
新疆地区绿僵菌菌株筛选及抗逆性研究
2022-07-14 04:13:16霍英慧叶小芳
环境昆虫学报 2022年3期
霍英慧,陈 娟,王 晗,叶小芳,于 非
(1.新疆师范大学生命科学学院,新疆特殊环境物种保护与调控生物学实验室,中亚区域跨境有害生物联合控制国际研究中心,乌鲁木齐 830054;2. 慕恩(广州)生物科技有限公司,广州 510700)
绿僵菌Metarhizium具有较强的致病力,防治效果好,对人畜和生态环境安全,因此被用于农业和林业的害虫防治(Zimmermann, 1993)。蝗虫为植食性昆虫,主要取食禾本科、豆科及莎草科植物,且分布范围广,可以栖息在农田、沼泽、滨湖、草原、森林、荒漠、沙漠等多种环境,并具有迁飞行为。因此,蝗灾被认为是世界性严重的生物灾害之一(黄博,2012)。长期以来,人们主要是采用化学农药进行农业害虫防治,虽然在短时间内对害虫能够起到一定的防治作用,但是,随着用药次数的增多,“3R”问题(即土壤中农药残留(Residue),害虫产生抗药性(Resistence),生态平衡被破坏造成的害虫再猖獗(Resurgence))逐渐显现。利用生物或生物代谢产物来控制病虫草害的生物防治技术是害虫持续控制不可缺少的组成部分(陈学新,2010)。生物防治作为一种环境友好型的害虫防治方法,具有不污染环境、不易产生抗药性等优点,越来越受到人们的关注(陶星虎,2014;Dara, 2019)。微生物农药是一种重要的农药类型,由于其毒性较小,对于生态环境的影响也比较小,且能够保护人畜安全,因此成为一种重要的生物防治手段(李增智和樊美珍,2000;王海龙,2017)。真菌杀虫剂能够与体壁接触对昆虫直接侵染,而不需要其他外力的帮助,从而实现杀虫的目的(Mecoy, 1981; Roberts, 1981; Gillespieetal., 2000)。绿僵菌作为一种昆虫病原真菌,是一种有效的真菌杀虫剂,已被广泛应用于多种害虫的生物防治,效果显著(张宗炳和曹骥,1990;郭素萍,2004)。
