
过表达非洲猪瘟病毒D1133L蛋白的MA-104细胞系建立及其初步应用
2022-07-07 08:23崔卉梅袁兴国赵登率杨金柯陈学辉闫文倩申超超史喜绢张大俊刘湘涛郑海学张克山
畜牧兽医学报 2022年6期
张 婷,杨 博,崔卉梅,袁兴国,赵登率,杨金柯,郝 雨,陈学辉,闫文倩,申超超,史喜绢,张大俊,杨 行,刘湘涛,郑海学,张克山
(中国农业科学院兰州兽医研究所,兰州大学动物医学与生物安全学院 家畜疫病病原生物学国家重点实验室,兰州 730000)
非洲猪瘟(African swine fever, ASF)是由非洲猪瘟病毒(African swine fever virus,ASFV)感染家猪和野猪引起的一种严重的出血性传染病,ASF的流行传播对养猪业造成重大的经济损失,目前没有商业化的疫苗和抗病毒药物。ASFV感染猪的靶细胞主要为单核细胞和巨噬细胞,肺泡巨噬细胞(pulmonary macrophages, PAMs)用于ASFV的感染机制和病毒复制动力学的研究。目前,非洲猪瘟活病毒的检测依赖PAMs,但分离PAMs的过程复杂、耗时。源于非洲绿猴的商业化细胞系MA-104可用于替代PAMs感染ASFV,但其感染ASFV后病毒的复制能力并不高。
ASFV编码150~167个开放阅读框,ASFV编码包括50多种结构蛋白和100多种非结构蛋白,其中,ASFV编码包括5种解旋酶,分别为D1133L、Qp509L、Q706L、B962L和A859L,在病毒转录过程发挥作用,这些基因对病毒复制过程中的转录阶段发挥重要的功能,其中,1133L基因与牛痘病毒的D6R具有相似的结构域,与病毒转录起始阶段有关。
目前,常利用慢病毒(lentivirus)构建稳定表达细胞系来研究病毒的功能。如小反刍兽疫病毒(peste des petits ruminants virus, PPPRV)的结构蛋白N蛋白在Vero细胞系中稳定表达,在BHK细胞系稳定表达蓝舌病病毒(bluetongue virus,BTV)的VP6蛋白,猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)的GP2和N蛋白在MARC-145细胞系上稳定表达,稳定表达基因的BHK-21细胞系的构建及其对新城疫病毒(Newcastle disease virus, NDV)增殖效果的评价等。本实验室前期已明确了1133L的基因序列分析、蛋白结构预测及亚细胞定位。前期研究提示,ASFV编码的解旋酶D1133L对于病毒复制十分重要,为进一步研究1133L基因功能,本试验利用慢病毒系统,构建了过表达D1133L的MA-104细胞,并比较评价了病毒增殖能力。本文通过该方法构建过表达D1133L细胞系来探究此基因对ASFV复制能力的影响。
1 材料与方法
1.1 材料
1.1.1 细胞与病毒 MA-104细胞由兰州兽医研究所口蹄疫流行病实验室保存,非ASFV CN/GS/2018分离株、缺失MGF360-9L基因(ASFV-G-Δ40)和同时缺失ASFV-110-9L、ASFV-505-7R两个基因的 (ASFV-G-Δ20-53)重组毒株均由兰州兽医研究所非洲猪瘟区域实验室构建并保存。
