
异养细菌作用下岩溶水体惰性有机碳变化及其环境影响因素分析
2022-07-15 03:37何若雪孙平安
地球学报 2022年4期
何若雪, 李 强, 于 奭, 孙平安
1)成都工业学院, 四川成都 611730;2)中国地质科学院岩溶地质研究所, 自然资源部/广西岩溶动力学重点实验室, 广西桂林 541004
2020年联合国大会一般性辩论上, 我国承诺力争在 2030年前达到 CO2排放峰值, 努力争取 2060年前实现碳中和。已有研究表明, 2010—2019年平均每年的碳收支不平衡量达 0.1×1015gC·yr-1(Friedlingstein et al., 2020)。寻找遗失碳汇是平衡大气CO2收支的关键问题。除海洋和大气外, 陆地生态系统是最有可能的容纳汇, 其中岩石风化由于其时间尺度、化学反应速率和稳定性问题, 在全球碳收支评估中的角色和地位一直被低估(蒲俊兵等,2015), 尤其是碳酸盐岩风化。近年来, 大量研究表明岩溶水体中的浮游植物可以利用岩溶水高浓度的作为碳源进行光合作用, 并将其转化为颗粒有机碳(POC, Particulate Organic Carbon)沉降。但也有研究指出, 大部分POC在沉降途中即被降解转化为 CO2, 以至每年最终埋藏的有机碳大约仅为初级生产力的0.1%(Jiao and Zheng, 2011; Stocker et al.,2013)。除颗粒有机碳外, 溶解有机碳(DOC, Dissolved Organic Carbon)也是水体有机碳存在的主要形式, 其中 95%是“生物难以降解”的惰性溶解有机碳(RDOC, Recalcitrant Dissolved Organic Carbon),可以在深层水体中保存数千年, 形成长期碳储, 在全球碳循环中起到重要作用(张传伦等, 2019), 而可以被水体微生物降解和利用的 DOC即为生物有机碳(BDOC, Bioavailable Dissolved Organic Carbon)。为此, 焦念志基于海洋中微型生物生态过程的研究提出了“微型生物碳泵”理论, 指出异养微型生物在利用 DOC的同时, 会产生部分 RDOC, 是海洋RDOC的主要贡献者(Jiao et al., 2010)。
异养细菌利用有机质进行呼吸作用, 维持异养生长(张霞等, 2012), 是水生生态系统的重要组成部分, 是水体有机质降解和转化的主要驱动者(王海丽等, 2020)。研究表明, 海洋中约有200亿吨浮游植物释放的有机碳被异养细菌迅速吸收利用, 是海洋碳循环的重要组成部分(Williams, 1998)。其中,好氧不产氧光合异养细菌(AAPB, Aerobic Anoxygenic Phototrophic Bacteria)由于其独特的生态功能,受到广泛关注。1979年, Shiba在日本海湾首次发现了好氧不产氧光合异养细菌(Shiba et al., 1979),AAPB保留了不发达的光合系统, 好氧异养兼有光合作用, 在依靠呼吸消耗有机质维持生长代谢的同时, 还能以光能作为补充, 光合作用产生的ATP(Adenosine Triphosphate)既减少其呼吸代谢的能量需求, 还增加了进入细胞的DOC量, 在减少呼吸作用对现有有机碳消耗的同时使更多 DOC留存在水体中。另一方面, 有研究表明AAPB细胞表面的黏性使其更易与其他颗粒物形成聚合物沉降, 加强基于沉降的生物泵作用(焦念志等, 2011)。
