油茶种间杂交F1代18个优良单株核型分析*
2022-07-20 09:55:32李艳民袁德义叶天文韩春霞肖诗鑫
林业科学 2022年4期
李艳民 袁德义 叶天文 陈 雅 韩春霞 肖诗鑫
(经济林培育与保护教育部重点实验室 经济林育种与栽培国家林业和草原局重点实验室 中南林业科技大学 长沙 410004)
油茶(Camelliaoleifera)为山茶科(Theaceae)山茶属(Camellia)植物,是我国南方重要的木本食用油料树种(庄瑞林, 2008)。近年来,油茶产业备受重视,但是油茶生产上普遍存在产量不高、大小年明显的问题,严重影响了农民生产的积极性。究其原因是目前我国油茶良种主要为普通油茶物种,基本都是从野生普通油茶或地方品种中选优而来,丰产稳产且适应性强的品种较少。长期的选择育种会大大缩小物种的基因库,降低物种的遗传变异,增加物种对环境胁迫的敏感性(Klaudiaetal., 2018),已经成为油茶育种工作中迫切需要解决的难题。因此,在油茶育种中急需通过杂交,尤其是种间杂交来引入优良的遗传信息,丰富油茶的遗传变异。
杂交育种是油茶遗传改良的一项重要措施,然而一直以来育种工作进展缓慢,主要原因是目前我国油茶杂交育种研究仍是集中在种内不同性状互补间的杂交(林萍等, 2017),种间杂交相关研究相对较少(何华, 2016)。虽然种内杂交在育种中容易实现,但变异幅度通常有限。而种间杂交能极大程度地扩展和丰富育种筛选的基因库,有力地促进各种异源优良基因的交流,因而能够融合出具有各种优良性状的新变异类型,创造出良种选育所需要的新种质(Masonetal., 2015; Kartaletal., 2020)。因此,开展普通油茶与其近缘种的种间杂交将有力推动油茶品种改良进程。然而,由于对油茶细胞遗传学研究不足,对油茶杂交过程中的细胞遗传学规律也缺乏研究,加上油茶倍性复杂,给油茶种间杂交育种工作带来了极大的困难。
核型分析是指对染色体条数和形态结构以及带型等参数进行的分析,不但可以帮助我们从细胞学的角度了解植物的遗传组成和变异规律,而且还可以为真假杂种的辨别、倍性育种的成果鉴定、物种起源与演化的探讨、以及不同物种间亲缘关系的分析提供十分重要的参考价值,为植物分类以及种质创新提供一定的细胞遗传学依据(徐顺超等, 2012)。目前,核型分析已被广泛应用于小麦(Triticumaestivum)(Orlovskayaetal., 2020)、棉属(Gossypium)(Yescas-Romoetal., 2021)、樱桃(Cerasuspseudocerasus)(Wangetal., 2018)和山核桃(Caryacathayensis)(徐川梅等, 2017)等植物的育种研究中。自20世纪80年代掀起的山茶属植物核型研究热潮,取得了良好的进展(孙静贤等, 2019; 叶天文等, 2020),然而,国内外关于油茶种间杂交的核型研究却鲜有报道。攸县油茶(Camelliayuhsienensis)具有春季开花、抗炭疽病、果皮薄、油具芳香及富含维生素E等优异特性,是油茶育种中非常珍贵的遗传资源(庄瑞林, 2008; Nieetal., 2020),是油茶远缘杂交育种中是春花、薄皮和抗炭疽病基因的主要来源。