茶幼苗吸收硝酸盐有抑制作用。在油茶苗木繁育过程中,可以用含有赖氨酸的有机肥部分替代化肥,提高氮素利用率,减少化肥的使用。
植物对氨基酸态氮的吸收利用具有一定的普遍性。有研究结果表明,氨基酸以分子形式被吸收,其供给植物氮素养分的作用较小[31]。氨基酸可能为生长调控因子或者信号传导物质,植物吸收氨基酸与氨基酸的分子特征以及植物细胞膜特异性有关[32-33],因而对于不同的氨基酸,植物吸收利用的能力也是有差异的。许玉兰等[34]的研究结果表明,施用甘氨酸使稻苗质量增加了23.7%,甘氨酸与亮氨酸混施处理中稻苗质量增加最多,增幅可达41.2%。吴良欢等[35]的研究结果显示,等氮量条件下,甘氨酸单施或甘氨酸与铵态氮配施处理中水稻的干质量均显著大于单施铵态氮处理。Sarwar 等[36]经研究发现,一定浓度的L-色氨酸对玉米株高、节间距和茎秆的鲜质量有显著影响。本研究结果表明,赖氨酸态氮可以显著促进油茶苗木高、地径、叶片数量的增加,说明赖氨酸态氮有利于促进油茶苗木的营养生长,这与李安亮等[24]的研究结果一致。
对玉米、小麦、水稻等作物的研究结果表明,叶片SPAD 值与氮含量间有较好的相关性[37-41]。本研究结果表明,不同氮源处理均显著提高了油茶叶片的SPAD 值,以赖氨酸态氮处理效果最佳,说明施用赖氨酸态氮可以显著提高叶片的叶绿素含量,这与李安亮等[24]得出的赖氨酸处理油茶叶片中光合色素含量最高的研究结果是一致的。
氨基酸可抑制植物对硝酸盐的吸收,使得植物体内硝酸盐含量降低。Muller 等[42]研究了施用氨基酸和硝酸盐对大豆吸收硝酸盐的抑制作用,结果表明天冬氨酸、谷氨酸、天冬酰胺、精氨酸、丙氨酸和β-丙氨酸对硝酸盐吸收的抑制作用最强。Aslam 等[43]的研究结果表明:当施用NO3-浓度为0.1 mmol/L 时,天冬氨酸、谷氨酸、天冬酰胺以及谷氨酰胺均会抑制大麦根系对硝酸盐的吸收,并且抑制效果相似,均达50%~60%;当NO3-的浓度为10 mmol/L 时,氨基酸对硝酸盐的吸收基本无影响。本研究结果与前人的研究结果基本一致,所有施用氨基酸态氮处理的叶片中硝态氮含量均低于施用硝态氮(KNO3)处理,并且施用赖氨酸态氮处理的叶片中硝态氮含量低于对照,说明其抑制硝酸盐吸收的作用更明显。这可能是由氨基酸对硝酸盐吸收、硝酸还原酶等的影响以及植物对氨基酸的优先吸收导致的,氨基酸态氮也可能是通过抑制硝酸转运载体的活性来降低植物对硝酸盐的吸收利用或抑制氮素代谢酶的活性[44]。
植物仅能利用水溶性有机氮,所以无机氮只有被同化为有机氮才能被植物体吸收和利用,绝大部分氨同化是通过GS/GOGAT 循环来实现的,GS 可以加速无机氮的转化,GOGAT 反映氨同化的强度[45-46]。本研究结果表明:施用氨基酸态氮和硝态氮后,叶片GS 活性均高于对照,说明施用氨基酸态氮和硝态氮均可以加速氮代谢途径的运转,促进氨基酸的合成和转化,其中施用赖氨酸态氮的效果最佳;施用不同形态氮素处理的叶片中GOGAT 活性间均不存在显著差异。
本研究结果表明,在油茶苗木繁育过程中,用少量含赖氨酸的有机肥[47]部分替代化肥,在一定程度上可以提高氮素利用率,避免因化肥过度施用带来的环境问题。
氨基酸态氮被植物吸收利用后不只是提供氮素养分或合成蛋白质的前体物质,其在植物体内的代谢过程十分复杂。氨基酸在植物体内的作用主要有合成蛋白质、作为合成植物激素的中间物质、螯合养分及其他物质、增强植物的抗逆性、增强植物的抗病能力。但是所吸收氨基酸对植物的影响是由其自身的代谢能力、环境条件以及氨基酸种类等众多因素综合决定的[32]。不同种类的氨基酸态氮对植物的生长及生理的促进作用是不同的,本试验中仅研究了单一氨基酸态氮对油茶苗生长及氮素吸收的影响,混合氨基酸态氮的种类及浓度对油茶苗木的影响及油茶对氨基酸态氮的吸收和转运机制均有待进一步研究。
猜你喜欢
天津农学院学报(2023年5期)2023-12-25 10:34:28
上海计量测试(2022年4期)2022-02-01 07:41:18
广东饲料(2016年3期)2016-12-01 03:43:11
水科学与工程技术(2016年2期)2016-07-10 10:19:57
人间(2015年11期)2016-01-09 13:12:58
动物营养学报(2015年10期)2015-12-01 03:30:54
邵阳学院学报(自然科学版)(2015年2期)2015-06-05 12:22:39
中国洗涤用品工业(2015年7期)2015-02-28 19:02:41
应用化工(2014年10期)2014-08-16 13:11:29
中国酿造(2014年9期)2014-03-11 20:21:03