已有研究表明, 异养细菌更倾向于利用浮游植物产生的 DOC(张海涵等, 2020), 而岩溶水偏碱和高含量的特征为浮游植物提供了充足的无机碳源(Liu and Dreybrodt, 2015; 杨明星等, 2017), 对浮游植物产生施肥效应, 使其生物量增大, 进而使流域内源有机碳比例增大(韩翠红等, 2020), 为异养细菌提供丰富的有机质底物。但目前对惰性有机碳的研究主要集中于海洋(Lønborg et al., 2018)、河口(Dang and Jiao, 2014)、土壤(Liang and Balser, 2011;Liang et al., 2017; Roth et al., 2019)。李强等(2019)对岩溶水体惰性有机碳的存在机理进行探讨, 认为岩溶筑坝河流中存在“生物惰性有机碳”。肖琼等(2020)对漓江惰性有机碳进行了研究, 结果表明漓江地表水体有平均2.42 mg·L-1的RDOC, 且受DOC浓度和水生生物活动影响, 存在明显的时空变化。尽管异养细菌的生物量和初级生产力均受到环境影响, 但其在环境因子的影响下如何变化需要进一步探讨。
为此, 本文于2016年6月和9月对广西壮族自治区柳江进行了监测采样和室内培养, 利用δ14C示踪法进行初级生产力培养实验、利用荧光实时定量PCR法测定研究区细菌基因丰度、利用原位微生物法进行RDOC培养实验, 并结合水体理化性质对流域内浮游生物初级生产力、细菌丰度和RDOC的变化和相互作用进行探讨, 以期为岩溶水体惰性有机碳研究提供一定的理论支持。
1 材料与方法
1.1 研究区概况
柳江位于广西壮族自治区北部(图1), 流域面积 5.84×104km2, 年平均流量 1.28×103m3·s-1。碳酸盐岩分布面积 4.93×103km2, 主要分布于流域西南一侧, 流域内出露地层主要包括: 晚泥盆系灰岩(D3)、早石炭系大塘组灰岩(C1d、C1d3)、中石炭系灰岩(C2d)、晚石炭系灰岩(C3)、晚二叠系碳酸盐岩夹碎屑岩(P2d、P2h)、早三叠纪碎屑岩夹碳酸盐岩(T1)、白垩系下统碎屑岩夹碳酸盐岩、第四系更新世(Q、Qp)、全新世(Qh)松散岩类。柳州市工农业发达, 人口集中, 是广西最大的工业城市。
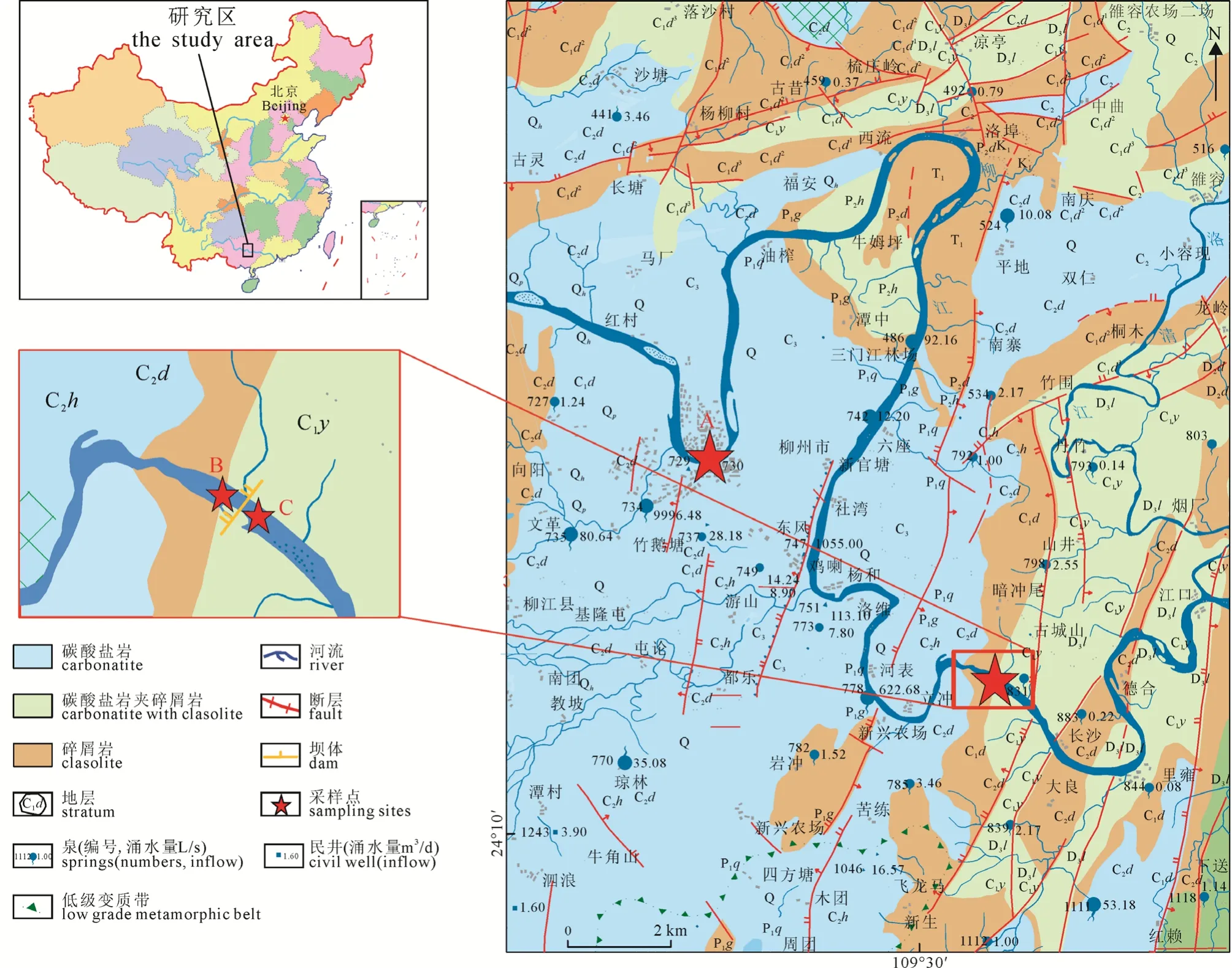
图1 采样点示意图Fig. 1 A sketch of the sampling site
本研究选取柳州市广雅大桥(A点)、红花水电站坝后(B点)和红花水电站坝前(C点)三个断面(图1), 分别于2016年6月和9月在各采样点进行分层(0m、-5m、-10 m)采样和监测。
1.2 研究方法
1.2.1 现场监测
水体物理性质监测利用法国 Ponsel公司ODEON多参数水质监测仪现场监测水体水温(WT)、溶解氧(DO)浓度、pH 值、电导率(EC), 精度分别为 0.1℃、0.01 mg·L-1、0.01个 pH 单位、1 µS·cm-1; 利用YSI EXO2多参数水质监测仪现场测定浊度、叶绿素a浓度, 精度分别为0.01 FNU、0.01 ng·L-1。