前期本课题组创制了一批攸县油茶和普通油茶的杂交后代,经研究发现,杂交子代存在显著的性状分离和倍性变异,产生了具有超亲优势的优株和非整倍体资源(陈雅, 2020; Lietal., 2021),但其在染色体核型上是否存在变异还尚未研究。为进一步探究油茶种间杂交过程中的细胞遗传学规律,本研究通过对‘攸杂2’ × ‘华硕’杂交亲本及子代进行染色体核型分析和似近系数聚类分析,明确杂交子代的染色体核型变异和遗传组成,以期为探明油茶种间杂交过程中的染色体遗传规律和为后续杂交育种研究提供参考。
1 材料与方法
1.1 实验材料 ‘攸杂2’(YZ2)为攸县油茶后代优株,具有攸县油茶春季开花、皮薄、油香、抗炭疽病等优异特性,而普通油茶‘华硕’(HS)为国审大果、丰产普通油茶品种,但是皮厚且不抗炭疽病。本研究以‘攸杂2’ × ‘华硕’杂交后代群体为材料(18个优良单株分别命名为YH-1、YH-2、…、YH-18),染色体制片所采用的植物材料为幼嫩茎尖和扦插苗根尖。植物材料均采自中南林业科技大学生科楼楼顶,长势良好,位于112° 59′ 31″E,28° 8′ 11″N,海拔约80 m,全年平均气温17.2 ℃,年平均降水量1 361.6 mm。土壤为红壤。
1.2 试验方法 1) 染色体标本制备 于晴朗温和天气,取生长状态良好的扦插苗根尖,在冰水中清洗干净。再用0.002 mol·L-1的8-羟基喹啉在20 ℃ 环境中避光处理5 h,随后用蒸馏水冲洗干净再置于新配制的卡诺氏固定液(95%乙醇∶冰醋酸 = 3∶1, V/V)中于4 ℃ 冰箱中固定24小时。将固定后的根尖于0.075 mol·L-1的KCl溶液中处理60 min。随后用酶液(1.75%纤维素酶∶1.75%果胶酶 = 1∶1,V/V)于25 ℃ 恒温条件下黑暗处理2 h。将酶解后的根尖洗净后于蒸馏水中后低渗30 min。最后将根尖置于固定液中处理30 min备用。用滴管吸取3~4个根尖于载玻片上,在解剖镜下剔除分生区以外的部分后快速用宽头镊子捣成浆状,轻轻吹散并迅速在酒精灯上干燥。待玻片冷却后,吸取50 μL卡宝品红染液进行染色10 s后随即在OLYMPUS显微镜下镜检并拍照记录。
2) 染色体核型分析 每个样品选取5张染色体分散且形态清晰的中期分裂相,利用Image J和核型分析软件对图片进行处理和染色体测量、配对。根据李懋学等(1985)的标准进行核型分析。主要核型参数计算如下:
CLR = LC/SC。
CLR,染色体长度比;LC,最长染色体长度;SC,最短染色体长度。
AR = LA/SA。
AR,臂比;LA,长臂;SA,短臂。
CI = SA/TLC。
CI,着丝粒指数;SA,短臂;TLC,该染色体总长度。
As. K = TLLA / TCL × 100%。
As. K,核型不对称系数;TLLA,长臂总长;TCL,全组染色体总长。
3) 核型似近系数聚类分析 核型似进系数及进化距离依照谭远德等(1993)的方法,主要依据的核型参数为配子的染色体数、染色体相对长度、着丝粒指数、臂比值以及平均数、方差、极差等,采用李峰等(2005)设计的核型似近系数聚类分析软件计算亲本‘攸杂2’与‘华硕’及其种间杂交子代18个优良单株的核型似近系数及遗传距离并进行聚类分析。
