
Lnc RNA TUC338 影响人类宫颈癌细胞的蛋白组学分析
2022-07-18 10:33蒋漪桦杜洪灵秦锦龙宋君雅蔡骁垚姝1
医学信息 2022年13期
蒋漪桦,朱 宇,杜洪灵,秦锦龙,刘 晔,宋君雅,蔡骁垚,金 姝1,,4
(1.南华大学衡阳医学院,湖南 衡阳 421001;2.上海市普陀区人民医院妇产科,上海 200060;3.上海市普陀区人民医院检验科,上海 200060;4.上海市普陀区人民医院中心实验室,上海 200060)
长链非编码RNA(Lnc RNA)是一类读码框之外的RNA,近年来Lnc RNA 被发现存在多种生物功能,在癌症、免疫、神经、心血管等多种系统中存在重要作用,尤其在癌症领域的研究成为热点。在肿瘤的诊断、药物治疗的靶点、判断肿瘤预后等方面均取得进展。Lnc RNA TUC 338 长度为590 bp,位于人类第12 对染色体长臂上。对于Lnc RNA TUC 338 的研究较少,已发现在肺癌[1]、膀胱癌[2]和食管癌[3]中,Lnc RNA TUC338 均通过不同机制促进癌症发展,提示其为一种癌基因。而宫颈癌作为女性生殖系统的三大肿瘤之一,严重影响女性的生理、心理健康[4],一定程度上也影响了两性关系的和谐[5]。Lnc RNA TUC 338 在人类宫颈癌中的相关研究较少,本研究探索了Lnc RNA TUC 338 在宫颈癌的发展中的作用及其影响宫颈癌细胞的发展的机制,现报道如下。
1 材料与方法
1.1 材料 人类宫颈癌细胞Hela 细胞购于中国科学院细胞库。胎牛血清DMEM 培养基、0.25%胰酶购于赛默飞世尔生物化学制品有限公司(北京);磷酸盐缓冲液(PBS),购于索莱宝科技有限公司(北京);DNA 分子量标准Ladder H1(100-1000 bp),购于生工生物工程股份有限公司(上海);RNAfast 200 总RNA 提取试剂盒,购于飞捷生物技术有限公司(上海);FastKing RT Kit 试剂盒、FastFire qPCR PreMix试剂盒购于天根生化科技有限公司(北京);引物合成:生工生物工程股份有限公司(上海);病毒包装:维诺赛生物技术有限公司(武汉)。
1.2 仪器 离心机:Eppendorf 离心机Centrifuge 5418;显微镜:重庆奥特光学仪器有限公司;荧光显微镜:Nikon ECLIPSE Ti 荧光显微镜;PCR 仪:BIORAD My Cycler;酶标仪:BioTek Synergy H1 全功能酶标仪;电泳仪:Tanon EPS 300 电泳仪;电泳设:Bio-Rad SDS-PAGE 电泳设备;凝胶成像分析系统:Tanon 1600。
1.3 方法
1.3.1 细胞培养与转染 将HELA 细胞以每孔1×106个置于6 孔板中,每孔加入浓度10%FBS 的DMEM 2 ml,5%CO2、37 ℃培养箱中培养,取对数生长期的细胞,每48 h 进行细胞传代,待细胞融合度达60%~70%时,按慢病毒使用说明书以脂质体法分别转染慢病毒,将HELA 细胞分为两组:转染rLVhTUC338-ZsGreen-Puro 慢病毒(实验组,以下简称TUC338 组)、转染rLV-ZsGreen-Puro 对照组病毒(对照组,以下简称rLV 组),转染48 h 后以2 μg/ml的比例加入嘌呤霉素杀灭未转染细胞,48 h 后再杀灭1 次,培养至细胞融合度达80%后进行以下各种实验。
1.3.2 RT-PCR 实验验证Lnc RNA TUC338 扩增 待细胞基本铺满板底时按照RNAfast 200 总RNA 提取试剂盒的说明书提取细胞RNA,按照天根FastKing RT Kit(with gDNase)说明书将RNA 逆转录为cDNA,按照天根FastFire qPCR PreMix(SYBR Green)试剂盒说明书,将cDNA 目的基因进行扩增,条件为:94 ℃3 min,94 ℃30 s、52 ℃30 s、72 ℃30 s循环39 次,72 ℃10 min 退火。TUC338 引物:Forward:TCAACTTAATCCCACACTGACC,Reverse:GAGCCTTGGAGACTGAACATC。以β-actin 为内参照,β -actin 引 物:Forward:CTCGCCTTTGCCGATCC,Reverse:TCTCCATGTCGTCCCAGTTG。将扩增的目的基因分别进行蛋白电泳,以1000 bpMarker为参照。
1.3.3 细胞送样及蛋白测序 将每组细胞样本分为3 个组别,送至北京诺禾致源技术公司,通过非标记定量方法(Label-free)对蛋白质酶解肽段进行质谱分析。
1.3.4 蛋白质谱分析 通过非标记定量方法(Labelfree)对2 组细胞的蛋白质酶解肽段进行定量、差异和富集分析。
1.3.5 TUC338 对蛋白基因本体功能(GO)的影响分析 分析基因本体功能(GeneOntology,GO)(www.geneontology.org),包括细胞组分(CC)、分子功能(MF)、生物过程(BP)。
1.3.6 KEGG 分析 通过Pathway 的主要公共数据库(http://www.genome.jp/kegg/)分析出已确定蛋白质参与的重要信号转导途径和生化代谢途径。
1.3.7 蛋白结构域分析 利用Interproscan 软件对蛋白质结构域分析,数据库包括Pfam、ProDom、SMART 等。
1.3.8 亚细胞定位分析 采用Cell-mPLOC 2.0 网站对测出的蛋白进行亚细胞的定位,亚细胞包括细胞核、线粒体、内质网、高尔基体、细胞膜和胞浆等。
1.3.9 蛋白互作分析 对鉴定到的蛋白进行蛋白互作分析,应用数据库http://string-db.org/(StringDB 蛋白互作数据库)。
1.4 统计学分析 使用SPSS 22.0 软件对所得到的实验结果进行统计学分析,采用t检验比较组间差异,P<0.05 有统计学意义。
2 结果
2.1 过表达TUC338 稳定细胞系和总蛋白鉴定 已筛选稳定的细胞系:荧光显微镜下见两组细胞荧光显色均为100%,慢病毒感染成功,见图1A;Lnc RNA TUC338 在实验组中的表达:经RT-PCR 实验,可见相较于对照组,实验组中的Lnc RNA TUC338 基因显著过表达,见图1B;总蛋白鉴定:对提取到的蛋白进行12%的聚丙烯酰胺凝胶电泳(SDS-PAGE),并进行考马斯亮蓝染色,两组均显示较多的蛋白条带,见图1C,提取蛋白成功。

