基于转录组测序的饲料添加淫羊藿对番鸭睾丸组织差异表达基因分析
2022-07-13 11:22:08施金虎孟宪娟卢立志许文武杜喜忠
中国畜牧杂志 2022年7期
施金虎,孟宪娟,卢立志,许文武,杜喜忠
(1.金华市农业科学研究院畜牧兽医与水产研究所,浙江金华 321017;2.浙江省畜牧技术推广与种畜禽监测总站,浙江杭州 317599;3.浙江省农业科学院畜牧兽医研究所,浙江杭州 310022)
睾丸是公番鸭生殖系统中重要的生殖器官,具有两大生理功能:一是通过精子发生产生精子;二是通过类固醇发生产生睾酮。精子发生是一个依赖于多种因素的复杂的细胞分裂过程,在睾丸的曲细精管中产生。雄性动物精子的形成受下丘脑-垂体-睾丸轴的调节,下丘脑分泌促性腺激素释放激素(Gonadotropin Releasing Hormone,GnRH),引发垂体释放促黄体生成素,促黄体素生成素能够刺激睾丸间质细胞增生,诱发睾丸分泌睾酮,影响雄性生殖器官的发育、刺激生精上皮的发育和精子发生,调节促性腺激素的分泌。精子形成的这个过程也同样受到很多基因的调控,基因家族由23 个成员组成,其在调节包括睾丸在内的几个生殖器官的生长和发育方面发挥重要作用。是家族的转录因子之一,已被证明是对哺乳动物的繁殖具有重要作用的基因。Yu 等发现在鸡的睾丸支持细胞中定位,与在哺乳动物中的定位一致,并且发现是表达上游支持细胞的重要标记物,是鸡胚胎睾丸分化发育和正常睾丸功能的潜在调节因子。Bai 等分别选取了24 只性腺分化前的莆田番鸭和性腺分化后的莆田番鸭进行转录组测序,发现有等9 个基因与睾丸的分化和发育有关,并且在性腺分化后,等基因在公鸭中特异性高表达。
淫羊藿是我国传统的中药药材,含有淫羊藿苷、淫羊藿次苷、黄酮、等多种微量元素,对生殖系统、性激素、性器官以及精液都具有保护作用。张长城等通过建立生精障碍的小鼠模型,发现在淫羊藿总黄酮的作用下,其睾丸质量、附睾质量、精子活力和精子密度均得到显著性提高,睾丸管腔内的精子数量增多,同时使生殖细胞形态得到了改善。韩贵芳等发现淫羊藿总黄酮使自然衰老大鼠的睾丸组织形态得到改善,生精细胞数量明显增多,推测淫羊藿总黄酮可能通过促进AMPK 磷酸化减轻其炎症反应。睾丸生殖功能的减退与支持细胞的功能衰退有着重要联系。
本研究通过在番鸭饲料中添加1%淫羊藿,首先研究其对公番鸭睾丸发育组织形态的影响以及调节体内激素分泌的影响,其次利用转录组测序技术(RNA-seq)挖掘实验组和对照组中番鸭睾丸的差异表达基因与相关通路,研究结果为淫羊藿对番鸭睾丸发育的影响以及调控机制提供理论基础。
1 材料与方法