1.1.2 试剂和抗体 胎牛血清(FBS)和0.25%EDTA-Trypsin(Gibco)购自伟博鑫生物科技有限公司,青霉素-链霉素-庆大霉素混合溶液(三抗)和PBS缓冲液购于北京索莱宝公司,大肠杆菌Trans5a感受态、LADNA聚合酶、限制性核酸内切酶Ⅰ和Ⅰ、T4 DNA 连接酶、均购于宝生物工程大连有限公司,RPIM1640培养基和高糖DMEM培养基购自购自Thermo Fisher Scientific公司,鼠抗β-actin单克隆抗体和TRizol Reagent购自英潍捷基(上海)贸易有限公司,HRP-conjugated Affinipure Goat Anti-Rabbit IgG(H+L),Goat Anti-Mouse,蛋白marker购自Proteintech公司,Western一抗稀释液、Western Bright ECL 化学发光底物和硝酸纤维素膜NC膜(0.22 μm)购于碧云天有限公司,预混型定量用反转录试剂盒和TB Green染料法标准型定量试剂盒购于上海百赛生物技术股份有限公司。
本研究所用的鼠抗ASFV p30单克隆抗体,兔抗ASFV p72多克隆抗体和鼠抗1133L单克隆抗体由中国农业科学院兰州兽医研究所口蹄疫与新发病流行病学团队提供。
1.2 方法
1.2.1 引物设计和质粒构建 根据NCBI数据库中ASFV1133L基因 (ID:59226965),以慢病毒表达载体pCDH-CMV-MCS-EF1-CopGFP-T2A-Puro进行构建,根据1133L的基因CDS序列和载体的MCS位点,设计特异性PCR primers,设计的引物序列见表1,双酶切酶为RⅠ和Ⅰ,酶切后重连插入到慢病毒载体 Lv-PCDH 的 MCS 区,命名为pCDH-CMV-D1133L-EF1-CopGFP-T2A-Puro。

表1 D1133L特异性PCR引物序列
1.2.2 ASFV病毒感染 ASFV毒株(MOI=1.5)感染MA-104/D1133L细胞系和野生型MA-104,48 h后收取病毒液。将MA-104/D1133L和MA-104细胞消化后铺于12孔板中,置于37 ℃、5%CO培养箱中,待细胞融合度至 70%~90%时,用无血清的DMEM 将细胞清洗2次,以去除细胞中残留的血清;用无血清的DMEM 将ASFV稀释至适当的浓度,之后将稀释好的毒液按照适当的体积加入到细胞中,置于37 ℃、5%CO培养箱孵育2 h 之后,弃去毒液,换成 2%FBS DMEM 维持液,继续培养。
1.2.3 RNA的提取及反转录 采用 Trizol 裂解法提取细胞的总 RNA,并反转录为 cDNA,用于Real-time qPCR,反转录体系 20 μL:4 μL 5×反转录酶,6 μL DEPC 水,10 μL模板 RNA。反应程序:37 ℃ 15 min,85 ℃ 5 s,4 ℃保存。