图1 过表达TUC338 稳定细胞系和总蛋白鉴定
2.2 蛋白质谱分析
2.2.1 差异蛋白分析 蛋白定量分析共检测到4323个蛋白。蛋白差异分析显示:与rLV 组比较,在TUC338 组中下调的蛋白有194 个,上调的蛋白有225 个。以火山图(图2A)和热图(图2B)表示。其中,重要的下调蛋白有:LAMB1、HNRNPH3、COX2、FADS2、GPATCH4、CAV1、AHNAK、PFKM、MAVS、ANP32E 等。重要的上调蛋白有:EPPK1、ITPR1、AKR1C2、MFF、PTGS2、PNPLA6、AKR1B1、ITGA1、PTGFRN、TGM2 等。

图2 TUC338 影响宫颈癌细胞中蛋白差异表达的分析
2.2.2 TUC338 对蛋白基因本体功能的影响 GO 富集注释结果柱状图显示,BP 前3 位为:膀胱介导转运(vesicle-mediated transport)29 个蛋白,转运(transport)45 个蛋白、细胞内蛋白转运(intracellular protein transport)45 个蛋白;MF 前3 位为:钙离子结合(calcium ion binding)79 个蛋白、蛋白激酶活性(protein kinase activity)85 个蛋白;CC 前3 位为:细胞外区域组分(extracellular region)15 个蛋白、内质网组分(endoplasmic reticulum)17 个蛋白,见图3A;与rLV 组比较,rLV-TUC338 组中显著下调BP,包括:真空转运、细胞组分组装、自噬体组装等6 组(P<0.05)。MF 包括:磷酸转移酶活性,醇基作为受体、细胞色素-c 氧化酶活性、激酶活性等7 组。无显著的下调CC 蛋白见图3B。与rLV 组比较,rLVTUC338 组中显著的上调BP 包括:细胞铁离子稳态、脂代谢过程、磷脂分解代谢过程等9 组。MF 包括:三价铁结合、内肽酶抑制剂活性、蛋白谷氨酰胺γ-谷氨酰转移酶活性等9 组。CC 包括:细胞外区域部分、细胞外空间、细胞骨架等5 组,见图3C。

