
我国南方地区桉树叶浸泡对水库沉积物致黑元素分布及迁移转化的影响*
2022-06-30 07:06李一平刘斯璇李荣辉唐春燕郭晋川黄亚男王亚宁
湖泊科学 2022年4期
魏 尧,李一平,朱 雅,刘斯璇,李荣辉,唐春燕,郭晋川,陈 刚,黄亚男,王亚宁,陈 春,何 菁
(1:河海大学环境学院,浅水湖泊综合治理与资源开发教育部重点实验室,南京 210098)(2:广西壮族自治区水利科学研究院,南宁 530023)(3:河海大学,水文水资源与水利工程科学国家重点实验室,南京 210098)(4:南京市水利建筑工程检测中心有限公司,南京 210098)
桉树原产于澳洲大陆,自18世纪末被首次发现和命名后,被迅速引种到世界各地[1-2].目前,桉树已在世界近130个国家和地区大规模引种,其种植面积已超过全球人工林种植面积的1/3,桉树人工林已成为世界人工林的重要组成部分[3].桉树是短轮伐期树种,一次造林采伐2~3次,桉树砍伐后大量新鲜剩余物(根、茎和叶)会被留在林区,随地表径流冲刷进入地表水体,直接影响水体水质.桉树种植区水体泛黑问题在国内外普遍存在,主要发生在河道、洼地、池塘和水库等水体[4-6].如2011年澳大利亚墨累达令河流域发生的严重泛黑事件造成了大量鱼虾死亡,其影响的河道长度超过2000 km,持续时间长达6个月[7-8].我国桉树人工林区洼地黑水主要源于桉树新鲜叶和嫩梢[9],池塘黑水暴发现象主要出现在高温高湿多雨天气[10],许多桉树人工林区水库出现不同程度的冬季突发性水体泛黑现象[11-12].我国桉树人工林区水库水体泛黑的最新研究成果表明,水体泛黑主要是由铁、锰、硫化物与溶解性有机物(单宁酸)等物质之间发生一系列的反应引起的[13].李一平等[14-15]认为缺氧/厌氧环境下产生的FeS、MnS沉淀是主要的致黑因素,杨钙仁等[16]研究发现淋溶作用下桉树残体富含黑色DOC的浸出液随暴雨冲刷进入库底可能导致泛黑,罗凡等[17-18]发现桉树浸出的DOC、单宁酸与Fe2+在有氧条件下可络合致黑,朱雅等[11]的进一步研究认为Fe和DOC是致黑的主要贡献物质.桉树人工林区水库黑水组成和来源十分复杂,水库水体水动力条件复杂,使得黑水具有滞后效应,导致目前研究对于三种可能致黑物质对水库泛黑的贡献程度和途径仍不清楚,关键的具体致黑物质有待甄别.且现有研究主要集中于水质氮磷等元素指标,并未深入研究沉积物内源释放的影响.
基于此,本研究选择我国南方地区广西壮族自治区南宁市天雹水库作为研究对象,选取夏季水体未泛黑期(水体热分层稳定)进行室内浸泡实验,通过对比桉树叶浸泡与第二大树种——马尾松叶浸泡对上覆水与沉积物性质的影响,利用多参数水质监测仪(YSI)测定每6 h上覆水体溶解氧(DO)、氧化还原电位(Eh)、透光度等理化指标,利用高分辨率原位被动采样技术(ZrO-Chelex-AgI DGT和HR-Peeper)捕捉沉积物中DGT有效态铁、锰、硫和DOC、单宁酸浓度分布,探究水库水体突发性泛黑原因,甄别致黑物质,为研究我国南方地区桉树人工林区水库泛黑机制提供一定的科学依据.
1 材料与方法
1.1 研究区域概况
天雹水库(22°52′~22°53′N,108°13′~108°14′E)位于广西壮族自治区南宁市西郊心圩镇,临近高新区.该水库面积约73.3万m2,集雨面积约50.8 km2,最大水深约18 m,总库容1360万m3,有效库容880万m3.水库入库水量主要来源于降雨,水库集雨区内桉树种植面积达72%,砍伐历史超过10年.南宁属于亚热带季风气候区,夏季高温多雨,冬季温暖干燥,5-9月降雨集中,年均降雨量1304.2 mm.南宁历年8月平均气温28℃,降水量178 mm,相对湿度78%.天雹水库沉积物中铁和锰含量丰富(以S3采样点为例,铁、锰含量分别可达34500~64200和650~1870 mg/kg)[11],且水温分层现象明显.