初级生产力测定将过滤水样(3 μm)和未过滤水样分别装入经过灭菌处理的西林瓶中, 加入标记过的NaH14CO3作为示踪剂, 进行48 h原位悬挂培养后, 过滤出水样中的颗粒有机质并测定其δ14C放射性强度, 分别计算得到水体细菌及总浮游生物的固碳速率, 计算公式(国家海洋局 908专项办公室,2006)如下:

其中,DMP为同位素计数率,DMPlive为所测样品计数率,DMPkill为加入 HgCl2后无生物作用的计数率背景值,DMPorg为碳源本底计数率,DIC为水体溶解无机碳浓度,T为实际培养时间。
1.2.2 采样及样品测试
水化学样品用于主要溶解离子、TOC、DOC测试的水样储存在 596 mL聚乙烯采样瓶中, 每次采样前用水样清洗采样瓶3~4次。阴离子采用离子色谱仪(861 Advanced Compact IC Metrohm)进行测试, 阳离子采用ICP-OES光谱仪进行测试, 精度均为0.01 mg·L-1, 测试工作在自然资源部岩溶动力学重点实验室完成。
碳同位素样品利用直径47 mm的GF/F滤膜(0.65 μm)过滤水样并记录过滤体积。将滤膜置于装有浓盐酸的干燥皿熏蒸24 h去除无机碳后, 用去离子水将滤膜清洗至中性后置于 50℃烘箱中 12 h恒温烘干, 送样至国家海洋局第三海洋研究所测试。
微生物样品利用经灭菌处理的直径47 mm混合滤膜(0.22 μm)过滤水样并记录体积, 滤膜用于微生物DNA提取。DNA提取采用MP(USA)公司土壤DNA快速提取试剂盒(Fast DNA® SPIN KIT FOR SOIL), 按操作进行。利用Realtime PCR实时荧光定量 PCR方法测定水体细菌和 AAPB丰度, 细菌16S rRNA基因和AAPB pufM光合基因定量PCR定量体系和反应程序见表1。
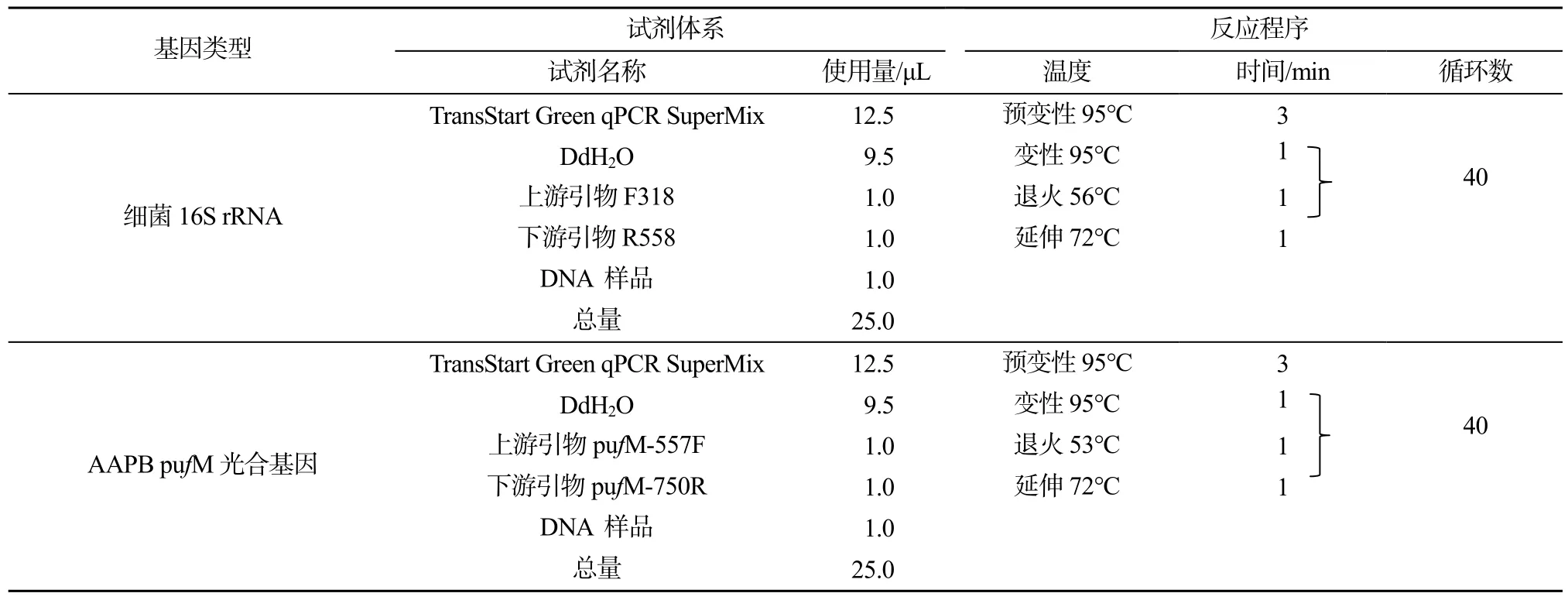
表1 细菌16S rRNA基因和AAPB pufM光合基因定量PCR定量体系和反应程序Table 1 The realtime qPCR formula and its reaction procedures of 16S rRNA and pufM gene
1.2.3 惰性有机碳培养
惰性有机碳培养参考 Mermillod-Blondin et al.(2015)的方法。将过滤水样(47 mm 混合膜, 孔径0.22 μm)装入经灭菌处理的玻璃试剂瓶中, 以1: 100的比例加入研究区原生接种液, 进行恒温暗培养。每组样品设置1个同比例加入接种液的超纯水样和1个不加入接种液的超纯水样以减少实验误差。每30天测试一次水体DOC浓度, 直到前后两次测试的DOC浓度变化幅度小于5%, 即认为此时的DOC为生物难以利用的惰性有机碳。DOC浓度采用总有机碳分析仪进行测定, 每 10个样品后插入一个空白水样, 并随机加测标准样品进行空白测定和质量控制。每个样品重复测试 2次, 保证变异系数小于1%, 取平均值作为此次培养数据。
2 结果
2.1 水化学特征
研究区WT介于23.84~30.88 ℃之间, pH介于7.58~8.02 之间, EC 介于 86.9~218.9 μS·cm-1之间,三者均呈丰水期低、平水期高的趋势(图2a), 但平水期 A点 WT和 pH值最低, EC最高。DO介于6.16~8.34 mg·L-1之间, 浊度介于 3.62~28.91 之间,均表现为丰水期高、平水期低, 其中平水期B点DO浓度最高、A点浊度最高。