目前,国内外已有较多关于绿僵菌的生物学特性和致病机理的研究。温度、紫外线、干旱均会影响绿僵菌的防治效果。温度在一定程度上影响绿僵菌的生长及致病力,温度过高或者过低均造成绿僵菌产孢量及致病力下降;紫外线和干旱也会对孢子的萌发产生阻碍作用,使得菌株的致病力降低。前期研究表明,新疆地区有丰富的绿僵菌资源(段玉林,2017),本研究通过高温、紫外线和干旱的强度测试来筛选出抗性和环境适应性强的菌株,从而提高防治蝗虫的效果。本课题组前期从野外土壤中分离、采集的菌株,经毒力测定挑选出5株高毒力的绿僵菌菌株。本文旨在测定这5株绿僵菌的抗逆性,筛选出致病力高且抗逆性好的菌株,为下一步开发适应于新疆高温、强紫外线、干旱生境蝗虫防治的绿僵菌生物农药提供依据。
1 材料与方法
1.1 高毒力绿僵菌菌株的筛选
1.1.1供试菌株
本实验室前期在新疆南北疆地区采集土壤样品,并从中分离得到绿僵菌、经初步筛选后保存的17株生长性状较好的菌株。将供试菌株接到PDA平板后在26.0±1.0℃培养,待产生分生孢子后备用。
1.1.2供试昆虫来源和饲养
东亚飞蝗Locustamigratoriamanilensis,新鲜蝗卵于培养箱中孵化后在培养室内以新鲜小麦苗和玉米草连续人工饲养至3龄,饲养条件为温度28.0℃~30.0℃、相对湿度40%~70%、光周期L ∶D=16 h ∶8 h。
1.1.3孢子悬浮液的配置
配制1.0×108孢子/mL孢子悬液:取实验待用菌株,用0.05%Tween-80溶液洗脱孢子,经搅拌、震荡、过滤后计数,配制成1.0×108孢子/mL的孢子悬浮液备用。
1.1.417株绿僵菌菌株对东亚飞蝗3龄蝗蝻致病力的初步测定
取浓度为1.0×108孢子/mL的孢子悬浮液,将此浓度设3个重复,每个重复15头试虫。用微量注射器吸取上述悬浮液滴于3龄蝗蝻前胸背板,接种剂量为10 μL,对照组用等量0.05%Tween-80溶液接种。处理后放入养虫盒中,用新鲜小麦苗和玉米草作食料,每天更换1次。在饲养条件下连续观察10 d,每天定期检查及记录死亡的蝗蝻数量,并将死亡的蝗蝻进行体表消毒后在培养皿中培养7~14 d,根据虫体是否长出菌丝及菌丝形态确定蝗虫死亡的原因,统计死亡数、僵虫数,计算死亡率、僵虫率,进行数据统计分析。
1.2 高毒力绿僵菌菌株抗逆性的研究
选取上述实验筛选出的5株较高毒力的绿僵菌,进行抗逆性研究。
1.2.1孢子悬浮液的配制
制成1.0×106孢子/mL孢子悬液备用。
1.2.2绿僵菌菌株耐短时高温能力的研究
1.2.2.1 高温处理对绿僵菌悬浮液分生孢子萌发率的影响