1.2.4 实时定量 PCR(quantitative real-time PCR) 实时荧光定量PCR(Real-time qPCR,RT-qPCR)的反应体系(10 μL):5 μL SYBR Permix ExⅡ,0.5 μL 上游引物,0.5 μL 下游引物,3 μL DEPC 水和1 μL cDNA。反应程序:95 ℃预变性 3 min;95 ℃变性 10 s,60 ℃退火和延伸 34 s,共 40 个循环,这些基因的相对mRNA水平归一化为猪mRNA水平,引物序列见表2。以ASFV72基因为靶点进行实时定量PCR,检测ASFV基因组DNA的拷贝数。采用QIAamp DNA Mini Kits(Qiagen,Germany)提取样品DNA。ASFV-72-R: 5′-CTGCTCATGGTATCAATCTTATCGA-3′;ASFV-72-F: 5′- GATACCACAAGATCAGCCGT-3;探针: 5′-CCACGGGAGGAATACCAACCCAGTG-3′;5′-CCACGGGAGGAATACCAACCCAGTG-3,扩增条件:95 ℃预热30 s, 95 ℃预热5 s,58 ℃预热30 s, 循环40次。用标准曲线计算ASFV基因组的数量,并以每微升基因组拷贝数表示。
1.2.5 Western blot 检测蛋白表达 MA-104/D1133L和MA-104细胞以每孔1×10细胞量接种于12孔板,感染ASFV(MOI=1.5)。不同时间点收取细胞裂解蛋白,用PBS洗涤后加入100 μL 1×loading蛋白上样缓冲液裂解细胞,取20 μL蛋白上样量进行SDS-PAGE,电泳结束后用100 V恒压转印至NC膜1.5 h,用TBST配制5%的脱脂奶粉室温封闭2.5 h,抗体稀释液稀释按照比列稀释一抗D1133L单克隆抗体(1∶1 000)、β-actin(1∶5 000)、p30(1∶1 000)和p72(1∶1 000),4 ℃过夜孵育,1×TBST配制HRP-conjugated Affinipure Goat Anti-Rabbit IgG(H+L),Goat Anti-Mouse(1∶10 000)室温孵育2 h,用Western Bright ECL 化学发光底物全自动化学发光成像分析。

表2 引物序列信息
1.2.6 ASFV病毒滴度测定 参考文献[19-20]方法来测定ASFV的HAD或TCID,利用ASFV荧光重组毒感染长满单层MA-104细胞和MA-104/D1133L细胞,36 h后收取,置于-80 ℃病毒反复冻融3次后,收集上清,1 200 r·min离心10 min, 取0.1 mL上清液进行10~10倍梯度稀释,接种长满PAM细胞的96孔细胞培养板,测定HAD,每孔加入25 μL的1%猪红细胞,每个稀释度接种8个孔,0.1 mL·孔,培养箱中37 ℃培养5~7 d,逐日观察记录细胞形成玫瑰花环数,而TCID根据出现的荧光数量并记录,用Reed-Munch 法计算。
1.2.7 数据分析 用GraphPad Prism 软件进行统计学分析并作图,采用独立样本检验进行显著性分析,*.<0.05 说明数据间有显著差异,**.<0.01 说明数据间有极显著差异。
2 结 果
2.1 MA-104/D1133L细胞系的构建