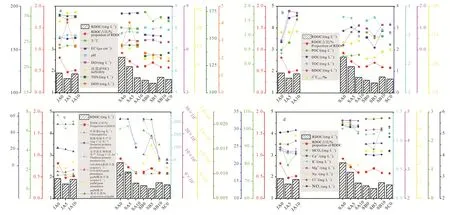
图2 研究区RDOC 浓度、水体理化性质、生物效应及碳源情况Fig. 2The RDOC concentration, hydrochemical parameter, biological parameter, andcarbonsources at the studyarea
研究区水化学类型为HCO3-Ca型, Ca2+为主要阳离子, 占平均总阳离子的72.88%, Mg2+为次要阳离子, 占19.09%;为主要阴离子, 占平均总阴离子的80.93%,为次要阴离子, 占11.39%。除Na+和 Cl-外, 各水体溶解离子浓度均表现为丰水期低、平水期高(图2d)。
2.2 细菌16S rRNA和AAPB pufM光合基因丰度
研究区细菌 16S rRNA 基因丰度(图2c)介于2.17×105~18.97×105copies·L-1之间, 丰水期平均值为 2.62×105copies·L-1, 平水期为 9.56×105copies·L-1,且表现为B点丰度最高。
pufM基因丰度(图2c)介于0.21×104~1.46×104copies·L-1之间 ,丰水期平均值为0.50×104copies·L-1, 平水期为 0.68×104copies·L-1, 表现为B点最高。
2.3 总浮游生物、细菌初级生产力
研究区浮游生物初级生产力(图2c)介于10.02~354.50 μgC·L-1·h-1之间, 丰水期平均值为302.88 μgC·L-1·h-1, 平水期为 166.39 μgC·L-1·h-1,表现为丰水期高、平水期低, 平水期表现为B点最高、C点最低。
细菌初级生产力(图 2c)介于 2.69~56.59 μgC·L-1·h-1之间,丰水期平均值为19.28 μgC·L-1·h-1, 平水期为 36.49 μgC·L-1·h-1, 表现为丰水期低、平水期高, 平水期C点初级生产力最低。细菌生产力占总浮游生物生产力的5.58%~32.16%, 丰水期平均占 6.47%, 平水期平均占23.81%。
2.4 有机碳分布特征
研究区 DOC 浓度(图2b)介于 2.10~3.58 mg·L-1之间, 丰水期平均值为 3.14 mg·L-1, 平水期为2.73 mg·L-1, 表现为 B 点最低。TOC 浓度介于2.38~3.82 mg·L-1之间, 丰水期平均值为 3.32 mg·L-1,平水期为3.37 mg·L-1, 表现为C点最低。POC浓度介于 0.27~1.55 mg·L-1之间, 丰水期平均值为0.93 mg·L-1, 平水期为0.65 mg·L-1, 表现为 B 点最高。
经实验培养(表2), 最终得到研究区 BDOC浓度介于 0.21~1.80 mg·L-1之间, 丰水期平均值为1.31 mg·L-1, 平水期为 0.87 mg·L-1, 无明显的沿流程变化趋势。RDOC浓度介于 1.46~2.66 mg·L-1之间, 丰水期平均值为 1.83 mg·L-1, 平水期为1.86 mg·L-1, 且表现为A点最高。RDOC占总DOC的 48.16%~92.61%, 丰水期平均占 60.78%, 平水期占67.99%, 表现为A点最高。
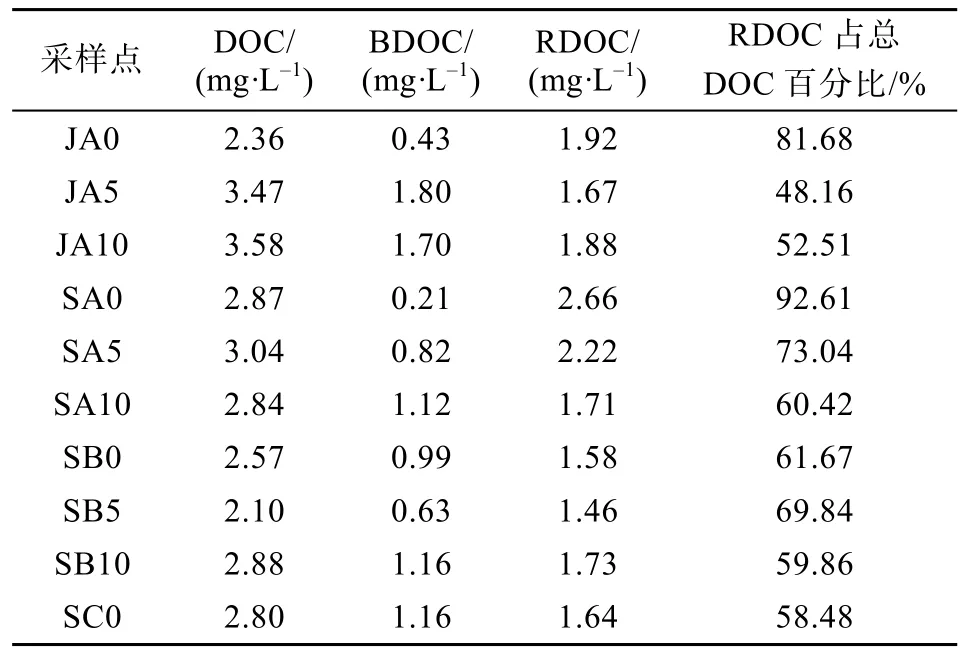