将孢子悬浮液分别装在灭菌的试管中,装好后封口,分别放置在温度不同的水浴锅中恒温加热30 min,温度分别为30、35、40、45、50℃。对照组不做加热处理,每组3个重复。取1 μL孢子悬浮液涂布于PDA培养基平板上,封口后置于26.0±1.0℃培养环境中恒温培养3 d,统计各个平板中的菌落数,计算孢子萌发率。
1.2.2.2 高温处理对绿僵菌生长情况的影响
取1.2.2.1中处理过的孢子悬液10 μL滴于PDA平板中央,对照组滴等量未经处理的孢子悬浮液,分别设3次重复。置于恒温培养箱中26.0±1.0℃条件下培养,第3天至第7天每天观察菌落生长情况,测量菌落直径,记录数据。
1.2.3绿僵菌菌株抗紫外线能力的研究
1.2.3.1 紫外线处理对绿僵菌悬浮液分生孢子萌发率的影响
取孢子悬浮液1 μL均匀涂布于PDA平板,将平板置于超净工作台紫外灯下,分别照射1、2、4、6、10 min,后于26.0±1.0℃恒温培养箱中培养,以未经照射的作对照,每个处理设3次重复。培养3 d后统计记录各处理平板中菌落数,计算各菌株各处理的孢子萌发率。
1.2.3.2 紫外线处理对绿僵菌生长情况的影响
吸取孢子悬浮液10 μL滴至PDA培养基中央,置于超净工作台紫外灯下,分别照射1、2、4、6、10 min,将平板置于26.0±1.0℃恒温培养箱培养。以未经照射的作对照,每个处理设3次重复。第3天至第7天每天观察菌落生长情况,测量菌落直径,记录数据。
1.2.4绿僵菌菌株抗旱力的研究
以SDAY液体培养基为基础培养基,加入10%,20%,30%,40%的PEG200(聚乙二醇),以未加PEG200的处理为对照,加入孢子悬液后,在26.0±1.0℃下培养24 h,置于显微镜下观察10个视野,统计萌发数,测定各菌株萌发率,每个处理设3个重复。
1.3 数据统计分析
利用Excel 2016分别计算各处理组的累计死亡率、僵虫率、LT50;利用Excel 2016分别计算各处理组的菌落直径、孢子萌发率,利用0rigin软件作图分析。
2 结果与分析
2.1 17株绿僵菌菌株对东亚飞蝗3龄蝗蝻致病力的测定