1.1 样品采集与RNA 提取 本实验将60 只49 周龄公番鸭随机分为2 组,每组3 个重复,每个重复为10 只。其中A 组为对照组,饲喂基础日粮,B 为实验组,饲喂基础日粮的基础上添加1%的淫羊藿,基础日粮组成及营养水平见表1。饲喂75 d 结束后,每个重复随机挑选5 只种公鸭进行翅下静脉采血,共采取75 管血液,静置3~4 h 待血液凝固后,3 000 r/min 离心5 min,收集血清并转移至1.5 mL EP 管中,于-20℃保存备用,用于测定血清生殖激素和抗氧化指标。另外,每个重复随机挑选1 只种公鸭,每组3 只进行屠宰。屠宰结束后,取其左右侧睾丸,统一采集公鸭左侧睾丸组织样本用于进行组织形态学观察;统一采集公鸭右侧睾丸组织样本用于进行转录组测序。
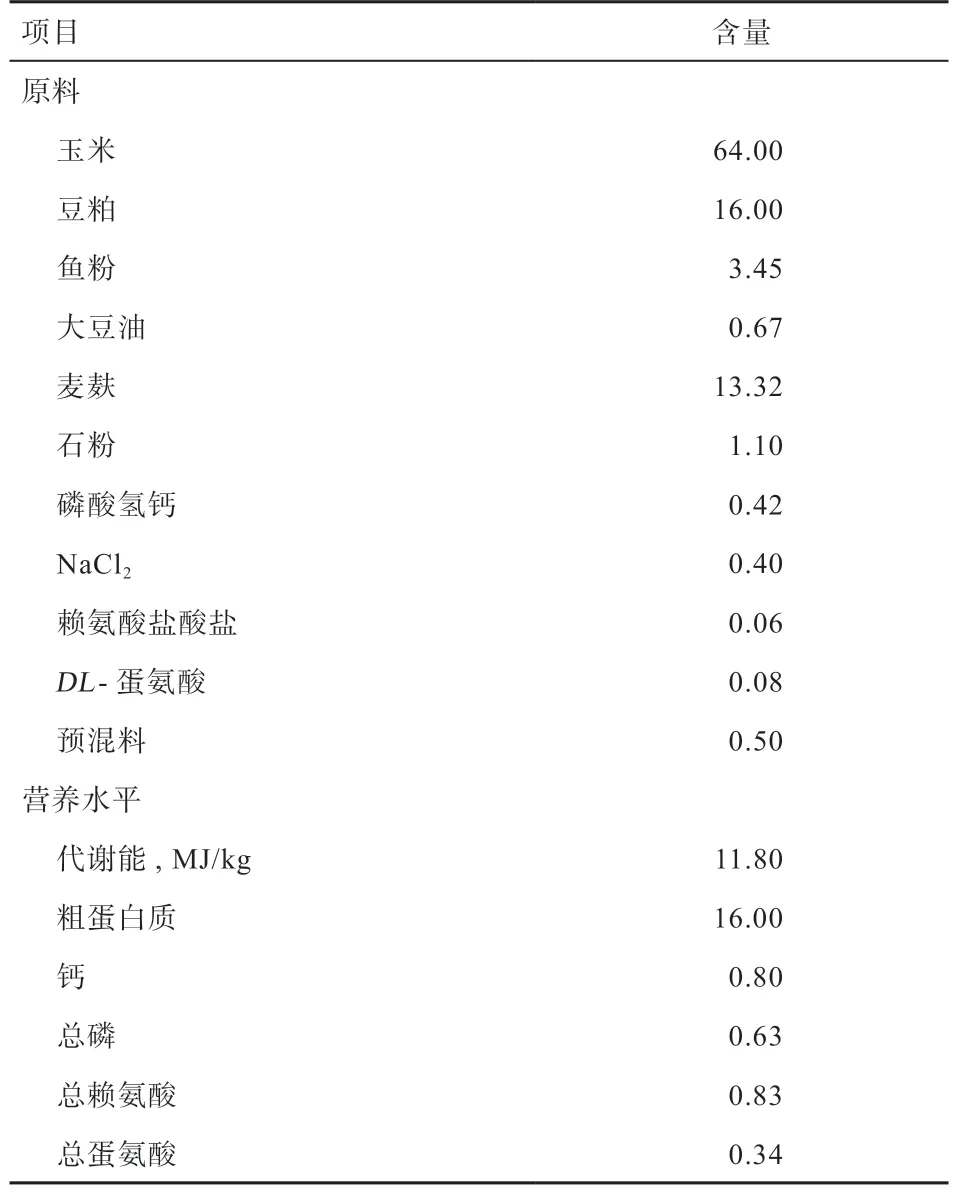
表1 基础日粮组成及营养水平(风干基础)
1.2 睾丸样本组织形态学观察 为了观察睾丸样本的形态学,本研究首先对番鸭睾丸进行石蜡切片制作。先将睾丸组织切成2 m的小块,用Bouin 液固定12 h;接下来将固定好的组织进行脱水,脱水后进行石蜡包埋,再进行切片;石蜡切片脱蜡后使用蒸馏水清洗,切片制作完成后分别使用苏木素和伊红染色,染色完成后进行封片。
1.3 血清生化水平指标检测 检测的血清生殖激素指标包括睾酮(T)、雌二醇(E)、促卵泡生成素(FSH)、促黄体生成素(LH),使用酶联免疫试剂盒进行测定,试剂盒由北京华英生物技术研究所提供。血清抗氧化指标包括超氧化物歧化酶(SOD)、丙二醛(MDA)、谷胱甘肽-过氧化物酶(GSH-Px)、总抗氧能力(T-AOC),使用生化分析仪进行分析,试剂盒由北京华英生物技术研究所提供。
1.4 RNA 的提取及转录组测序 RNA 的提取使用Omega 试剂盒。得到RNA 后本研究使用Nanodrop 2000 检测RNA 浓度、纯度和完整度,使用Agilent bioanalyzer 2100 记录样本的OD 值和RIN 值。RNA 提取后本研究使用PrimeScriptRT regent Kit With gDNA Eraser 试剂盒进行文库构建,构建好的文库使用q-PCR法进行定量,以保证文库的有效浓度,库检合格后使用Illumina Hiseq 平台进行150 个碱基对配对的双末端测序。
1.5 测序数据比对组装 将测序得到的原始数据使用Fastx_toolkit 软件进行质控和过滤,包括去除含有接头的Reads,去除低质量的Reads,从而得到高质量的测序数据(Clean Reads)。使用Hisat2 软件将Clean Reads 映射到鸭参考基因组中(CAU_duck1.0)。在已有参考基因组的基础上本研究使用stringTie 软件进行转录本组装拼接,与已有的转录本进行比较,获得没注释的新转录本。