表2 研究区DOC、BDOC、RDOC含量Table 2 Concentration values of DOC, BDOC, and RDOC
2.5 环境因子对有机碳的影响分析
研究区 RDOC和 BDOC均表现出明显的季节变化和空间变化特征(图2), 与各环境因子间无直接的相关关系, 但其变化趋势与部分环境因子呈一定的响应关系。为分析各环境因子对流域内RDOC的具体影响, 利用 SPSS20.0对各环境因子进行主成分分析(表3)。
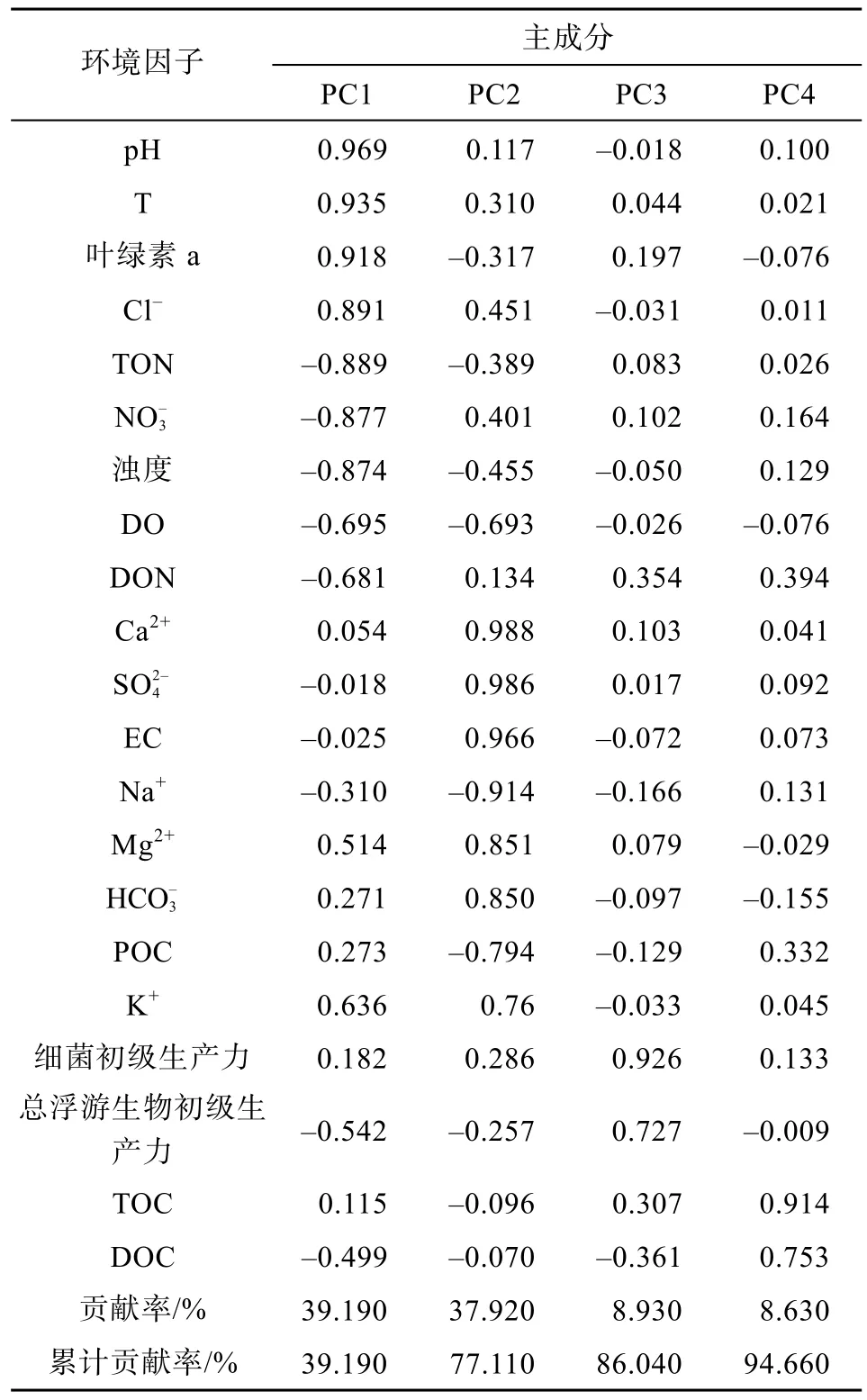
表3 各环境因子的载荷及方差贡献率Table 3 Component loading and variance contribution rate of the influential factors
经最大方差法旋转后, 获得四个特征值大于 1的主成分(PC1、PC2、PC3、PC4), 其方差贡献率分别为39.19%、37.92%、8.93%、8.63%, 累计贡献率达94.66%。
与PC1相关性最高的是pH、T、叶绿素a浓度、Cl-, 主要反应水体物理性质及浮游植物的影响。与PC2相关性最高的是 Ca2+、、EC、Mg2+和, 主要反应流域内硫酸参与的碳酸盐岩风化。与PC3相关性最高的是细菌初级生产力和总浮游生物初级生产力。与PC4相关性最高的是TOC浓度和DOC浓度。综上, PC1的部分因子和PC3、PC4代表研究区浮游生物和细菌的生物效应对RDOC的影响; PC1和PC2的部分因子代表水体理化性质的影响; PC4也代表流域内有机碳情况对RDOC的影响。因此, 通过研究区生物效应、碳源和水体理化性质的分析, 可探讨在环境因子影响下研究区惰性有机碳的响应机制。
3 讨论
3.1 生物效应对惰性有机碳的影响
浮游植物光合作用产生的有机碳是水体内源有机碳的主要来源, 也是细菌代谢所需有机质的主要贡献者, 而异养细菌是RDOC的主要贡献者, 因此浮游植物及细菌的生物量和活性可能是影响水体惰性有机碳含量的重要影响因素。
研究区细菌丰度、细菌初级生产力均表现出丰水期低、平水期高的季节变化特征, 与RDOC浓度变化特征一致(图2c), 表明细菌丰度越高、初级生产力越高, RDOC浓度越高, 即作为RDOC的主要贡献者, 细菌代谢活动越强, 其产生的 RDOC越多。
然而对RDOC浓度、细菌丰度和细菌初级生产力进行多元线性回归分析得到式(1):

统计分析结果与观察到的季节变化趋势相反。主要是由于平水期细菌丰度的空间变化特征与RDOC 浓度呈明显的负相关关系(R2= -902,p<0.05)。B点细菌和AAPB丰度最高但RDOC浓度最低, A点细菌和AAPB丰度较低但RDOC含量高,这一现象可能是水库筑坝所致。B点位于水库坝后,一般而言, 筑坝使水体流动速度明显变缓, 有利水生生物生长繁殖, 因此库区DOC浓度最高, 较高的DOC浓度可为异养细菌提供充足的有机质, 产生的RDOC也相应增多, 这在丁冰岚(2021)和肖琼等(2020)对大沽河和漓江的研究中得到证实。平水期B点叶绿素a浓度、细菌丰度、总浮游生物初级生产力及细菌初级生产力均较高, 也表明较缓的流速有利于水生生物生长代谢。然而, 图2c可见, 平水期B点的DOC和RDOC浓度明显最低。较缓的流速也有利于有机质沉降, 尤其部分细菌的胞外黏性分泌物使其更易形成较大聚合物, 导致细胞内留存的DOC也随之沉降(Shi et al., 2010)。