2 结果与分析
2.1 亲本及杂交子代核型分析 亲本及杂交子代核型参数分析结果如表1所示,核型模式图如图1所示。

表1 亲本及杂交子代核型参数Tab.1 Karyotype parameters of parents and hybrids
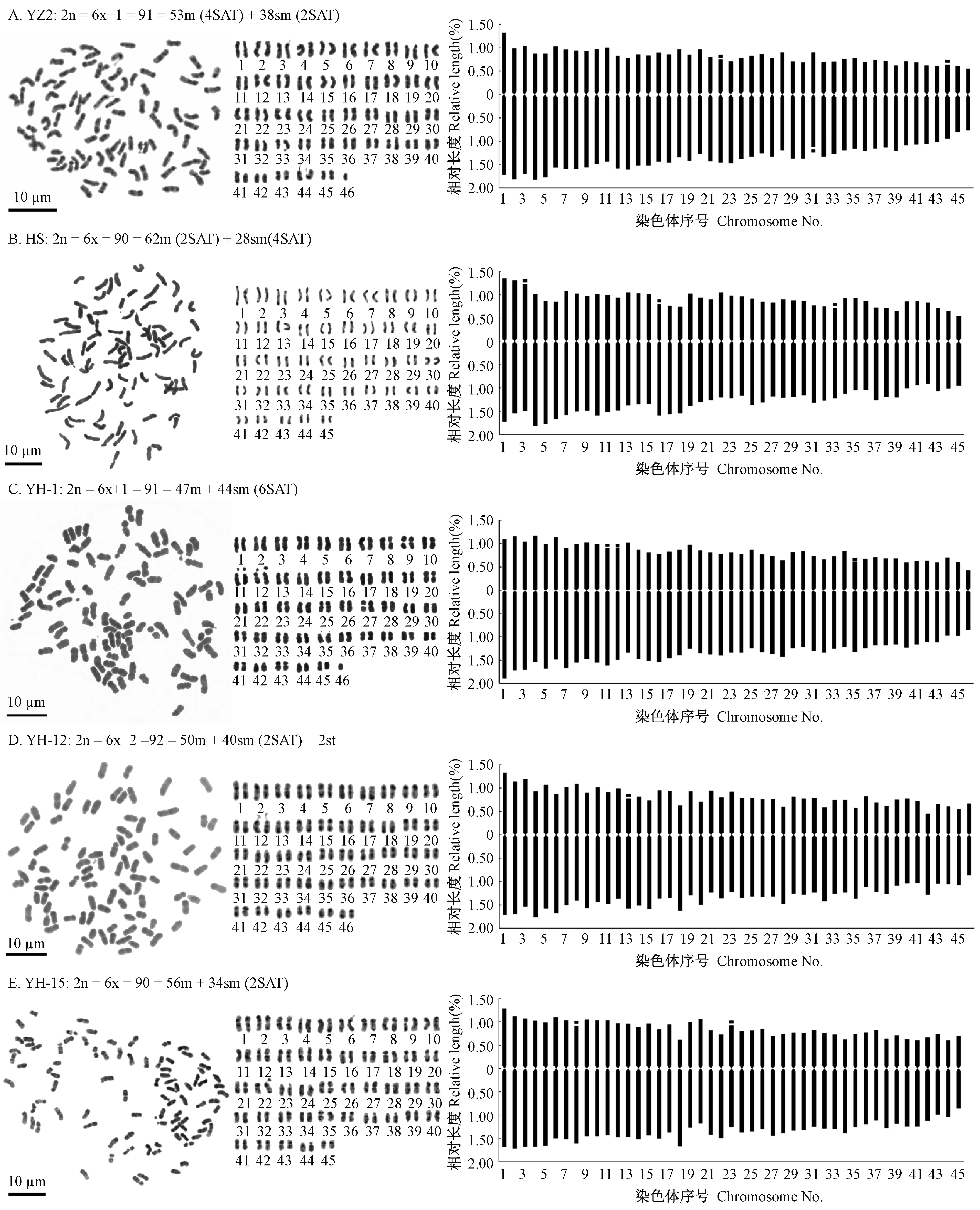
图1 亲本和杂交后代的染色体制片(左)、核型图(中)及核型模式图(右)Fig. 1 Photomicrographs of metaphase chromosome (left), karyogram (middle) and ideogram (right) of parents and hybrids
母本‘攸杂2’(YZ2)核型分析结果: ‘攸杂2’染色体条数为2n = 91,为非整倍体,核型公式为2n = 91 = 53m (4SAT) + 38sm (2SAT),由53条中部着丝粒(m)染色体和38条近中部着丝粒(sm)染色体,具有3对随体; 其染色体相对长度变化范围为1.26%~2.97%,其中有6条长染色体(L)、40条中长染色体(M2)、38条中短染色体(M1)和7条短染色体(S); 平均臂比值为1.67,其中臂比值大于2的染色体比例为17.8%,染色体长度比为2.36,核型不对称系数为62.36%,着丝粒指数的变异幅度为29.86%~44.04%,核型类型为2B型。
父本‘华硕’(HS)核型分析结果: ‘华硕’染色体条数为2n = 90,核型公式为2n = 90 = 62m (2SAT) + 28sm (4SAT),由62条中部着丝粒(m)染色体和28条近中部着丝粒(sm)染色体,具有3对随体; 其染色体相对长度变化范围为1.50%~3.00%,其中有6条长染色体(L)、40条中长染色体(M2)、42条中短染色体(M1)和2条短染色体(S); 平均臂比值为1.48,其中臂比值大于2的染色体比例为4.4%,染色体长度比为2.00,核型不对称系数为59.38%,着丝粒指数的变异幅度为32.14%~47.15%,核型类型为2B型。
18个杂交子代单株核型分析结果: 18个杂交子代单株中有11份染色体数目为90,6份染色体数目为91,1份染色体数目为92,杂交子代间染色体形态特征存在差异。研究发现其中绝大部分杂交子代的核型参数介于父本与母本之间。杂交子代的染色体类型仍是以m型和sm型染色体为主,其中YH-7和YH-12还含有少量st型染色体,均有随体; 大部分杂交子代中不同类型染色体条数所占的比例是m > sm > st,与父本‘华硕’和母本‘攸杂2’相同,但同时也出现sm型染色体数目大于m型染色体数目的特殊情况(YH-14、YH-18); 杂交子代的平均臂比值均高于父本‘华硕’,而更倾向与母本‘攸杂2’ ; 杂交子代的核型不对称系数变异范围为61.01%~63.38%,均高于父本‘华硕’,大多数介于父母本之间,但同时也出现部分杂交子代的核型不对称系数大于亲本的变异种质(YH-1、YH-13、YH-14、YH-17和YH-18); 核型类型为2A型的有6份,占33.3%,2B型的有12份,占66.7%。