17株绿僵菌菌株对3龄蝗蝻的致病力的初步测定结果,可以看出,17株供试绿僵菌菌株对东亚飞蝗均有一定的致病性且致病力有比较大的差异,平均僵虫率在40.00%~96.67%之间,LT50在2.92~9.95之间。其中,M1-17菌株的平均僵虫率达到了95.00%以上,且LT50仅为2.92;M1-13、M1-09的平均僵虫率达到90.00%,LT50为3.11和3.20;M1-16、M1-05的平均僵虫率达到80.00%,LT50为3.46和3.56,表现出对东亚飞蝗较强的致病性。初选出M1-17、M1-13、M1-09、M1-16、M1-05五株菌株为对蝗蝻高致病力的菌株(表1)。
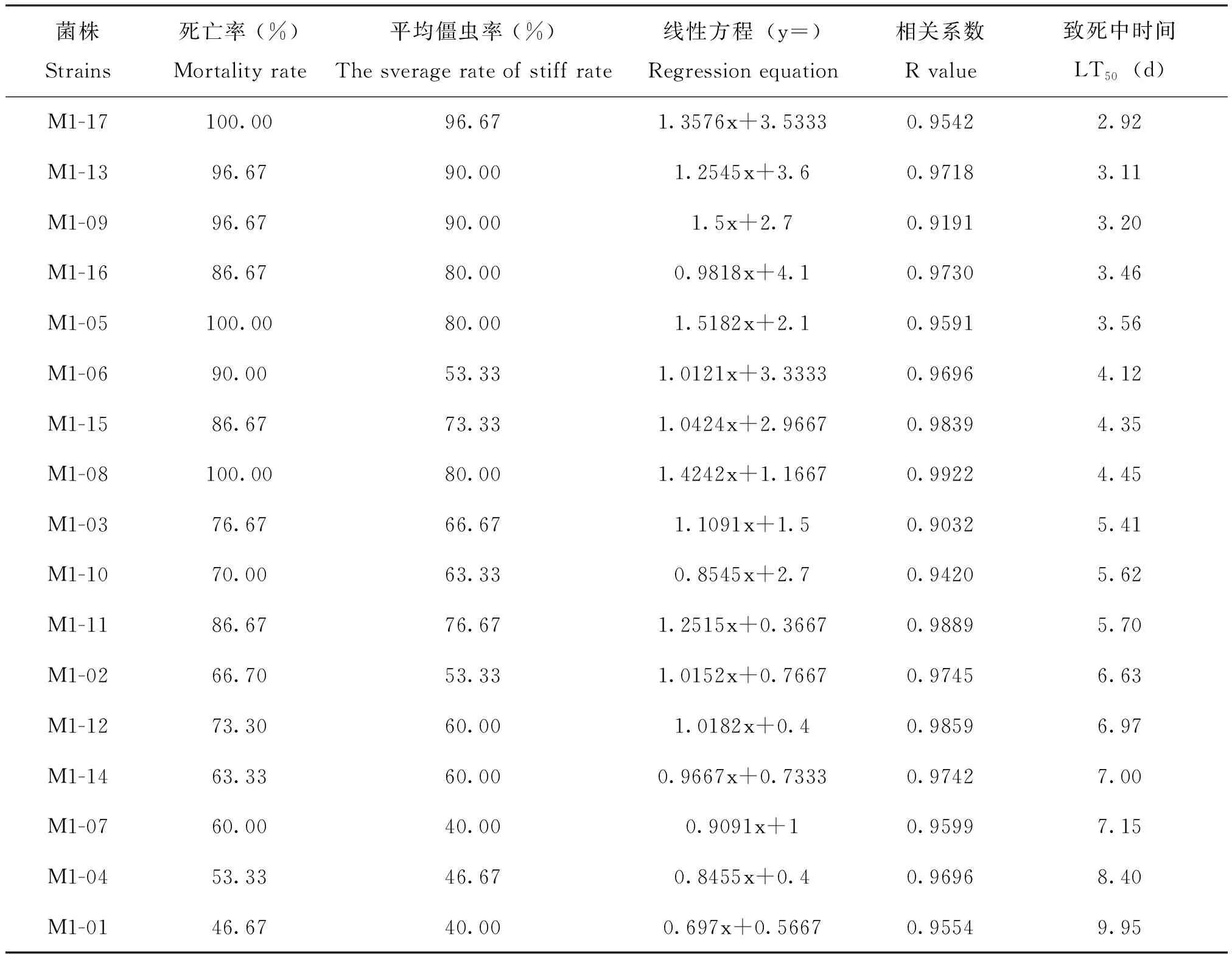
表1 17株绿菌菌株对东亚飞蝗的死亡率、僵虫率和致死中时间Table 1 Mortality,rigidity and LT50 of 17 Metarhizium strains to Locusta migratoria manilensis
2.2 对高毒力绿僵菌菌株抗逆性的测定
2.2.1绿僵菌菌株耐短时高温能力的测定
2.2.1.1 高温对绿僵菌孢子萌发率的影响
对初步筛选获得的5株菌株进行耐短时高温的测试,可以看出5株菌株分生孢子的萌发率随着温度升高而逐渐降低。在30℃和35℃处理时,萌发率差异较小,其中30℃处理时,萌发率最高的是M1-17,为86.55%;其次是M1-13,为83.45%;萌发率最低的是M1-16,为62.99%。40℃时M1-17仍有较高的萌发率,萌发率为78.65%;而M1-09和M1-16的孢子萌发率仅为40.00%。通过观察,供试菌株对高于45℃的高温较敏感,当温度为45℃时,菌株分生孢子的萌发率均可达到30%以上,其中M1-17萌发率最高,为63.57%。当处理温度为50℃时,5株菌株仍有分生孢子萌发,萌发率在18.36%~35.34%之间。绿僵菌M1-17和M1-13在50℃处理后表现出较好的耐高温能力,较其它3株菌株稳定(图1)。

图1 高温对绿僵菌分生孢子萌发率的影响Fig.1 Effect of high temperature on bourgeon rates ofMetarhizium strains
2.2.1.2 高温对绿僵菌生长量的影响
