
天然多糖抑菌活性及机理研究进展
2022-07-18 12:52崔宏春
湖北农业科学 2022年11期
张 彤,赵 芸,黄 伟,崔宏春
(1.湖北工业大学生物工程与食品学院,武汉 430068;2.杭州市农业科学研究院,杭州 310015;3.湖北省药品监督检验研究院,武汉 430075)
近些年来,传染性疾成为世界范围内日益严重的公共卫生问题,每年报告的食源性疾病病例约4 800万例,东南亚和地中海东部流行率最高。食源性病原体是造成传染病的主体。尽管如此,食源性疾病并不局限于发展中国家,美国的食物链被认为是世界上最安全的,但每年仍有约128 000人因此住院、3 000人死亡[1,2]。
为了延长保质期,在食品中加入食品防腐剂(如亚硝酸盐和二氧化硫)是较为普遍的做法。然而,大多数食品防腐剂是通过化学合成开发的,长期食用合成防腐剂可能会对人体产生不良影响[3],需要寻找对人体无害的适合用于食品中的天然防腐剂。
高感染率和长期接触抗生素导致微生物对抗生素产生耐药性[4]。全球范围的微生物耐药性促使人们从海洋、陆地等寻找天然存在的候选药物、防污剂和食品保鲜剂[5]。因此,新的抗微生物药物的发现和开发受到广泛关注。
多糖的生物活性一直是国内外科学领域的研究热点,近年来,从植物、动物、真菌、海藻等自然资源中分离出的多糖因具有多种药理活性,如抗肿瘤[6]、免疫调节[7]、抗氧化[8]、抗炎[9]等而受到越来越多的关注。多糖是一种天然高分子聚合物,通常由10多种单糖通过糖链或支链连接而成,分子质量为数万Da甚至数百万Da,它们广泛存在于植物、微生物、藻类和动物中[10,11]。与蛋白质和多核苷酸类似,多糖是生命活动中必不可少的大分子,它们参与许多生物过程,如细胞间通讯、细胞粘附、细菌或病毒感染以及细胞免疫等过程。因此,多糖与多核苷酸、蛋白质、脂质一起构成生命科学中最重要的四大生物大分子[12]。
多种天然多糖对革兰氏阳性菌和革兰氏阴性菌均有抗菌活性。研究表明,壳聚糖比壳寡糖具有更高的抗菌活性,对大多数细菌的生长均有明显的抑制作用,且壳聚糖对革兰氏阳性菌的杀菌作用一般强于革兰氏阴性菌。由于壳聚糖可溶于大多数pH低于6的有机酸,因此研究的上限pH为5.9,在这个范围内,壳聚糖的抗菌活性受pH影响,pH越低,其抗菌活性越强[13]。同时,不同DD(脱乙酰度)的壳聚糖水溶液对大肠杆菌和金黄色葡萄球菌的抑制效果也不同,随着壳聚糖DD的增加,抑菌效果也增强[14]。
天然k-卡拉胶不具备抗菌活性,经氧化处理后对大肠杆菌、金黄色葡萄球菌、铜绿假单胞菌和单核细胞增生李斯特菌表现出抑菌效果,并且氧化型k-卡拉胶对以上4种细菌的最低抑菌浓度和最低杀菌浓度随着氧化水平的升高而逐渐降低。扫描电镜分析了细菌细胞形态变化,可知氧化型k-卡拉胶能破坏细菌细胞壁和细胞膜,能有效抑制细菌的生长[15]。
许多种天然多糖对革兰氏阳性菌和革兰氏阴性菌均有抗菌活性,来源广泛,具有生物相容性和明显无毒性,可作为新型抗菌剂广泛应用于食品、医疗、化妆品等行业,但目前关于多糖的抑菌机理尚没有确切的说法,本文从以下几个方面进行概述。