表2 核型似近系数与进化距离①Tab.2 Similarity coefficient of karyotype and evolutionary distance

图2 核型似近系数和进化距离聚类图Fig. 2 Karyotype proximity coefficient and evolutionary distance clustering graph
2.2 核型似近系数及聚类分析 核型似近系数与进化距离分析结果能很好地反映试验材料的亲缘关系,本研究根据亲本及杂交子代的染色体数及相对长度等一系列核型参数为指标计算种间杂交群体核型似近系数及遗传距离,结果表明: 杂交子代当中YH-3和YH-4之间的核型似近系数最大(0.998 2)而核型进化距离最小(0.001 8),表明YH-3和YH-4之间的亲缘关系最近; 母本‘攸杂2’与杂交子代YH-3的核型似近系数最大为0.996,进化距离为0.004,与杂交子代YH-14的核型似近系数最小为0.961 6,进化距离为0.039 2,这表明在杂交子代中YH-3与母本‘华硕’的亲缘关系最近而YH-14与‘攸杂2’亲缘关系最远; 父本‘华硕’与杂交子代YH-11的核型似近系数最大为0.979 5,进化距离为0.020 7,与杂交子代YH-14的核型似近系数最小为0.938 8,进化距离为0.063 1,这表明在杂交子代当中YH-11与父本‘华硕’亲缘关系最近而YH-14与父本‘华硕’亲缘关系最远; 此外,母本‘华硕’与父本‘华硕’的核型似近系数较小,为0.962 8,进化距离为0.037 9,表明亲缘关系也较远。
采用李峰等设计的核型似近系数聚类分析软件对‘攸杂2’和‘华硕’及其杂交子代18个优良单株进行聚类分析并绘制聚类图(图2),结果显示: 在核型似近系数为0.966 0,距离系数为0.034 0时,可分为两大类: 父本‘华硕’与YH-11、YH-15聚为一类,母本‘攸杂2’与其余的所有杂交子代可聚为一类; 在核型似近系数为0.974 5,距离系数为0.025 5时,可分为三大类: 其中母本‘攸杂2’可与YH-1、YH-2、YH-3、YH-4、YH-5、YH-6、YH-7、YH-8、YH-9、YH-10、YH-12、YH-13、YH-16、YH-18归为一类,父本‘华硕’与YH-11、YH-15聚为一类,杂交子代YH-14和YH-17聚为一类。聚类结果表明大部分子代单株的核型与母本‘攸杂2’的核型更相似,亲缘关系更近; 其中,杂交子代YH-11和YH-15的核型相对而言与父本‘华硕’的核型更相似,亲缘关系更近; 此外,研究发现,当核型似近系数为0.974 5,距离系数为0.025 5时,杂交子代YH-14和YH-17在一定程度上与亲本的核型差异较大,亲缘关系均较远,可单独聚为一类。
3 讨论