1.2 样品采集
于2020年8月进行采样,并综合考虑库区地形、水动力、周边桉树种植情况等因素,选取3个代表性采样点,其中S1代表浅水区(水深约为3.0 m),S2代表过渡区(水深约为10.7 m),S3代表深水区(水深约15.2 m)(图1).利用柱状沉积物采样器(中国,EasySensor)于每个采样点分别原位采集4管沉积物样柱(沉积物样柱直径为11 cm,高度均大于20 cm),采用原位水进行培养(上覆水水柱高约25 cm),采集后立即用保鲜膜进行封口,并用锡箔纸进行避光处理,后运送回实验室(南宁),静置24 h.同时利用多参数水质监测仪(美国,EXO2)实地监测水体垂向溶解氧(DO)、氧化还原电位(Eh)等水体理化指标.

图1 南宁市天雹水库及采样点位置
1.3 样品分析
本研究选择典型泛黑水库天雹水库周围采集新鲜的桉树叶与马尾松叶作为浸泡对象.将收集好的原始样品挑除杂质,剪碎成1~2 cm,并分别等量分成3份(每份5 g).待沉积物静置后,将处理好的树叶加入采集的沉积物样柱中浸泡54 h.实验共设置4组沉积物样柱处理,分别为缺氧空白组(编号为S1Q、S2Q、S3Q)、有氧空白组(编号为S1X、S2X、S3X)、有氧桉树组(编号为S1A、S2A、S3A)和有氧马尾松组(编号为S1M、S2M、S3M).缺氧组表示样柱始终用保鲜膜封口,浸泡过程中无氧气加入;有氧组表示样柱为敞口,与空气充分接触.在浸泡过程中,每6 h用多参数水质监测仪(美国,EXO2)测量水体溶解氧浓度(DO)和氧化还原电位(Eh),同时用LISST-200X激光粒度仪(美国,Sequoia)测量水体透光度(为了保持缺氧处理,没有连续监测缺氧空白组的水体理化指标).
在沉积物样柱浸泡6 h时,将HR-Peeper(中国,EasySensor)(每个样柱插入两个HR-Peeper,取样体积为400 μL)垂直插入沉积物中,在其浸泡30 h时将ZrO-Chelex-AgI DGT(中国,EasySensor)也插入相同的沉积物中,待DGT放置24 h,将Peeper与DGT同时取出.用去离子水快速冲洗DGT,标记沉积物-水界面后,装入湿润的自封袋保存.用纸巾将Peeper表层底泥擦除,并用移液枪将Peeper间隙水样转移至离心管(规格为1.5 mL),密封冷藏保存待分析.用陶瓷切片刀将DGT固定膜按垂向方向切成条状(宽度1~5 mm,长度20 mm),后将所有条状固定膜依次放到离心管中,加入1 mol/L HNO3,确保固定膜完全浸没,室温静置提取16 h以上,取出固定膜,保存提取液待测定,分别采用比色分析方法、ICP-MS、计算机密度成像计量(CID)技术测定DGT有效态Fe、Mn、S浓度[19-20],其中DGT有效态浓度是指测量期间内DGT与沉积物界面的平均浓度[21].对于间隙水样品中的检测指标,分别采用顺序注射-紫外线在线氧化分光光度法[22]和分光光度法(NY/T 1600-2008)测定溶解性有机碳(DOC)和单宁酸含量.
1.4 数据处理
为了反映沉积物-水界面的扩散方向,本研究采用表观扩散通量(apparent diffusion flux,J)的概念,其计算公式如下[23]:
(1)
式中,Jw和Js分别表示离子从上覆水到沉积物-水界面和离子从沉积物到沉积物-水界面的通量(mg/(m2·d)),(∂CDGT/∂xw)x=0和(∂CDGT/∂xs)x=0分别表示上覆水和沉积物中各离子DGT浓度梯度,本文选用垂向深度-30~10 mm范围进行浓度梯度计算,Dw和Ds分别表示离子在上覆水和沉积物中的扩散系数.本研究D0(Fe2+)、D0(Mn2+)和D0(S2-)分别取7.19×10-6(25℃)、6.88×10-6(25℃)和6.95×10-6cm2/s(18℃)[24].φ表示沉积物孔隙度,计算公式如下:
φ=Wds/[(1-W)dw+Wds]
(2)
式中,W表示沉积物含水量(%),ds和dw分别表示沉积物和上覆水的平均密度(g/cm3).