图3 基因本体功能分析

图3 基因本体功能分析(续)
2.2.3 KEGG 分析 KEGG 富集注释结果柱状图显示,共注释到了包括:细胞过程,转运和代谢(Transport and catabolism)313 个蛋白、真核生物(Cellular community-eukaryotes)162 个蛋白、细胞运动(Cell motility)75 个蛋白等,共44 个KEGG 途径。其中,注释到最多蛋白的途径为代谢总谱(541 个蛋白)见图4A。显著下调KEGG 通路有半乳糖代谢(Galactose metabolism)、氧化磷酸化(Oxidative phosphorylation)等共6 个(P<0.05),见图4B、表1。显著的上调KEGG 通路有肌动蛋白细胞骨架调节(Regulation of actin cytoskeleton)等共20 个,见图4 C,表2。

表1 KEGG 分析中重要的下调蛋白

表2 KEGG 分析中重要的上调蛋白

图4 KEGG 分析
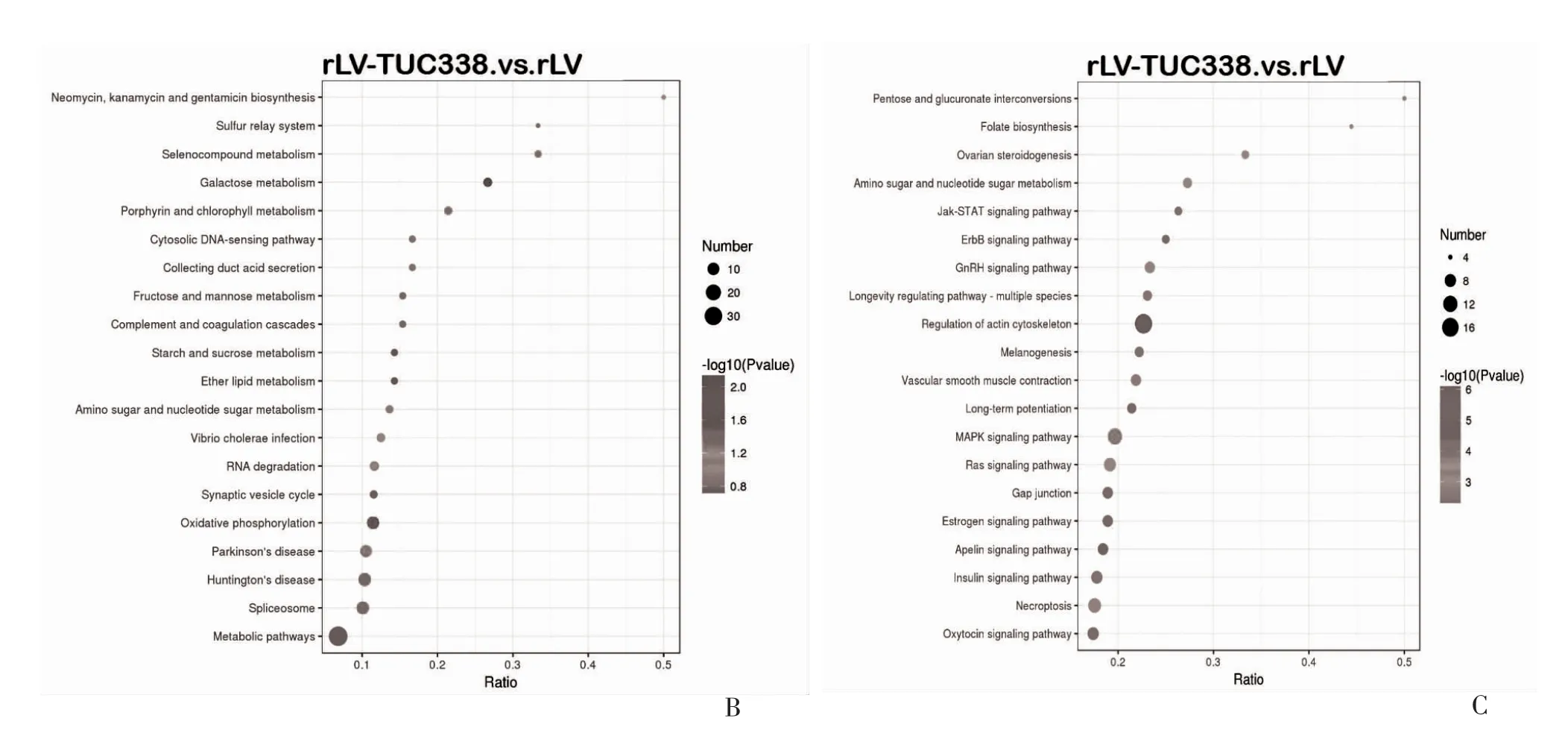
图4 KEGG 分析(续)
2.2.4 蛋白结构域分析 结构域注释结果柱状图显示共检测到含WD40 repeat 结构域(WD40 repeat)等20 个蛋白结构域,见图5A。rLV 组和rLV-TUC338组中有显著差异的结构域包括血红蛋白过氧化酶结构域,动物(Haem peroxidase,animal)等,见图5B。