1 不同种类的抑菌天然多糖
目前,具有抗菌活性的天然多糖已广泛应用于医药、食品、环保、家用及个人护理用品等领域,如壳聚糖、黄原胶、黄芪多糖等。不同种类天然多糖的抑菌效果或机制、剂量及试验菌种见表1。
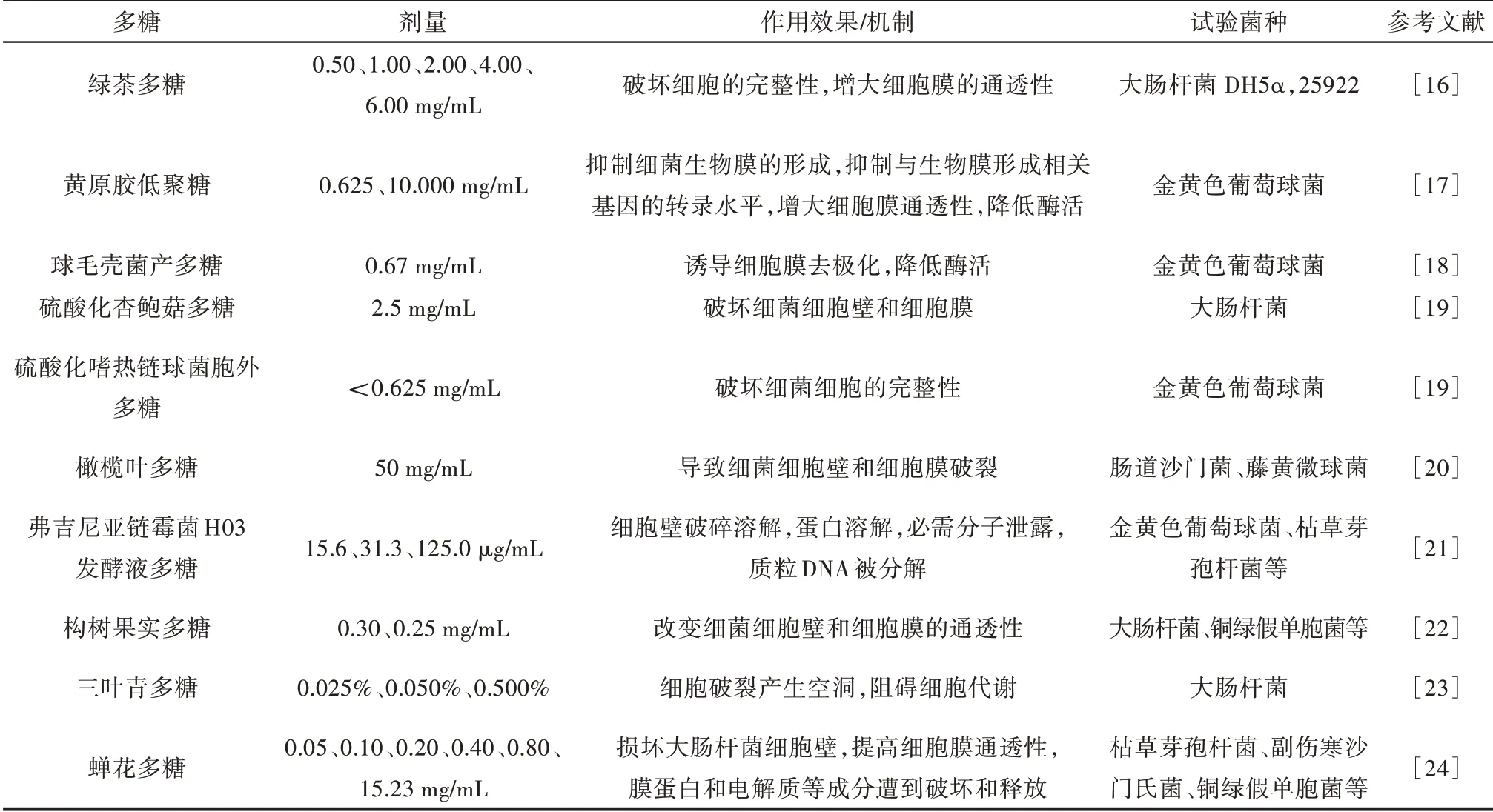
表1 具有抑菌作用的天然多糖
2 天然多糖的抑菌机制
2.1 破坏细菌细胞壁和细胞膜
细胞壁在细胞膜外是一个厚而坚韧的微弹性结构,它可以维持细胞形态,增加细胞的机械强度,还参与一系列生理活动,如物质运输、减少蒸腾、防止水分流失等[25,26]。细胞膜为细胞的生命活动提供了一个相对稳定的内部环境,同时也起屏障的作用。膜两侧的物质不能自由通过,细胞膜会选择性地运输物质。细胞与周围环境的物质交换是通过细胞膜的转运功能来实现的。细胞膜在细胞与外部环境之间的能量转换和信号转导过程中也发挥重要作用[27-29]。
2.1.1 增大细胞壁和细胞膜的通透性 Wang等[17]采用碱性磷酸酶(AKP)、β-半乳糖苷酶2个指标观测多糖对细胞壁和细胞膜通透性的破坏,AKP存在于细胞壁和细胞膜之间,β-半乳糖苷酶位于细胞膜内。试验结果表明,空白对照组和添加了黄原胶低聚糖的试验组培养基中AKP活性在2~6 h基本维持不变,这说明金黄色葡萄球菌的细胞壁是完整的;对照组的β-半乳糖苷酶活性基本不变,而试验组的β-半乳糖苷酶活性显著增加。说明黄原胶低聚糖能提高金黄色葡萄球菌的细胞膜通透性,但对细胞壁几乎没有影响。
Zhou等[16]使用NPN探针的荧光强度作为指标来探测细胞外膜通透性的改变,以ONPG作为指标测定β-半乳糖苷酶浓度来观察内膜是否受损,结果表明绿茶多糖(gTPC)可提高大肠杆菌的外膜通透性,但对大肠杆菌细胞内膜通透性无显著影响。总的来说,gTPC增强了大肠杆菌外膜的通透性,破坏了大肠杆菌细胞膜的完整性。
Meng等[30]使用抑菌圈法测定了核桃分心木多糖对大肠杆菌、铜绿假单胞菌、金黄色葡萄球菌、粪肠球菌的抗菌活性,结果表明核桃分心木多糖对革兰氏阴性菌和革兰氏阳性菌均有显著的抗菌活性,其抗菌活性随浓度的增加而增加。多糖可以增大大肠杆菌和金黄色葡萄球菌细胞壁和细胞膜的通透性,这种变化导致细胞内水溶性蛋白质迅速增加,从而阻止营养物质的输入。He等[21]的试验结果也表明多糖提高了金黄色葡萄球菌细胞质膜的通透性。
以上说明多种天然多糖能够提高细菌细胞膜的通透性,导致细胞内水溶性蛋白渗漏,营养物质无法输入,从而达到抑菌效果。
2.1.2 破坏细胞壁和细胞膜的完整性 Chen等[24]探究了三叶青多糖对大肠杆菌的抑菌机制,SEM结果表明,与未添加多糖的空白组相比,添加了三叶青多糖的试验组的细菌发生了细胞破裂,有的细菌表面有空洞,这表明多糖破坏了细胞壁和细胞膜的完整性。