通过培养经高温处理后的5株绿僵菌菌株,分别于第3~7天观察菌落的生长情况并用十字交叉法测量菌落直径,观察各菌株生长情况。结果发现,5株菌株在未经处理时,生长情况差异不明显。随着温度升高,各菌株菌落的直径表现出不同。其中,菌株M1-05和M1-13的生长情况在30℃和35℃时优于对照,可能由于特定的温度促进了分生孢子的生长速度,在40℃和45℃时稍有减弱,当温度达到50℃,则生长十分缓慢;菌株M1-09和M1-17在温度低于45℃时生长情况大致相同,对低于45℃的高温抗性较好,在45℃和50℃时菌株生长情况稍有减弱,第3~4天生长较慢,在第5天开始迅速生长,初步推断可能由于高温推迟了孢子的萌发时间,但在孢子萌发后,菌株生长迅速;而菌株M1-16随着温度的升高,菌株生长态势明显减弱。各菌株对高温的抗性效果依次为M1-17>M1-05 >M1-13>M1-09>M1-16(图2)。
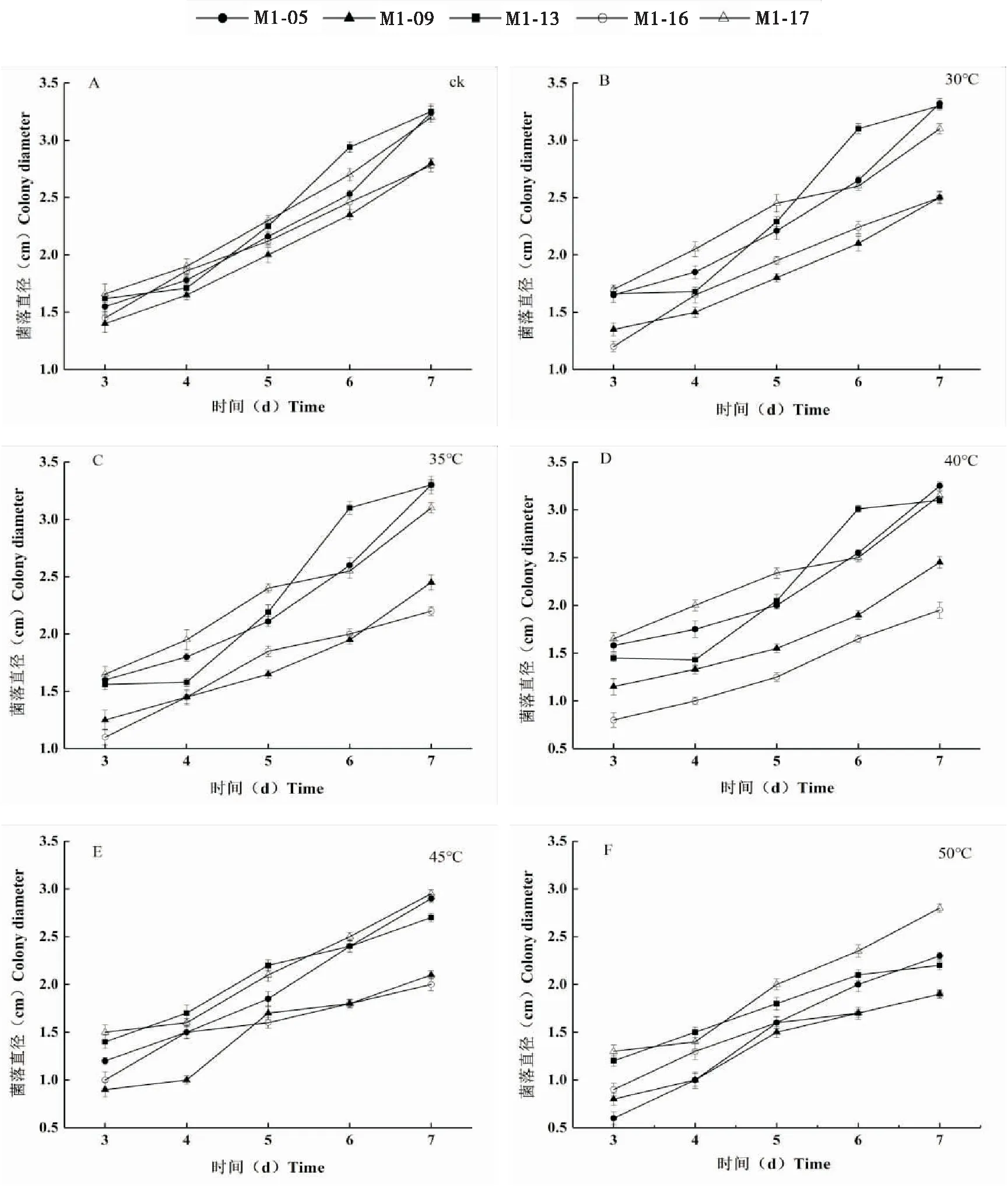
图2 高温对绿僵菌生长量的影响Fig.2 Effect of high temperature on diameter growth of Metarhizium strains
2.2.2绿僵菌菌株抗紫外线能力的测定
2.2.2.1 紫外线对绿僵菌孢子萌发率的影响
对获得的5株菌株进行抗紫外线能力的测试,结果发现,不同菌株经紫外照射后,均对孢子的萌发产生影响,5株不同菌株对紫外线处理均比较敏感,不同菌株间对紫外线抗性的差异十分明显。5株绿僵菌菌株的孢子萌发率基本都随紫外照射时间的增加而减小。在照射1 min时,菌株M1-17萌发率最高,为86.72%;M1-13萌发率最低为51.33%。在照射2~4 min时,菌株M1-16的孢子萌发率最高且差异不明显。总体来看,M1-16和M1-17菌株孢子的紫外线照射萌发率最高,紫外线照射对孢子萌发的影响最小,M1-09菌株次之。在紫外照射10 min时,菌株M1-17对紫外线的抗性最强,萌发率仍有13.40%,明显优于其它4株菌株(图3)。
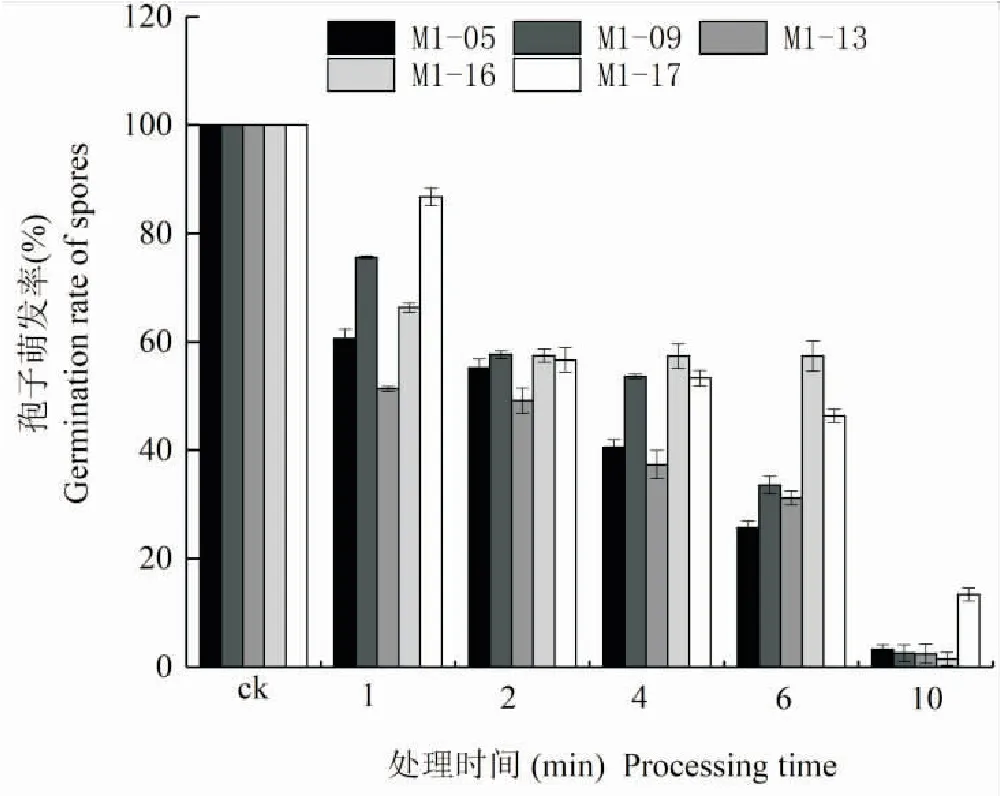
图3 紫外线对绿僵菌分生孢子萌发率的影响Fig.3 Effect of ultra-violet ray on bourgeon rates of Metarhizium strains