图5 蛋白结构域分析
2.2.5 亚细胞定位分析 蛋白亚细胞定位占比统计分析显示,主要亚细胞成分为:核蛋白(nucleus protein)占34.23%、细胞质蛋白(cytoplasm protein)占16.16%、线粒体蛋白(mitochondrion protein)占14.22%、内质网蛋白占8.44%;剩余由细胞膜蛋白、细胞外蛋白、高尔基体蛋白、细胞骨架蛋白、溶酶体蛋白等构成,见图6A。差异蛋白亚细胞定位占比统计分析显示,主要差异亚细胞成分为:核蛋白(nucleus protein)31 个,占24.22%、细胞质蛋白(cytoplasm protein)23 个,占17.97%、细胞外蛋白(extracell protein)20 个,占15.62%,见图6B。

图6 亚细胞定位分析
2.2.6 蛋白互作分析 在rLV 组和rLV-TUC338 组中,下调的互作网主要包括:prot1(P37268)-prot2(O95864)、prot1(Q6FGH9)-prot2(Q13363)等。上调的互作网主要包括:prot1 (P60520)-prot2(A0A024RBQ5)、prot1(P60520)-prot2(Q06330)等,见图7。

图7 蛋白互作网络图
3 讨论
差异蛋白分析发现Lnc RNA TUC338 下调了Hela 细胞的一些蛋白,从多种途径抑制肿瘤发展。其中PFKM 是糖酵解重要调节酶之一,负责催化6-磷酸果糖磷酸化为1,6-二磷酸果糖。糖代谢是支持肿瘤发展的重要代谢途径,PFKM 在多种肿瘤中呈高表达状态,比如在头颈部鳞状细胞癌(HNSCC)中被确定为靶基因[6]。PFKM 对糖酵解的代谢重编也使乳腺癌细胞增殖、迁移和侵袭能力提高[7]。在宫颈癌中,PFKM 也存在异常高表达[8]。另一下调蛋白ANP32E即酸性核磷蛋白32 家族成员E,是一种组蛋白伴侣,负责调节各种基因的表达。已有研究证实其与乳腺癌的发生相关[9];在甲状腺癌的研究中,ANP32E 上调激活AKT-mTOR-HK2 信号通路,增强糖酵解,促进甲状腺癌细胞增殖和迁移[10]。这些与PFKM 协同,均提示Lnc RNA TUC338 可降低糖代谢,从而抑制宫颈癌细胞的增殖。Lnc RNA TUC338 上调了一些蛋白,其中透明相关formin 1(DIAPH1)是一种激动蛋白成核酶,参与细胞骨架的调节,可与纺锤体-微管(MT)高亲和力结合,起到稳定染色体的作用[11]。这种作用机制与紫杉醇相同,且DIAPH1 可协同紫杉醇稳定微管,增加卵巢癌对紫杉醇的药物敏感性[12]。DIAPH1 的上调提示Lnc RNA TUC338 通过改变细胞骨架通路,稳定细胞状态,抑制肿瘤进展。
GO 分析发现,Lnc RNA TUC338 下调了一些GO,其中重要的下调BP 为磷酸化。磷酸化途径中的膜相关酪氨酸/苏氨酸蛋白激酶1(PKMYT1)为膜相关激酶,是WEE 家族的一员,特异性磷酸化络氨酸(Tyr)15 和Thr14[13],络氨酸的磷酸化影响代谢酶活性,使肿瘤发展[14]。另外,PKMYT1 可通过磷酸化并灭活细胞周期蛋白依赖性激酶1,调控G2/M 期之间的转换,并对DNA 进行损伤修复,导致细胞周期失控,在多种癌症中均高表达[15]。