染色体核型分析可以为研究物种间的亲缘及进化关系和遗传多样性提供一定的参考依据。本研究中,父本‘华硕’为普通油茶,核型类型为2B型,平均臂比值为1.48,核型公式为2n = 6x =90= 62m (2SAT) + 28sm (4SAT),这与前人报道的普通油茶核型(2n = 6x = 90 =60m + 22sm + 8st和2n = 6x = 90 = 60m + 29sm (3SAT) + 1st)均存在较大差异(李光涛, 2001),主要为本研究中未发现st型染色体以及存在3对随体,推测其可能是由于同一物种不同栽培品种的核型会存在一定的差异所导致。同时制片的质量也会对核型参数的测量产生较大的影响,为了避免因制片质量带来的误差,本研究采用改良后的去壁低渗法替代传统的压片法,获得的制片染色体分散,形态清晰,提高了核型分析结果的准确性。作为同一品种,本研究中‘华硕’的核型与叶天文等(2020)报道的核型(2n = 6x = 90 = 63m (3SAT) + 27sm)差异较小,证明了该方法在研究油茶染色体核型中具有较高的可靠性,同时,较小的差异也表明常规的染色体核型分析存在一定的局限性,由于油茶遗传背景相对复杂,许多非同源染色体形态相近,在研究核型时同源配对较为困难,对核型分析的准确性会产生一定的影响,需要观察更多的分裂相或结合分带及荧光原位杂交等技术以消除误差。此外,叶天文等(2020)在绘制‘华硕’核型图时,依照同源配对原则,将染色体分为15对,每对6条; 根据前人的研究(秦声远等, 2018)笔者认为普通油茶应为异源多倍体,且课题组前期研究发现,‘华硕’减数分裂终变期染色体配对主要为二价体(Lietal., 2021),因此,本研究认为应该将染色体分为45对,每对2条。但目前仍没有充分的证据证明普通油茶为异源六倍体,且其染色体组成与二倍体祖先种至今仍不清晰,需利用荧光原位杂交和分子标记等技术进行更深入的探究。
前人研究认为,种间杂交可能会导致高频的核型变异(Douetal., 2017)。本研究首次对油茶种间杂交群体的核型进行比较分析,其核型特征与前人对山茶属植物的核型研究结果相一致,即以m型染色体占大多数,其次为sm型,少数有st型,尚未发现t型(张文驹等, 1999)。但在杂交子代中出现个别单株的sm型染色体数目大于m型染色体数目(YH-14、YH-18)。而且,在种间杂交群体中,虽大多数杂交子代的核型参数介于亲本之间,但也有部分杂交子代的核型参数产生较大的变异,其原因可能是由于种间杂交过程中发生染色体重组或减数分裂过程中异常的染色体行为导致(陈琼等, 2007; Amosovaetal., 2017)。染色体融合和分裂重排被认为是影响植物核型结构的重要因素之一(Mandkovetal., 2013; Fonsecaetal., 2016; Suseketal., 2016),易位、倒位、重复以及缺失等染色体结构异常往往会引发染色体形态发生变化而导致核型变异,进而对物种的进化产生影响(Tzanoudakis, 1983)。