为构建1133L基因过表达MA-104细胞系,利用慢病毒表达质粒与病毒包装辅助质粒共转染MA-104细胞,转染48 h后。慢病毒Lv-PCDH、Lv-D1133L分别感染MA-104细胞,8 h后终止。加入3 μg·mL的puromycin对各组细胞系进行药筛培养,48 h后更换培养基并传代,维持培养,收集各组细胞系,提取各组细胞RNA,进行RT-qPCR验证,D1133L在该细胞系中能够高效、稳定表达。经测序后将获得的过表达D1133L稳定传代的细胞系命名为MA-104/D1133L。
2.2 MA-104/D1133L细胞系鉴定
利用慢病毒包装后Lv-PCDH、Lv-D1133L感染D1133L细胞后细胞形态均正常,药物筛选后能够正常的传代培养,将筛选出的阳性克隆细胞系连续传代15代后,用TRizol法提取细胞中的总RNA,反转录为cDNA,用RT-qPCR检测细胞系中的1133L的基因转录水平,Western blot鉴定细胞系中D1133L蛋白是否稳定表达。利用RT-qPCR检测了D1133L的转录水平表达(图1A),MA-104/D1133L 细胞系中的D1133L显著高于野生型MA-104(<0.05);Western blot结果(图1B)显示,细胞系连续传代15代后仍能稳定表达D1133L蛋白,以上表明该细胞系连续传代后仍能稳定转录和表达D1133L。RT-qPCR试验结束后,利用核酸电泳技术对RT-qPCR的产物进行检测,结果如图2所示:对于1133L基因,MA-104/D1133L细胞系的RT-qPCR扩增产物的条带与MA-104细胞系相比,差异显著;对于-基因,两个细胞系的RT-qPCR扩增产物的条带基本相当。
2.3 MA-104/D1133L细胞中ASFV基因表达检测
将MA-104/D1133L和MA-104细胞接种于12孔板,待细胞密度长至70%,ASFV(MOI=1.5)感染两种细胞,在12、24 h时收取的样品,利用RT-qPCR测定MA-104/D1133L细胞中30、72/mRNA表达量显著(<0.001)高于野生型MA-104细胞中30、72/mRNA表达量(图3A、3B)。即MA-104/D1133L细胞系与野生型MA-104细胞系相比在12、24 h均能促进ASFV30和72基因转录水平的表达。同时用Western blot检测p72蛋白表达,MA-104/D1133L细胞中p72蛋白表达量显著高于野生型MA-104细胞中p72表达量(图3C)。同样ASFV重组荧光毒株感染MA-104和MA-104/D1133L细胞,36 h后收取样品,检测p72和p30的表达量,与上述结果一致(图3D),MA-104/D1133L细胞系中p30和p72的高于野生型MA-104细胞中。综上所述,ASFV在MA-104/D1133L细胞中的复制能力强于MA-104。
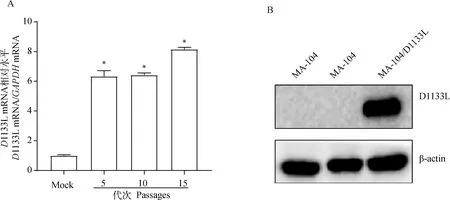
A. RT-qPCR鉴定D1133L基因转录水平;B. Western blot鉴定D1133L蛋白水平。与Mock相比,*.P<0.05A. Identification of D1133L gene transcription level by RT-qPCR; B. Western blot identification of D1133L protein level.Compared with Mock,*.P<0.05图1 连续传代至15代,MA-104/D1133L细胞系中D1133L稳定性鉴定Fig.1 Identification of D1133L stability in MA-104/D1133L cell line of successive generations to 15 generations