研究结果(图2b, c)表明, 平水期A点和B点总浮游生物和细菌的初级生产力大致相同, TOC浓度也较接近, 但B点DOC浓度明显最低、POC浓度最高, 表明该点浮游生物生产的有机质大部分形成沉降, 水体中部分RDOC也随之沉降, 留存的DOC较少, RDOC浓度也较低。汪旋等(2020)对太湖沉积物有机碳组分的研究表明惰性有机碳增加了沉积物TOC含量, 是沉积物碳库中较为稳定的组分。
异养细菌生长主要依靠内源有机质, 浮游植物光合作用产生的有机质为异养细菌代谢提供充足的碳源, 二者可能呈明显的正相关关系(乐凤凤等,2015; Ofrat et al., 2015; 贺成等, 2019)。但异养细菌生长也依赖于无机营养盐, 因此二者也相互争夺必须的养分, 产生竞争抑制(周进等, 2016)。多元线性回归分析得到式(2),R2较小(0.37), 表明浮游植物对RDOC可能更多地产生间接影响。

浮游植物利用DIC生成有机质和氧气, 因此往往采用叶绿素a和DO浓度表征生物量和生物碳泵效应强度(Beaver et al., 2018)。研究区叶绿素a浓度呈丰水期低、平水期高的季节变化特征(图2c), 一方面平水期水温较高, 促进浮游植物生长繁殖, 同时B点较缓的流速更利于浮游植物生长繁殖, 生物量增加, 叶绿素 a浓度明显升高。但总浮游生物初级生产力和 DO浓度却呈相反的季节变化趋势, 丰水期高、平水期低。丰水期水温较低, 较高的氧分压使水体中DO浓度升高(Boyer et al., 1999), 同时A点位于市区内, 人类活动产生大量的营养盐输入,其中氮是浮游植物重要的生源要素(叶琳琳等,2014), 图2a和 2d可见, 丰水期和平水期 A点和 DON 浓度都较高, 为浮游植物代谢提供丰富的营养盐, 该两点总浮游生物初级生产力较高,RDOC浓度也较高。
与细菌初级生产力相比, 浮游生物初级生产力季节性差异较小, 除平水期B点因水动力条件影响导致有机碳大量沉降外, 其余采样点均表现出浮游生物初级生产力高时、RDOC浓度较低的趋势(图2c)。已有研究表明浮游植物分泌的有机物较易降解(张霞等, 2012), 研究区丰水期A点浮游生物初级生产力最高, 但 RDOC浓度和所占比例相对偏低,BDOC浓度和所占比例较高, 表明浮游生物代谢产生的DOC活性较大, 更易被降解。平水期A、B两点浮游生物和细菌生产力均较高, 异养细菌利用浮游植物产生的DOC生长代谢, 产生RDOC, 因此该两点RDOC的相对浓度均较高。C点位于水库坝前,水位较浅且受水库放水扰动, 初级生产力明显下降,但可以观察到细菌初级生产力与总浮游生物初级生产力相比较高, 因此 RDOC浓度和所占比例也较高。
3.2 碳源对惰性有机碳的影响
有机碳是异养细菌代谢的重要能量来源, 异养细菌需要消耗有机碳以维持代谢生长, 不同碳源使淡水环境的细菌群落、活性、及对碳的吸收利用发生变化(赵吉睿, 2013; 叶琳琳等, 2014)。水体有机碳主要来源于陆源输入和内生有机碳, 其时空变化主要受控于流域内的化学侵蚀、大气沉降和水体初级生产力(原雅琼, 2016)。Ofrat et al.(2015)、贺成等(2019)对海洋中异养细菌的研究认为异养细菌更倾向于利用浮游植物产生的内源有机碳, 也有研究表明部分流域外源碳是浮游细菌生长代谢的主要碳源(Gao et al., 2007)。