1.6 差异表达分析 本研究使用Deseq2 软件分析实验组和对照组间的差异表达。采用FPKM(Fragments Per Kilobase of transcript per Million fragments mapped)作为衡量基因表达水平的指标,将|log2FoldChange|≥1且value<0.05 作为筛选标准获得差异表达的基因集。实验组差异基因表达水平高于对照组为上调基因(Up Regulated Gene),低于对照组的为下调基因(Down Regulated Gene)。
部分危险品码头因建造时间早于《装卸油品码头防火设计规范》(JTJ237-99)实施时间,消防改造设计时因充分考虑装卸货种和码头实际情况确定危险品码头分类等级,避免不必要地大规模改造码头,干扰码头的日常生产作业。
1.7 差异表达基因富集分析 Gene Ontology(GO)分析是大规模基因功能富集研究中比较常用的方法,包含3 个主要分支:生物学过程(Biological Process)、分子功能(Molecular Function)和细胞组分(Cellular Component)。KEGG(Kyoto Encyclopedia of Genes and Genomes)分析有助于把基因及表达信息作为一个整体,进一步解读基因功能。差异表达基因的GO 功能注释采用超几何分布的GOseq R 包实现,KEGG 通路分析采用KOBAS 在线网站完成。其中以<0.05 和富集基因数为标准筛选差异表达基因显著富集的GO 条目和KEGG 通路。
2 结果
2.1 睾丸组织形态学观察 番鸭睾丸组织的石蜡切片结果如图1 所示,对照组(图1A、图1B)生精上皮层数少,生精细胞数量少,管腔内的空腔面积大,成熟精子少;淫羊藿组(图1C、图1D)生精上皮层数多,生精细胞数量多,管腔饱满,成熟精子多。
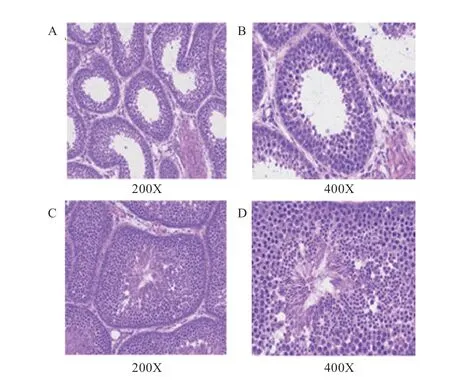
图1 对照组和淫羊藿组番鸭睾丸切片
2.2 血清生化指标水平 由表2 可见,血清生殖激素中,淫羊藿组睾酮含量、雌二醇含量、促卵泡素含量和促黄体素均要显著高于对照组。由表3 可见,血清抗氧化指标中,淫羊藿组的超氧化物歧化酶(SOD)含量、谷胱甘肽过氧化物酶(GSH-Px)含量、总抗氧化(T-AOC)水平均显著高于对照组;而丙二醛(MDA)含量显著低于对照组。