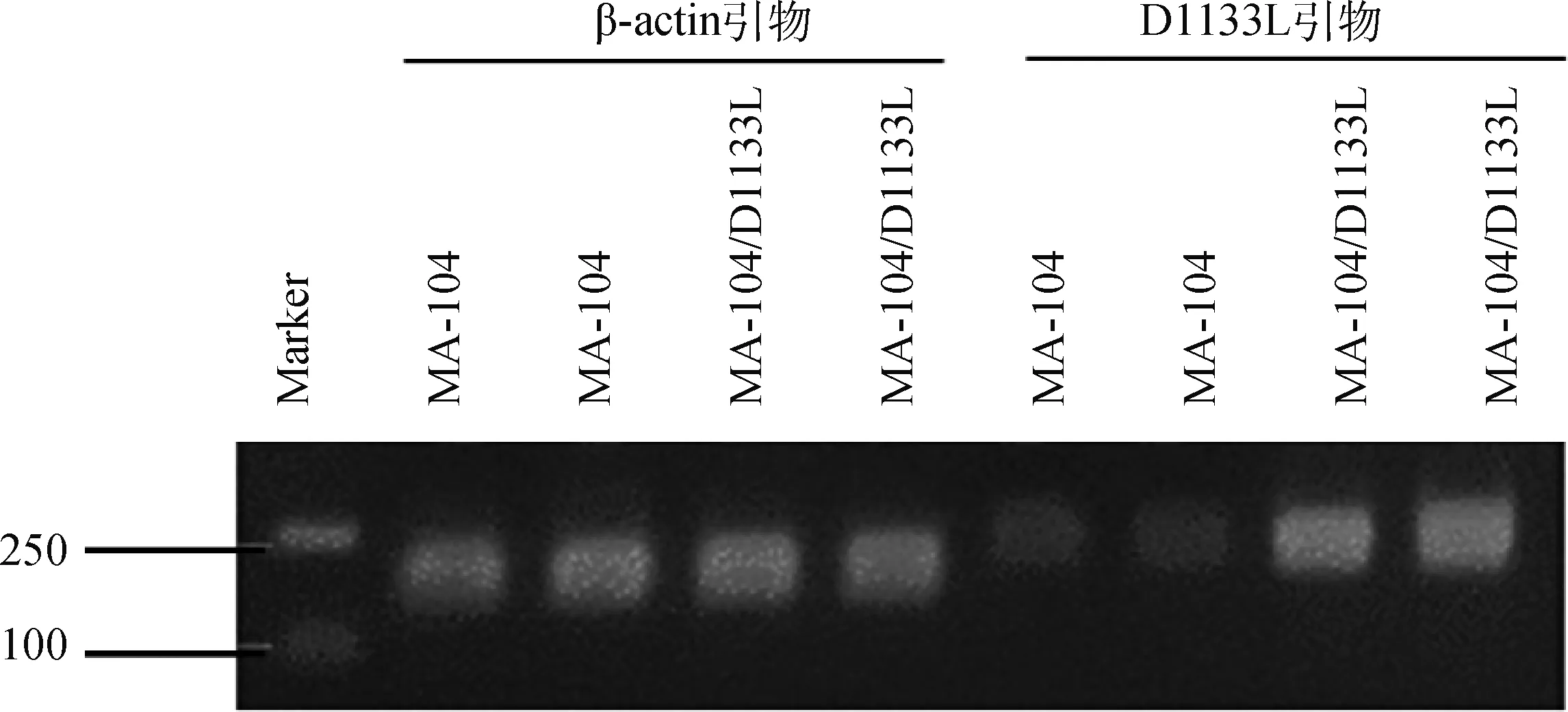
图2 MA-104和MA-104/D1133L中RT-qPCR扩增产物的电泳条带分析Fig.2 Electrophoretic band analysis of RT-QPCR products in MA-104 and MA-104/D1133L
2.4 MA-104/D1133L细胞中ASFV滴度及拷贝数测定
为进一步比较分析不同毒力毒株在MA-104/D1133L细胞中的增殖能力,ASFV强毒株ASFV CN/GS2018毒株和两种弱毒株ASFV重组荧光缺失毒株,分别为ASFV-G-Δ20-53和ASFV-G-Δ40株,在MA-104/D1133L细胞系和野生型MA-104细胞中的增殖水平,利用 RT-PCR 方法检测ASFV基因组,即病毒复制拷贝数。用HAD或TCID测定病毒滴度,在猪红细胞存在的情况下,CD2v蛋白介导ASFV在感染细胞周围的血液吸附,形成玫瑰花环,使用血液吸附试验(HAD)计算ASFV CN/GS2018毒株感染细胞后的病毒滴度,由于本试验构建的重组病毒均含有p72-eGFP启动子,可替代CD2v基因,用荧光代替花环来统计,因此每个样品的TCID是根据样品稀释后的eGFP荧光测定,统计荧光显微镜下荧光数量检测病毒滴度。
2.4.1 ASFV滴度(HAD/TCID)测定 如图4A所示,测定了强毒株ASFV CN/GS2018毒株HAD,即强毒株ASFV在MA-104/D1133L细胞系中的病毒滴度HAD(4.75和4.875)显著高于野生型MA-104细胞系病毒滴度HAD(4.125和4.25),同时也比较两种重组弱毒株在MA-104/D1133L细胞系和野生型MA-104细胞上的增殖能力,在双基因缺失毒株ASFV-G-Δ20-53中病毒滴度TCID,MA-104/D1133L细胞系病毒滴度TCID(3.50和4.00)比野生型MA-104细胞系增殖滴度TCID(2.875和2.875)ASFV的高(图4B),在单基因缺失毒株ASFV-G-Δ40中病毒滴度TCID,MA-104/D1133L细胞系病毒滴度TCID(3.25和3.50)比野生型MA-104细胞系增殖滴度TCID(2.625和2.750)ASFV的高(图4C)。综上结果表明MA-104/D1133L细胞系病毒增殖能力显著高于野生型MA-104细胞(<0.05)。