2.2.2.2 紫外线对绿僵菌生长量的影响
紫外处理对5株高毒力绿僵菌菌株生长量的影响结果可以看出,5株菌株在未经处理时,生长情况差异不明显。不同菌株经紫外照射后,均对菌株的生长产生影响,各菌株的菌落直径随紫外照射时间的增加而减小,不同菌株间对紫外线的抗性均有差异。在照射1、2、4 min时,生长情况最好的菌株是M1-05,最差的为M1-09。但随着照射时间的延长,在照射6 min和10 min时,生长情况最好的菌株为M1-17,最差的为M1-13。每株菌株经紫外处理,菌落生长虽受影响,但都保持着一定的生长量,对紫外光均有一定的耐受性,抗紫外能力依次为:M1-17>M1-16>M1-05>M1-09>M1-13(图4)。

图4 紫外线对绿僵菌生长量的影响Fig.4 Effect of ultra-violet ray on diameter growth of Metarhizium strains
2.2.3绿僵菌菌株抗旱力的测定
随着PEG200浓度的增加,菌株的萌发率逐渐降低。在PEG含量为10%时,对绿僵菌孢子萌发的抑制作用不明显,5株菌株萌发率均可达到85%以上,当PEG200浓度为40%时,对菌株孢子萌发率的影响与10%处理时有十分显著的差异,5株菌株的萌发率仅为22.47%~31.58%,其中,40% PEG200处理对M1-09的抑制作用最小,其次是M1-17和M1-16。在PEG的含量为40%时,绿僵菌M1-09、M1-17和M1-16的萌发率比较高,说明它们的抗旱能力较其它菌株高(图5)。
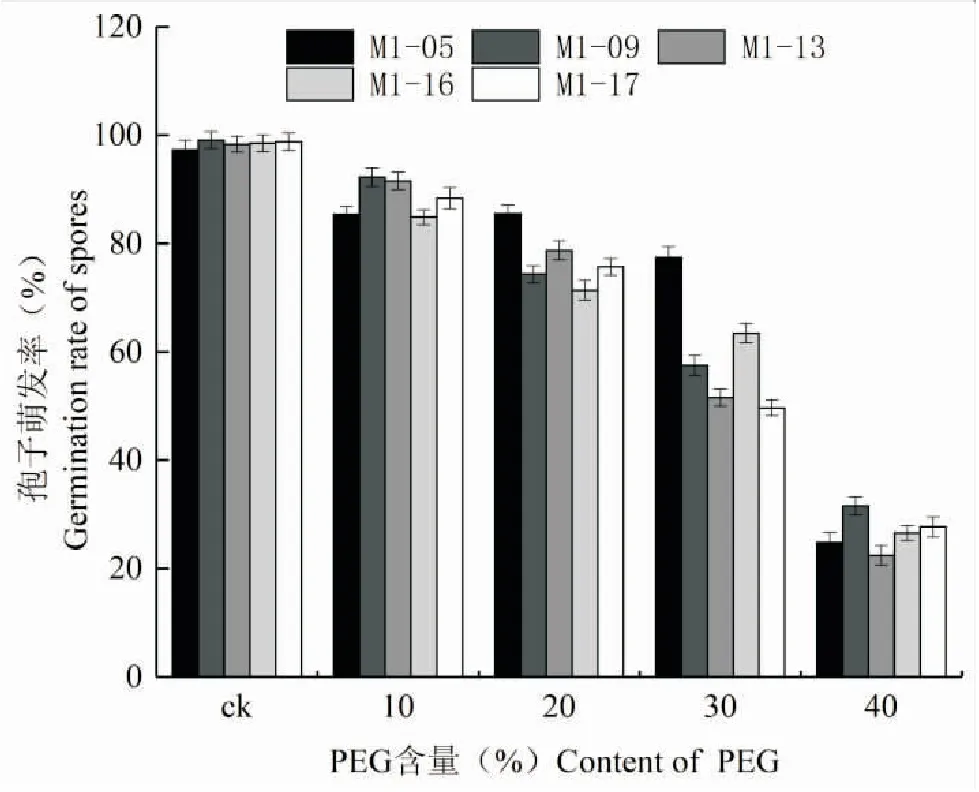
图5 PEG处理对绿僵菌分生孢子萌发率的影响 Fig.5 Effect of PEG on bourgeon rates of Metarhizium strains
3 结论与讨论
在自然界当中,绿僵菌分布的比较广泛,且数量较多,但是不同的菌株对于其寄主昆虫的毒力各不相同,且外部环境也会对菌株的毒力产生很大的影响,绿僵菌作为重要的昆虫病原真菌之一,具有无残留、对非靶标昆虫安全、可在田间持续存留、防治效果好等优点。