He等[21]从弗吉尼亚链霉菌H03发酵液中分离得到一种新的多糖,并对金黄色葡萄球菌进行抑菌试验。TEM结果表明,经多糖处理过的金黄色葡萄球菌细胞壁总是破碎、变厚并出现一些溶解现象,这表明抗菌多糖对细胞壁有严重的影响。此外,金黄色葡萄球菌的形态发生了很大改变,细胞变得不规则、皱缩和相互粘附,同时金黄色葡萄球菌上清液水溶性蛋白含量迅速增加,这表明金黄色葡萄球菌的细胞可能被多糖破坏,导致蛋白溶解、必需分子渗漏和细胞死亡。
Khemakhem等[20]也发现橄榄叶多糖对肠道沙门菌和藤黄微球菌有较强的抑菌作用,多糖的这种最高抗菌活性可能是由细菌的细胞壁和细胞膜破裂引起的。Zhou等[16]通过扫描电镜和透射电镜观察经绿茶多糖(gTPC)处理后的大肠杆菌,发现大肠杆菌细胞形态发生明显变化,部分细胞壁变模糊,细胞质内容物渗漏,细胞膜表面粗糙、不规则,并且通过流式细胞仪进一步证实gTPC破坏了大肠杆菌细胞膜的完整性。
由此可见,天然多糖的加入能够破坏细菌细胞壁和细胞膜的完整性,细胞壁破碎变厚溶解,细胞内容物流出,菌体形态也变得不规则、皱缩,必需分子渗漏从而达到抑菌的效果。
2.2 阻碍细菌代谢
代谢是细菌最基本的特征之一,包括细胞内所有的化学和物理反应。细菌的代谢包括合成代谢和分解代谢。分解代谢为合成代谢提供能量和原料,而合成代谢是分解代谢的基础。
根据Kochanowski等[31]的研究,大肠杆菌生长与FBP(1,6-二磷酸果糖)浓度有关,大肠杆菌增殖速度越快,FBP浓度越高。果糖-6-磷酸(F6P)通过6-磷酸果糖激酶-1(6-PFK-1)的催化作用转化为FBP,是糖酵解的重要过程。
Chen等[24]研究表明,三叶青多糖的加入抑制了6-PFK-1酶活性,即抑制了F6P的二次磷酸化,从而阻碍了大肠杆菌的糖酵解和糖异生过程,使大肠杆菌不能获得足够的能量物质,如三磷酸腺苷(ATP)或还原态烟酰胺腺嘌呤二核苷酸(NADH+),从而导致大肠杆菌的生长受到抑制。
2.3 抑制细菌生物膜的形成
细菌生物膜(Bacterial bioflim,BBF)也称生物膜,是生活在细胞外聚合物基质中的微生物聚集体,它们通过细胞外聚合物分子粘附在生物和非生物表面,除非迅速清洗,否则无法去除,是与游走态细胞相对应的另一种存在方式[32]。微生物生物膜是否会在无生命或固体表面形成,是一种胞外多糖基质形成的结果,它为生物膜中微生物的相互作用提供了强度[33]。细菌在生物膜状态下比游走状态更加难以应付,生物膜的形成是导致抗菌治疗失败的重要原因[34]。因此,控制生物膜的形成或者消除成熟的生物膜是抗菌的防治策略之一。
Wang等[17]采用结晶紫染色法测定了黄原胶低聚糖对金黄色葡萄球菌生物膜形成的影响。结果表明,空白对照组随着时间的增加,595 nm处的吸光度也不断增加,而添加了最低抑菌浓度的黄原胶低聚糖的试验组,随着时间的增加吸光度始终维持在较低水平,这表明多糖抑制了金黄色葡萄球菌生物膜的形成。
2.4 抑制酶活性与提高细胞内离子水平
钙离子是所有细胞中各种细胞过程的重要调节因子,是细胞质功能的核心因子[35]。钙离子在细胞质中的积累导致细胞内活性氧水平的增加,从而导致细胞质功能障碍和细胞凋亡[36]。