A. ASFV p30的mRNA水平;B. ASFV p72的mRNA水平;C. 感染ASFV CN/GS2018毒株不同时间点ASFV p72蛋白水平;D. 感染ASFV重组毒株36 h后ASFV p30、p72蛋白水平。*. P<0.05;**.P<0.01;***. P<0.001A. ASFV p30 mRNA level; B. ASFV p72 mRNA level; C. different time points of infection with wild strain ASFV p72 protein level; D. AFSV P30 and ASFV p72 protein levels 36 h after infection with recombinant strains.*. P<0.05; * * P<0.01; ***. P<0.001图3 MA-104、MA-104/D1133L细胞系感染ASFV后p30、p72转录和蛋白水平变化Fig.3 Changes in p30 and p72 transcription and protein levels of MA-104 and MA-104 /D1133L cell lines infected with ASFV
2.4.2 ASFV基因拷贝数测定 采用RT-qPCR方法测定不同毒力毒株中病毒复制拷贝数来评估病毒的复制差别。如图5所示,ASFV CN/GS2018毒株、双基因重组缺失病毒ASFV-G-Δ20-53和单基因缺失重组病毒复制拷贝数结果表明MA-104/D1133L的细胞系均显著高于比野生型MA-104病毒拷贝数(<0.05)。综上表明,ASFV毒株在MA-104/D1133L的复制能力均高于MA-104细胞。
3 讨 论
目前,一些商业化的传代细胞可感染ASFV。ZMAC-4 猪巨噬细胞系可替代PAM细胞研究其复制动力学,MA-104细胞可替代肺泡巨噬细胞检测ASFV。最新报道也发现ASFV 可以感染HEK293T细胞,但复制效率较低,有趣的是在HEK293T细胞中连续传代,获得了RT-qPCR测定MA-104/D1133L细胞中30、72/mRNA表达量显著(<0.001)高于野生型MA-104细胞中30、72/mRNA表达量(图3A、3B)。即MA-104/D1133L细胞系与野生型MA-104细胞系相比在12、24 h均能促进ASFV30和72基因转录水平的表达。同时用Western blot检测p72蛋白表达,MA-104/D1133L细胞中p72蛋白表达量显著高于野生型MA-104细胞中p72表达量(图3C)。同样ASFV重组荧光毒株感染MA-104和MA-104/D1133L细胞,36 h后收取样品,检测p72和p30的表达量,与上述结果一致(图3D),MA-104/D1133L细胞系中p30和p72的高于野生型MA-104细胞中。综上所述,ASFV在MA-104/D1133L细胞中的复制能力强于MA-104。
A. ASFV CN/GS2018毒株 (WT) HAD50测定;B. ASFV-G-Δ20-53重组毒株TCID50测定;C. ASFV-G-Δ40重组毒TCID50测定A. ASFV CN/GS2018 HAD50 assay; B. TCID50 assay of ASFV-G-Δ20-53; C. TCID50 determination of ASFV-G-Δ40图4 不同ASFV毒株感染MA-104、MA-104/D1133L细胞后病毒滴度测定Fig.4 Titer determination of different ASFV strain infected MA-104 and MA-104/ D1133L cells