从本研究17株绿僵菌菌株对3龄蝗蝻的致病力的测定结果可以看出,17株供试绿僵菌菌株对东亚飞蝗均有一定的致病性且致病力有比较大的差异,平均僵虫率在40.00%~96.67%之间,LT50在2.92~9.95之间。初选出M1-17、M1-13、M1-09、M1-16、M1-05五株对蝗蝻高致病力的菌株。
致病力是衡量绿僵菌菌株优良性的重要指标,而菌株致病力的强弱受环境影响很大。其中,温度是影响绿僵菌致病力的一个重要因素。温度可以影响绿僵菌孢子萌发、菌丝生长、产生分生孢子及孢子活力,绿僵菌制剂在过冷或者过热的条件下都有可能丧失其活力或者毒力。温度会对致病速度、致病率产生影响,在一定的温度范围之内,温度的升高会促进孢子的萌发,使得孢子菌丝的生长速度加快,致病性也能有一定的增加。但是当温度过高,孢子萌发则会被抑制,还会使得孢子死亡。具有较高耐热性的菌株能够抵抗野外的高温,并且在环境温度方面的适应能力也更好。因此,对于孢子在高温胁迫下耐受力进行测试与分析,是对优良绿僵菌进行筛选的一个重要指标。本研究结果显示,5株菌株分生孢子的萌发率随着温度升高而逐渐降低。在30℃和35℃处理时,萌发率差异较小;45℃处理时,萌发率均可达到30%以上;50℃处理时,仍有分生孢子萌发。绿僵菌M1-17和M1-13在50℃处理时较其它3株菌株稳定。5株菌株在未经高温处理时,菌落生长情况差异不明显。随着处理温度升高,各菌株菌落的生长直径表现出不同。各菌株对高温的抗性效果依次为M1-17>M1-05>M1-13>M1-09>M1-16。
在自然生境当中,紫外线是另外一个对于绿僵菌致病力产生影响的重要因素。不同波长的紫外光及太阳光谱的组合成份对绿僵菌有不同程度的伤害(黄冬如,2004)。本研究结果显示,不同菌株经紫外照射后,均对孢子的萌发产生影响,5株菌株对紫外处理均比较敏感,萌发率基本都随紫外照射时间的增加而减小。总体来看,M1-16和M1-17菌株孢子的紫外线照射萌发率最高,紫外线照射对孢子萌发的影响最小,M1-09菌株次之。在紫外照射10 min时,菌株M1-17对紫外线的抗性最强,萌发率仍有13.40%,明显优于其它4株菌株。5株菌株在未经紫外处理时,菌落生长情况差异不明显。经紫外照射后,各菌株的菌落直径随紫外照射时间的增加而减小。每株菌株经紫外处理,菌落生长虽受影响,但都保持着一定的生长量,对紫外光均有一定的耐受性,抗紫外能力依次为:M1-17>M1-16>M1-05>M1-09>M1-13。