2 结果与分析
2.1 树叶浸泡对水体理化性质的影响
桉树叶浸泡与马尾松叶浸泡均消耗水体中的DO.S1、S2、S3点位样柱水体的初始DO浓度平均值为5.57、4.48、5.35 mg/L.有氧桉树组S1A、S2A、S3A在浸泡54 h后分别消耗4.18、1.78、3.51 mg/L DO,该消耗量分别是其原浓度的68.86%、43.73%、60.73%.而有氧马尾松组S1M、S2M、S3M在浸泡54 h后分别消耗2.91、1.79、3.08 mg/L的DO,分别是其原浓度的50.61%、38.09%、56.41%.两者消耗量分别相差1.27、0.01、0.43 mg/L的DO,分别是初始平均浓度的22.80%、0.22%、8.03%.由此可见,不存在水温分层的S1点位在加入2种新鲜叶浸泡后消耗的DO浓度之差大于存在水温分层的S2和S3点位.值得注意的是,S2点位的初始DO浓度低于其它2个点位,这可能是因为S2点位水深较深,在水体热分层的条件下,底部处于厌氧环境.此外S2点靠近桉树种植区,由于砍伐与降雨径流双重影响,底部可能有桉树叶淤积,导致微生物耗氧及内源释放严重.而S1点位水深较浅,能直接获取大气复氧补给;S3点位于库尾坝前,虽然水深较深,但由于联合出水,故有DO补给(图2).
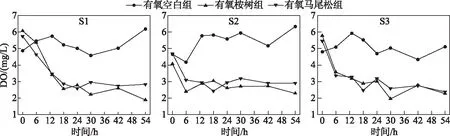
图2 树叶浸泡下水体DO浓度随时间的变化
有氧桉树组Eh值总体呈现先降后升的趋势,而有氧马尾松组Eh值总体呈现下降趋势(图3).S1、S2、S3点位样柱水体的初始Eh平均值为413.1、307.5、287.0 mV.有氧桉树组S1A、S2A、S3A分别在浸泡24、18、30 h后Eh达到最小值66.7、23.0、-33.2 mV,而有氧马尾松组S1M、S2M、S3M在浸泡42 h后Eh分别达到最小值98.1、47.3、72.5 mV.因此,桉树叶浸泡后对水体氧化还原电位的影响显著大于马尾松叶浸泡后对水体氧化还原电位的影响,且相比不存在水温分层的S1点,存在水温分层的S2和S3点在桉树浸泡后的还原作用更强,这可能是因为S2、S3点的水温分层使其底层水体成为缺氧环境[11],从而致使沉积物中的还原性物质(如Fe2+、Mn2+等)释放.
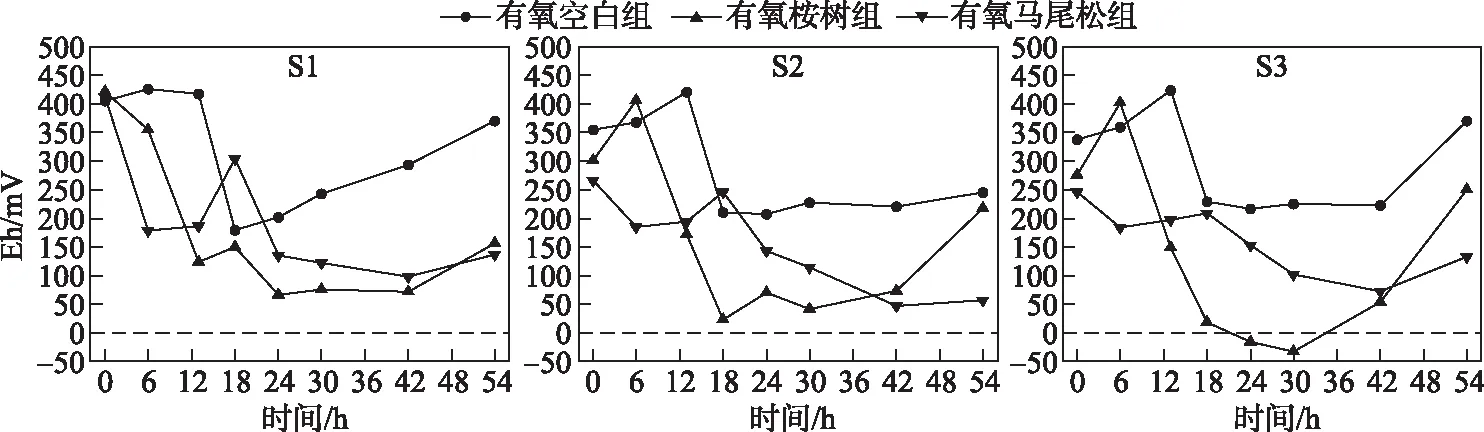
图3 树叶浸泡下水体Eh随时间的变化
水体透光度主要受颗粒粒径的影响.S1、S2、S3点位样柱水体的初始透光度平均值分别为 73.17%、49.48%、56.08%,有氧桉树组S1A、S2A、S3A在浸泡54 h后透光度分别下降65.01%、49.25%、61.79%,而有氧马尾松组S1M、S2M、S3M在浸泡54 h后透光度分别下降27.53%、31.72%、9.36%(图4).因此,桉树叶浸泡后对水体透光度的影响显著大于有氧马尾松叶组.可以推测,桉树叶浸泡透光度下降是由于黑色颗粒的形成,而马尾松则是由于松油的分泌(其样柱表层可见明显松油光泽).