Stebbins(1971)指出高等植物核型进化的基本趋势是由对称向不对称方向发展。本研究中亲本和种间杂交群体的不对称系数变异幅度为59.38% ~ 63.38%,核型不对称系数较大,属于较为进化的类群。其中父本‘华硕’的核型不对称系数最小(59.38%),母本‘攸杂2’为62.38%,表明在山茶属植物系统发育中攸县油茶较普通油茶更为进化。杂交子代的核型不对称系数大多介于父母本之间,但也出现部分杂交子代的核型不对称系数大于亲本,说明其在进化程度上高于双亲,这与张素勤等(2018)对小麦与野燕麦(Avenafatua)远缘杂交群体的研究类似,表明种间杂交能够产生比亲本更为进化的子代,从而可以丰富油茶的遗传背景及种质资源。但是长期以来,由于山茶属倍性复杂多样、同源染色体间差异不明显、杂合度较高,导致仅从传统核型角度难以准确研究同一组内该属植物的系统进化(李光涛, 2001),还需结合荧光原位杂交和分子生物学等多种手段研究该属植物的系统演化。
近年来,核型似近系数分析越来越多地被应用于植物亲缘关系和系统演化等研究中(乔永刚等, 2020; 陈永毕等, 2019),但在山茶属植物中却鲜有研究。本研究针对‘攸杂2’和‘华硕’及其杂交子代群体进行核型似近系数聚类分析,发现大部分杂交子代的核型与母本更相近,而YH-11和YH-15的核型与父本更相近。同时,结合表型研究发现,大部分杂交子代在花期和果实经济性状上也存在倾向于母本的遗传趋势(陈雅, 2020),与核型分析结果一致,表明在油茶种间杂交过程中,母本的遗传信息能更好地传递给子代,而母本的性状可能有更高的遗传力。同时也有可能是因为母本为非整倍体(2 n+1),本身能传递更多的遗传物质给子代,且非整倍体植株可能会生成可遗传的表观遗传变化使得整倍体后代也能遗传非整倍体的表型特征(Henryetal., 2010)。此外,研究还发现,杂交子代YH-14和YH-17在一定程度上与亲本的亲缘关系均较远,这可能是由于种间杂交增加了基因重组频率,以及外源染色体片段的渗入,导致杂交子代在遗传组成上与双亲均产生一定的差异(渠云芳等, 2013),为种间杂交创制油茶新种质奠定了一定的理论依据。
4 结论
种间杂交群体中以m型和sm型染色体为主,少数有st型,均有随体。大部分杂交子代的核型参数介于父本与母本之间,少数杂交子代的核型出现较大变异。杂交子代YH-1、YH-2、YH-3、YH-4、YH-5、YH-6、YH-7、YH-8、YH-9、YH-10、YH-12、YH-13、YH-14、YH-16、YH-17和YH-18的核型与母本‘攸杂2’更相近,而杂交子代YH-11和YH-15的核型与父本‘华硕’更相近。
猜你喜欢
植物研究(2023年5期)2023-09-09 08:01:22
河南师范大学学报(自然科学版)(2022年5期)2022-08-08 14:07:42
宁夏医学杂志(2020年3期)2021-01-21 08:23:24
广西林业科学(2016年2期)2016-03-20 05:53:21
广西林业科学(2016年1期)2016-03-20 05:32:58
广西林业科学(2016年4期)2016-03-16 05:44:47
广西林业科学(2016年3期)2016-03-16 05:43:34
杂草学报(2015年2期)2016-01-04 14:57:55
哈尔滨医药(2015年2期)2015-12-01 03:57:21
中国当代医药(2015年9期)2015-03-01 02:02:12