研究区 RDOC浓度和所占比例与 POC、DOC浓度变化趋势均有明显的响应关系(图2b)。研究区RDOC浓度与POC浓度变化趋势相反, 与DOC浓度变化趋势基本一致, RDOC所占比例与二者均呈相反趋势, 多元线性回归分析(式3)也表明这一点。

POC一般来源于陆源输入和水生生物活动(李春川等, 2020)。研究区δ13CPOC值呈丰水期偏正、平水期偏负的季节特征, 与乌梁素海的变化特征相近(耿悦, 2021)。丰水期流域内机械侵蚀较强, POC多来源于降水补给带来的流域内植被和土壤有机碳,其δ13CPOC值介于-24.45‰ ~ -24.31‰之间, 与西江流域土壤有机质的δ13CPOC值(-27.46‰ ~ -26.88‰)相比明显偏正(Chen et al., 2005), 可能是陆源C4植被来源的有机碳进入河流系统所致(魏秀国等,2008)。平水期样品δ13CPOC值介于-29.04‰ ~-25.94‰之间, 相对偏负, 尤其B、C两点,δ13CPOC值介于-29.04‰ ~ -27.46‰之间, 明显低于西江流域的值, 可能主要受水生生物活动的影响。如上文所述, B点总浮游生物初级生产力和POC浓度均最高, 也说明平水期B点颗粒有机碳主要为内源碳。这一结果与Pierson-wickmann et al.(2011)和叶琳琳等(2014)对Rophemel水库、太湖有机碳来源的研究结果一致。
研究区 DOC浓度丰水期最高, 除陆源输入外,丰水期A点浮游生物初级生产力较高, DOC可能受到外源碳和内源碳的共同影响。平水期 DOC浓度在 B点最低, 加之水库环境使有机质更易沉降, 表现出DOC浓度最低、POC最高的特征。
由此可见, 内源有机碳是研究区异养细菌代谢的主要有机质来源。丰水期外源输入较多, 细菌丰度低、初级生产力较小, 因此RDOC所占比例偏小。平水期内源有机碳占比大, 细菌丰度和初级生产力增加, RDOC占比增大。
3.3 水体理化性质对惰性有机碳的影响
研究表明, 水温、水体透明度、无机营养盐等环境因子会影响水生生物群落结构和生态功能(Fernandes et al., 2014; Mao and Li, 2018; 李蕊等,2018)。另外Ni and Li(2019)的研究表明DOC的生物降解性也会受水温影响。
尽管 RDOC与水体物理因子无明显统计关系,但温度是影响水生生物, 尤其是细菌代谢生长的关键因子, 部分研究表明水体细菌丰度会随温度升高而增加(Ofrat et al., 2015; 张喆等, 2017)。研究区水温与RDOC浓度无明显相关性, 但与叶绿素a浓度呈显著正相关(R2=0.78,p<0.01), 表明水温升高促进浮游植物生长, 其生物量显著增加, 生产旺盛,初级生产力较高, 为异养细菌提供充足有机质。
除此之外, 丰水期水体浊度明显高于平水期(图2a), 与RDOC浓度的季节变化趋势相反。受降水补给影响, 丰水期陆源输入物质增多、水体浊度升高对光照条件产生影响, 不利于光合生物生长代谢。丰水期叶绿素a浓度和pufM基因丰度较低也表明这一点。平水期水体浊度较低, 更利于浮游植物和光合细菌光合生产, 使RDOC增加。