表2 对照组和实验组血清生殖激素含量的比较

表3 对照组和实验组血清抗氧化指标的比较
2.3 RNA 数据分析 通过Illimina 测序平台,本实验对睾丸样本的6 个cDNA 文库进行了高通量测序,经过质控后,每个样本平均可获得5.38×10个Clean Reads(表4)。本研究使用DESeq2 软件,以value<0.05且|log2FoldChange| ≥1 为标准筛选差异表达基因。本研究最终得到2 414 个差异表达基因,其中1 538 个基因显著上调表达,876 个基因显著下调表达(图2)。
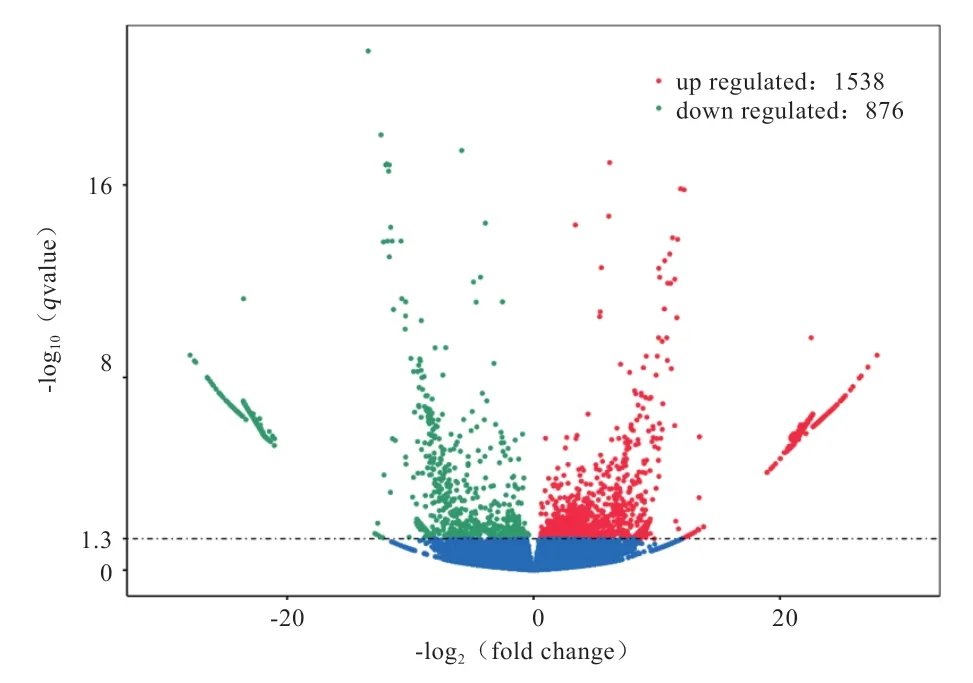
图2 实验组和对照组番鸭睾丸组织中差异表达基因火山图

表4 测序质量评估
2.4 差异基因的GO 注释及富集分析 对差异表达基因进行GO 功能注释,共发现了实验组和对照组显著富集的GO 条目有759 条,在生物学过程中多注释到白细胞激活(GO:0045321)、淋巴细胞激活(GO:0046649)、单个多细胞生物过程(GO:0044707)等过程,在细胞组分中多注释到受体(GO:0043235)、离子膜(GO:0005886)、细胞膜(GO:0009986)、细胞膜突触(GO:0120025)等,在分子功能上主要集中于转录因子、RNA 聚合酶II 核心启动子区域特异性结合(GO:0000982),转录激活因子、RNA 聚合酶II 核心启动子区域特异性结合(GO:0001077),RNA 聚合酶II 特异性结合(GO:0000977)(表5)。

表5 差异表达基因富集的GO 分类条目(前10 位)
KEGG 富集分析用于确定差异表达基因参与的主要生化代谢途径和信号通路。结果表明,差异表达基因显著富集在14 个通路中(图3),分别是细胞因子受体相互作用(ko04060)、铁死亡(ko04216)、黏附连接(ko04520)、溶酶体(ko04142)、Hippo 信号通路(ko04391)、轴突引导(ko04360)、矿物质吸收(ko04978)、鞘脂生物合成(ko00603)、抗原生成和传 递(ko04612)、Rap1 信号通路(ko04015)、Jak-STAT 信号通路(ko04630)、Hippo 信号通路(ko04390)、PI3K-Akt 信号通路(ko04151)、内吞作用(ko04144)。
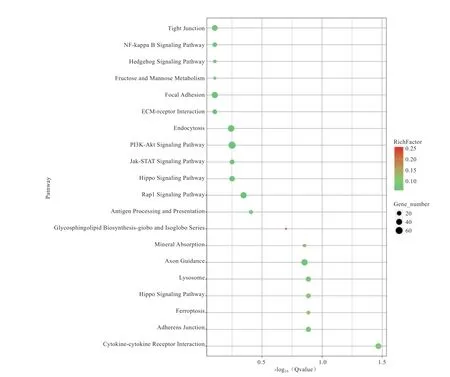
图3 差异表达基因显著富集通路气泡图
3 讨论