含水量也是影响孢子活力的一个重要因素,含水量过高,能够导致孢子提前萌发及自溶(石妍,2011),而含水量过低,能够导致孢子不萌发。本研究结果表明,不同菌株的抗旱能力不同,同一菌株在不同的干旱条件下抗旱能力也不同。随着PEG200浓度的增加,菌株的萌发率逐渐降低。PEG200浓度为40%时对菌株孢子萌发率的影响与10%处理时差异极其显著。在PEG的含量为40%时,绿僵菌M1-09、M1-17和M1-16的萌发率比较高,因此M1-09、M1-17和M1-16菌株的抗旱能力较其它菌株高。
综合以上指标可以看出,对高温的抗性效果较好的菌株为M1-17和M1-05;对紫外线的抗性效果较好的菌株为M1-17和M1-16;而菌株M1-09、M1-17和M1-16抗旱能力较好。研究结果显示菌株M1-17较其它菌株具有更好的抗逆性,更加适应新疆高温、干旱、强紫外线的环境,具有很好的开发利用价值。本研究通过对新疆本地绿僵菌资源的致病力测定及筛选,得到高致病力及高抗性的菌株,对蝗虫防治生物制剂的开发具有重要意义。
致谢:感谢新疆特殊环境物种保护与调控生物学实验室、新疆特殊环境物种多样性应用与调控重点实验室、干旱区植物逆境生物学实验室、中亚区域有害生物联合控制国际研究中心、新疆师范大学校级重点学科生物学学科、新疆师范大学沙漠藻研究院以及慕恩(广州)生物科技有限公司的大力支持与帮助。
猜你喜欢
湖北植保(2022年5期)2022-12-24 21:26:00
中国森林病虫(2021年6期)2021-12-20 08:45:42
中国生物防治学报(2021年1期)2021-05-11 10:31:06
中国森林病虫(2018年4期)2018-09-19 12:13:08
西部林业科学(2018年2期)2018-05-16 09:55:02
茶叶学报(2018年4期)2018-02-28 01:00:48
渔业致富指南(2016年12期)2016-11-11 07:07:18
科学大众(中学)(2015年9期)2015-10-12 09:37:53
少儿科学周刊·少年版(2015年3期)2015-07-07 19:30:33
塔里木大学学报(2015年1期)2015-04-25 02:38:44