除水温、浊度等物理性质外, 水体溶解离子也对水生生态环境产生影响。DON营养盐输入有利于促进浮游植物代谢。营养盐除供给浮游植物光合作用、为异养细菌提供有机质底物外, 也会刺激细菌的呼吸作用(Yuan et al., 2010)。此外, NO3-N也是细菌重要的氮源。刘会亮等(2019)对光合细菌的研究表明部分无机盐能明显促进光合细菌生长。Fernandes et al.(2014)的研究也提出水体微生物的营养需求可以在相对低水平的无机营养盐条件下得到满足。研究区主要溶解离子表现出丰水期低、枯水期高的季节变化特征, 这是丰水期降水补给带来的稀释作用造成的。平水期空间变化表现出A点溶解离子浓度最大、B点最小, A点位于市区内, 受人类活动影响较大,浓度明显大于B、C两点也表明这一点。
研究结果表明, 除平水期B点受水体流速影响外, 其余采样点基本表现出溶解离子浓度越高、RDOC浓度越高的变化特征, 可能是营养盐供给充足的情况下, 更利于水生生物代谢, 产生RDOC。
此外, 本文RDOC培养实验在恒温暗环境下进行。与原生自然环境相比, 培养环境生物效应以呼吸作用和分解作用为主。实验中异养细菌不会面临捕食压力, 同时实验环境与原生环境的水动力条件有显著差异, 稳定的环境使异养细菌丰度维持在一个较高值(肖喜林, 2020), 从而消耗更多的DOC。因此, 今后需要进一步开展原位培养实验来厘定其结果的准确性。
4 结论
研究区RDOC浓度介于1.46~2.66 mg·L-1之间,平均1.85 mg·L-1, 占DOC的48.16%~92.61%, 平均占65.83%, 其浓度变化受流域内生物效应、碳源和部分水体理化性质影响。浮游植物是流域内BDOC的主要贡献者, 异养细菌是流域内RDOC的主要贡献者。
从季节变化而言, 流域内RDOC浓度和RDOC相对浓度均表现出丰水期较低、平水期较高的特征。平水期水温升高、水体浊度降低, 浮游生物初级生产力增加, 产生较多内源有机质, 为异养细菌提供充足的有机质底物, 加之细菌丰度和初级生产力增加, 以至RDOC的浓度和相对比例都明显增加。
RDOC浓度的空间变化特征与环境因子之间呈现不同的响应机制, 尤其在水库坝后的采样点。这主要是水动力条件变化造成的。位于水库坝后的采样点流速平缓, 有利水生生物生长代谢, 叶绿素 a浓度、细菌丰度、总浮游生物初级生产力和细菌初级生产力均明显增加。但较缓的流速使有机质和生物聚合物产生沉降, 导致留存在水体中的 DOC比例减少, RDOC和DOC均表现出与初级生产力相反的趋势。
Acknowledgements:
This study was supported by Guangxi Natural Science Foundation Program (No.2015GXNSFGA139010), and Chengdu Technological University (No. 2021ZR023).
猜你喜欢
河南科技(2021年14期)2021-11-28
广西文学(2021年1期)2021-03-03
中国卫生产业(2020年25期)2020-11-30
银幕内外(2020年5期)2020-11-25
飞天(2020年9期)2020-09-06
生物工程学报(2020年6期)2020-07-31
陶山(2019年2期)2019-12-15
大东方(2019年8期)2019-10-21
作文周刊(中考版)(2017年21期)2017-07-10