rLV-TUC338 组中重要的下调MF 为磷酸转移酶活性;其中,丝裂原活化蛋白激酶激酶2(MAP2K2)磷酸化并激活MAPK1/ERK2和MAPK2/ERK3,在MAPK 信号转导中起到关键作用,促进有丝分裂。通过抑制MAP2K1 及MAP2K2可阻断该信号转导,从而抑制细胞增殖[16]。磷酸化及磷酸转移酶活性途径的下调,提示Lnc RNA TUC338抑制肿瘤增殖的机制,可能与抑制磷酸化过程有关。
KEGG 分析发现,Lnc RNA TUC338 下调了一些KEGG 通路。KEGG 中最显著的下调信号通路为半乳糖代谢。半乳糖是细胞代谢中必不可少的单糖,参与多种生物学功能。目前研究发现越来越多的疾病涉及半乳糖代谢[17]。半乳糖糖代谢在肿瘤发展过程中亦起到重要作用。其中磷酸葡萄糖变位酶1(PGM1),具有催化葡萄糖1-磷酸(G-1-P)和葡萄糖6-磷酸(G-6-P)之间互相转化的作用,参与糖原分解及合成,被认为是一种癌基因。在肺癌中,葡萄糖剥夺状态时,AMP 活化蛋白激酶(AMPK)作为PGM1 的上游调控因子,使PGM1 表达增强,促进糖酵解,强化肿瘤代谢,促进癌细胞进展[18]。敲低PGM1可显著降低糖酵解,抑制肿瘤细胞增殖[19]。KEGG 中最显著的上调信号通路为肌动蛋白细胞骨架调节。肌动蛋白细胞骨架有多种生物学功能,包括维持细胞内钙离子稳态[20]、通过氧化还原作用调节血管细胞的迁移、增殖以及收缩[21]等。肌动蛋白细胞骨架在肿瘤细胞中对细胞迁移能力有很大影响[22]。其中磷脂酰肌醇-5-磷酸4-激酶2 型α(PIP4K2A),涉及磷酸化及细胞骨架2 个途径。在磷酸化途径中,可磷酸化5-磷酸磷脂酰肌醇,参与调节多种重要的细胞功能,包括增殖、迁移、免疫、物质运输及葡萄糖摄取等[23,24]。在细胞骨架途径中,可以维持线粒体的结构及功能稳态[25]。有研究显示[26],PIP4K2A 为抑癌基因,其在脑胶质母细胞瘤中负向调节磷酸肌醇3-激酶(PI3K)信号传导,降解P85,从而抑制肿瘤生长。上述结果提示Lnc RNA TUC338 抑制肿瘤增殖,其机制可能与抑制半乳糖代谢及活化细胞骨架通路有关。
综上所述,Lnc RNA TUC338 改变了人类宫颈癌细胞的蛋白组学,是宫颈癌的抑癌基因。主要通过糖代谢、磷酸化、细胞骨架途径实现其抑癌作用。
致谢:感谢上海同济大学生命科学院博导、教授陆东东对本次研究的技术指导与论文撰写指导。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
天津医科大学学报(2019年6期)2019-08-13
生物学通报(2019年3期)2019-06-15
广州大学学报(自然科学版)(2019年1期)2019-05-07
分析化学(2017年12期)2017-12-25
天津科技大学学报(2016年1期)2016-02-28
中国学术期刊文摘(2016年2期)2016-02-13
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
安徽医科大学学报(2015年9期)2015-12-16
现代检验医学杂志(2015年2期)2015-02-06