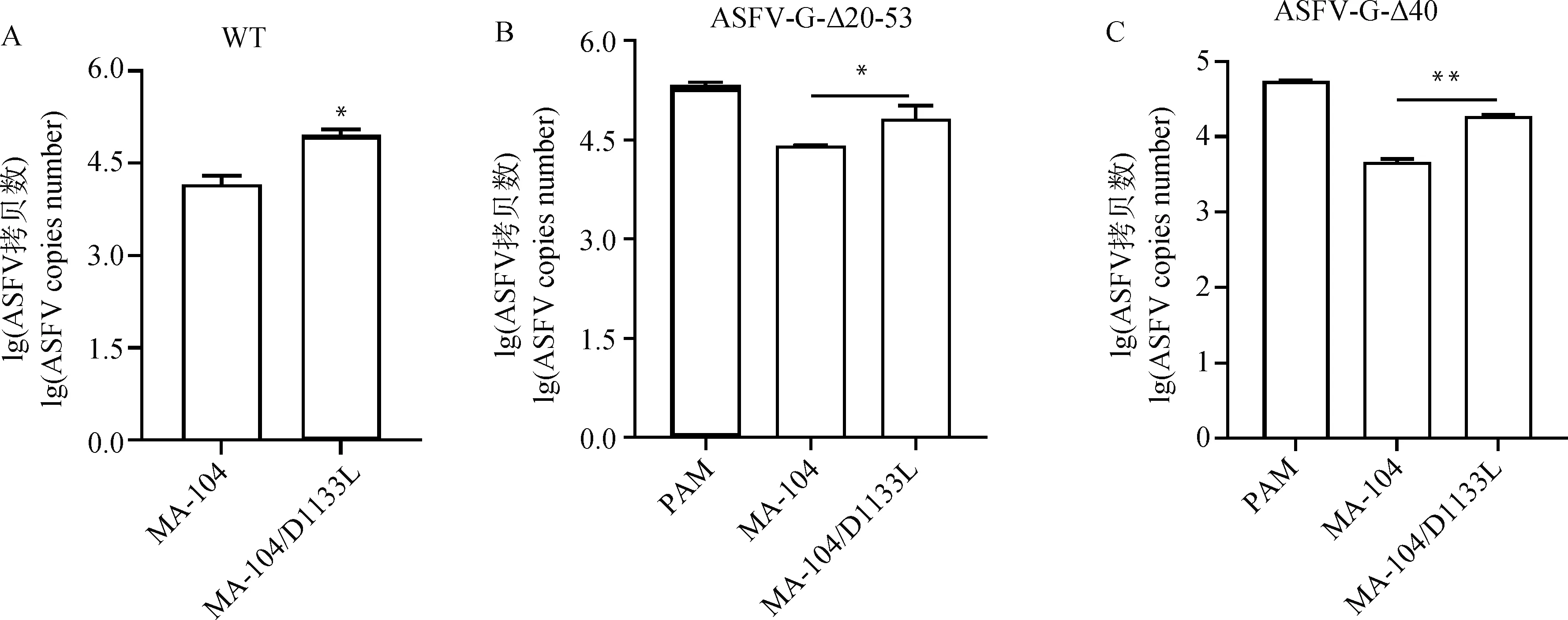
A. ASFV CN/GS2018毒株 (WT) 病毒拷贝数;B. ASFV-G-Δ20-53重组毒株病毒拷贝数;C. ASFV-G-Δ40重组毒株病毒拷贝数A. CN/GS 2018 virulent strain (WT) virus copies number; B. Copies number of ASFV-G-Δ20-53 recombinant virus; C. Copies number of ASFV-G-Δ40 recombinant virus图5 评估不同毒株感染MA-104、MA-104/D1133L细胞系的病毒拷贝数Fig.5 Evaluation of the copies number of different virus strains infected MA-104 and MA-104/D1133L cell lines
ASFV编码1133L基因与病毒复制相关,其对病毒复制具有重要作用。利用D1133L蛋白解旋酶基序的特性,探究D1133L在病毒复制中发挥的作用是具有重要意义。本试验构建稳定表达D1133L的MA-104/D1133L细胞系,评估了ASFV CN/GS2018分离株和ASFV基因缺失重组毒株在MA-104/D1133L细胞系的病毒复制水平和增殖能力,通过RT-qPCR和Western blot技术检测了MA-104、MA-104/D1133L细胞系感染ASFV后,早期蛋白p30和晚期蛋白p72转录和蛋白水平之间的差异,测定病毒滴度HAD或TCID和病毒拷贝数在两种细胞中ASFV的增殖水平和病毒复制,从数据可知ASFV在MA-104/D1133L细胞增殖能力高于MA-104。综上所述,ASFV感染该细胞系后病毒增殖能力高于野生型MA-104,过表达D1133L蛋白能进一步促进ASFV复制。
4 结 论
D1133L在ASFV复制过程中扮演重要角色,本试验通过慢病毒表达系统,成功构建过表达D1133L的MA-104/D1133L细胞系,不同毒力的ASFV毒株感染此细胞系,ASFV增殖能力强于野生型MA-104,本研究构建的D1133L过表达MA-104细胞系为研究1133L基因功能及针对该基因有效药物的开发积累了生物材料。
猜你喜欢
中国现代医生(2022年21期)2022-08-22
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
环球时报(2021-07-12)2021-07-12
广东教育·职教版(2021年3期)2021-04-20
奥秘(2019年8期)2019-08-28
学校教育研究(2018年27期)2018-05-14
小猕猴学习画刊(2017年3期)2017-07-19
小猕猴智力画刊(2016年6期)2016-05-14
科学中国人(2016年1期)2016-01-13