正常的睾丸组织结构和功能能够维持动物机体的繁殖能力。本研究通过制作石蜡切片,观察到实验组中生精上皮层数多,生精细胞数量多,管腔饱满,成熟精子多,说明淫羊藿的确可以促进番鸭睾丸的发育,促进精子的发生与成熟。激素调控在动物的繁殖生理过程中是必不可少的。种公鸭垂体分泌的促卵泡素和促黄体素、睾丸分泌的睾酮和雌二醇激素都是重要的繁殖激素,属于类固醇激素。本研究显示饲料中添加淫羊藿可以显著提高番鸭体内睾酮、雌二醇、促卵泡素、促黄体素的含量。有大量研究与本研究的结果类似,孙业波发现淫羊藿可以显著提高辽宁绒山羊种公羊血液中睾酮、促卵泡素和促黄体素的含量。淫羊藿中发挥作用的主要成分是淫羊藿苷,其可能是通过影响下丘脑-垂体-睾丸轴之间的平衡来诱导激素分泌。本研究还检测了实验组中血液的抗氧化指标,结果显示淫羊藿可以提高番鸭体内超氧化物歧化酶含量、谷胱甘肽过氧化物酶含量和总抗氧化水平,降低丙二醛含量。淫羊藿苷具有很强的清除二苯基-2-苦苯肼自由基(DPPH)、超氧阴离子自由基,还原力和总抗氧化能力,同时有较强的清除羟自由基的能力,这些能力赋予了淫羊藿苷显著的抗氧化活性。
通过差异表达基因进行GO 功能注释,本研究发现差异表达基因主要富集在生物学过程中的白细胞激活、淋巴细胞激活以及single-multicellular 组织进程等过程。相关研究表明动物的睾丸具有特殊的免疫环境,主要体现在它具有显著的免疫特权和有效的局部先天免疫的特性,睾丸炎的发生可能使免疫平衡受损,而白细胞和淋巴细胞对维持睾丸免疫平衡至关重要。郭明明等研究表明淫羊蕾对鸡的免疫功能有一定影响,高焕等研究表明淫羊蕾可提高淋巴细胞IL-2、IL-4 和IFN-mRNA 的表达,从而提高家禽的免疫能力,猜测淫羊蕾可能通过对睾丸免疫系统的改进从而提升番鸭的生殖能力。差异表达基因在细胞组分中主要注释到受体复合物、质膜、细胞膜、溶酶体腔等组分。激素受体在调节睾丸形成和雄性生殖细胞发育过程中有非常重要的作用,Heckert 等人证明FSH 受体mRNA 存在于成年大鼠的睾丸中,并且这种mRNA 水平在生精上皮细胞的周期中发生变化,还有如糖皮质激素受体和可溶性促性腺激素受体,ALK 受体家族对雄性睾丸和睾丸生精过程均具有调控作用。质膜和细胞膜是精子主要的膜结构,溶酶体腔泡对精子细胞和精母细胞释放入管腔内有重要作用,可能的机制是淫羊蕾通过对质膜、细胞膜、受体复合物、溶酶体腔等细胞组分形成的影响来影响番鸭精子的形成和发生。差异表达基因分子功能方面主要注释到转录因子活性、RNA 聚合酶特异性结合,DNA 结合等。相关研究表明诸多转录因子在动物睾丸发育过程中起到关键作用,其中转录因子Sp1通过调节睾丸中细胞周期蛋白A1(GATA1)特异性表达,Sox9、Sox5 和Sox13 转录因子可影响睾丸间质细胞的表达,Semba 等的研究表明由基因编码的转录因子HMG 对性腺发育不全有关。