Ca2+-Mg2+-ATP酶是细胞膜上的一种钙泵,它能水解ATP,将细胞内钙离子泵送到细胞外,维持细胞内钙离子浓度相对较低,从而保持细胞的稳定性并确保细胞的正常功能。
Wang等[17]研究表明,未添加多糖的空白对照组Ca2+-Mg2+-ATP酶的活性基本维持不变,而添加了黄原胶低聚糖的试验组的酶活性明显下降,同时细胞质中钙离子浓度显著线性增加。这表明黄原胶低聚糖抑制了金黄色葡萄球菌细胞膜Ca2+-Mg2+-ATP酶活性,从而导致细胞内钙离子不能及时转移至细胞外,细胞质内钙离子累积,从而对金黄色葡萄球菌的生长起到了抑制作用。
Wang等[18]发现GCP作用于金黄色葡萄球菌细胞膜的Ca2+-Mg2+-ATP酶活性0~8 h明显下降,这说明GCP能有效抑制Ca2+-Mg2+-ATP酶活性。同时,GCP处理的金黄色葡萄球菌细胞质中钙离子的相对水平在0~8 h呈线性增加,表明Ca2+-Mg2+-ATP酶活性受GCP的限制,细胞质中钙离子的积累会导致细胞内活性氧含量增加和细胞质功能紊乱,从而导致细胞凋亡,达到抑菌效果。
2.5 降低基因转录水平
Wang等[17]测定了与金黄色葡萄球菌生物膜形成相关的基因,未添加黄原胶低聚糖的对照组中,金黄色葡萄球菌的fnbA、fnbB和clfB3个基因的相对转录(RT)水平均在1.0左右,最低抑菌浓度的黄原胶低聚糖干预4 h后,金黄色葡萄球菌这3个基因的RT水平明显降低。结果表明,黄原胶低聚糖对金黄色葡萄球菌生物膜形成相关基因具有抑制作用。
2.6 降解质粒DNA
He等[21]发现多糖可以与质粒DNA结合,然后分解成小块。多糖对质粒DNA有显著的降解作用,因此,DNA可能是多糖抑菌的作用靶点。
2.7 耗散细胞膜电位
Wang等[18]使用产自球毛壳菌的多糖(GCP)对金黄色葡萄球菌进行抑菌试验,研究了GCP对膜电位的影响,发现GCP能耗散膜电位,引起金黄色葡萄球菌细胞膜去极化。细胞膜去极化将通过细胞质膜电压门控钙离子通道(Cch1/Mid1复合物)和其他未知转运通道诱导摄取钙离子[36],从而诱导细胞凋亡和程序性细胞死亡。
2.8 多糖的高铁螯合作用
由于铁是细菌生长的必需元素,因此通过清除环境中的铁来抑制细菌对铁的吸收是多糖可能的抗菌机理,具有高铁(III)亲和力的螯合剂已被证明具有抗菌活性[37,38]。
Shao等[39]研究表明,浒苔羧甲基化降解多糖(CDPE)通过羧基和羟基与铁结合,具有较低的铁(Ⅲ)结合亲和力,而浒苔异羟肟酸化降解多糖(HCDPE)能以相对高的亲和力结合铁(Ⅲ)。与CDPE相比,HCDPE对金黄色葡萄球菌、枯草芽孢杆菌、沙门氏菌和铜绿假单胞菌表现出更强的抗菌活性,也具有更低的最小抑菌浓度值。
3 影响多糖抑菌效果的因素
虽然具有抗菌活性的天然多糖已被广泛发现,但仍有大部分的天然多糖被证实不具有或具有微弱的抗菌活性,天然多糖的结构与其生物活性紧密相关。通过改变原有结构,能够使部分多糖呈现出较强的抗菌活性,因此探究抗菌多糖的构效关系,揭示多糖的抗菌活性和其结构之间的关系,成为当下的一大研究热点。