本研究差异表达基因主要在细胞因子受体相互作用、铁死亡、黏附连接、溶酶体、Hippo 及jak-stat 信号等通路中富集,表明这些信号通路与番鸭睾丸发育和生精能力有关。Li 等人研究表明细胞因子和睾酮调节原代精母细胞在血睾屏障和精子形成中的运输中起重要作用,Loveland 等研究表明一些重要的细胞因子在建立和维护睾丸的免疫特性方面有重要作用,如细胞因子SOCS 可通过jak 信号通路调节睾丸发育和免疫微环境建立,肿瘤坏死因子(TNF-a)、白细胞介素-1(IL-1)对睾丸自身免疫同样起重要作用。陈蕙芳对淫羊蕾的粗提取物进行研究,发现这些提取物中具有抗炎、雌激素样、抗衰老等多种生物活性,这与本研究得到的结果吻合。粘附连接是细胞和细胞直接的关键点,其在决定和维持组织结构方面起重要作用,在睾丸中,支持细胞之间以及支持细胞和生殖细胞之间的连接是由肌动蛋白连接的,这些连接对于精子的发生非常重要。Hippo 信号通路和jak-stat 信号通路在细胞分化中起关键作用,相关研究表明Hippo 信号通路对鲑鱼青春期睾丸支持细胞有调节作用,jak-stat 信号通路可以促进果蝇成年睾丸中的生殖干细胞分化。
4 结论
饲料日粮中添加淫羊藿可提高番鸭血清生殖激素中睾酮含量、雌二醇含量、促卵泡素含量和促黄体素表达,提高超氧化物歧化酶含量、谷胱甘肽过氧化物酶含量、总抗氧化水平。饲料日粮中添加淫羊藿对番鸭睾丸发育有显著影响,可激活白细胞和相关免疫受体表达,激活细胞因子受体相互作用、铁死亡等代谢途径和信号通路。本研究结果对饲料中添加淫羊藿以提高番鸭的繁殖力具有重要意义。
猜你喜欢
新民周刊(2022年27期)2022-08-01 07:04:49
中国临床医学影像杂志(2022年5期)2022-07-26 07:11:58
传染病信息(2021年6期)2021-02-12 01:52:58
中国生殖健康(2019年7期)2019-01-06 09:27:30
中成药(2017年4期)2017-05-17 06:09:20
中国男科学杂志(2016年9期)2016-03-20 15:00:13
医学研究杂志(2015年12期)2015-06-10 06:57:46
中国当代医药(2015年1期)2015-03-01 02:00:32
生物医学工程学进展(2015年1期)2015-02-28 14:53:42
化学工业与工程(2015年1期)2015-02-10 03:01:41