3.1 分子质量对抗菌活性的影响
Fleita等[23]从红藻中提取了高分子质量的S1组分多糖和低分子质量的S2组分多糖,并对金黄色葡萄球菌进行抑菌试验。结果显示,S1部分组分比标准抗生素显示出更高的抑菌活性,而S2组分没有显示任何抗菌作用。
Han等[22]采用滤纸片琼脂扩散法进行了构树果实多糖(BPPs)对4种细菌的抑菌试验。结果表明,在3种多糖组分中,相对小分子质量的BPP-3对大肠杆菌、铜绿假单胞菌、枯草芽孢杆菌和金黄色葡萄球菌的抗菌活性最好,且BPP-3的MIC(最低抑菌浓度)值低于其他2个组分。
3.2 聚合度对抗菌活性的影响
Ho等[40]使用木糖多糖和木糖寡糖测定人体粪便细菌对其体外的发酵能力,发现低聚合度的木糖寡糖比高聚合度的木糖多糖更易于被有益细菌吸收利用。
3.3 多糖改性对抗菌活性的影响
通过硫酸化、乙酰化、硒化、羧甲基化等化学修饰方法改变多糖的取代基,能够使部分多糖表现出较强的抑菌活性。Li等[19]对杏鲍菇多糖(PEPS)和嗜热链球菌胞外多糖(ST1275 EPS)进行硫酸化处理,并测定硫酸化多糖和粗多糖的抗菌活性。结果表明,硫酸化PEPS的抗菌活性强于PEPS,硫酸化ST1275-EPS的抗菌活性强于ST1275-EPS,这表明磺化是提高多糖生物活性的一种有效的改性方法,通过在多糖中引入硫酸基,可以提高PEPS和ST1275 EPS对细菌细胞壁和细胞膜的破坏能力,从而使硫酸化PEPS和ST1275 EPS具有更强的抗菌活性。
Moratti等[41]总结了壳聚糖化学改性的几种方法,发现壳聚糖的季铵盐化,如N,N,N-三甲基壳聚糖碘化物,对几种细菌的MIC值较未改性壳聚糖低;壳聚糖的羧甲基化则提高了壳聚糖的溶解度,O-羧甲基化壳聚糖(OCMC)比壳聚糖更能抑制细菌的生长。
4 结论及展望
具有抗菌活性的天然多糖种类很多,但其作用机制并无显著差别,主要通过破坏细菌的细胞壁和细胞膜、调控细菌内酶活性和离子水平、调控能量代谢、影响基因等方面,达到抑菌效果,并且其抗菌活性与分子大小、聚合度、取代基等构效关系密切相关。
目前,天然多糖抗菌方面的研究主要集中在抗菌活性及作用机制上,对多糖的构效关系研究较少,研究中尚存在一些问题亟待解决,因此开展多糖构效关系研究对抗菌功能性食品的研发具有重要意义。
猜你喜欢
创新作文(1-2年级)(2018年2期)2018-09-13
创新作文(小学版)(2018年4期)2018-07-06
浙江工业大学学报(2017年5期)2018-01-22
学习报·教育研究(2017年33期)2017-08-31
广东农业科学(2017年5期)2017-08-29
中国医疗美容(2015年4期)2015-04-27
科学中国人(2015年13期)2015-02-28
食品工业科技(2014年23期)2014-03-11
食品工业科技(2014年9期)2014-03-11
华东师范大学学报(自然科学版)(2014年3期)2014-03-11