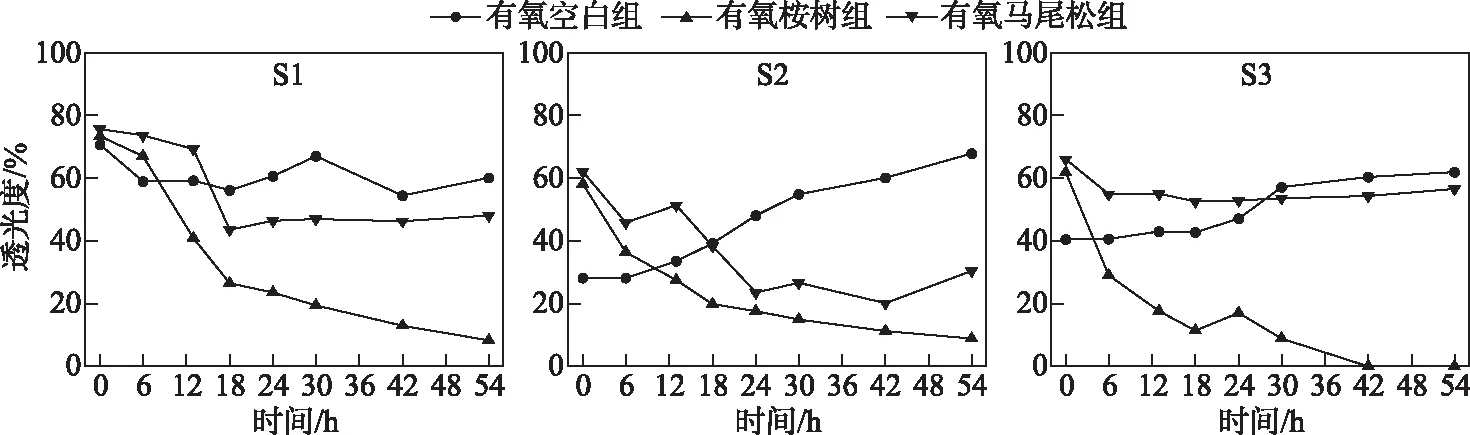
图4 树叶浸泡下水体透光度随时间的变化
2.2 树叶浸泡对水体DOC和单宁酸浓度的影响
桉树叶浸泡和马尾松叶浸泡均在水体中释放大量有机质,导致上覆水中DOC含量突增.有氧桉树组S1A、S2A、S3A上覆水中DOC含量分别较有氧空白组的12.5%、12.5%、5.9%突增至96.2%、85.1%、65.1%(表1).有氧马尾松组中S1M和S2M上覆水DOC含量较有氧空白组略有增加,而S3M变化趋势与S3A变化趋势基本一致,上覆水中DOC含量显著上升.
缺氧空白组S1Q、S2Q、S3Q沉积物单宁酸浓度分别为9.03、12.31和4.68 mg/L(表1),说明天雹水库表层沉积物中富含单宁酸,且S2采样点的单宁酸浓度最高,再次验证了陆域桉树残体输入是天雹水库沉积物单宁酸的主要来源.有氧桉树组S1A、S2A、S3A上覆水中单宁酸浓度分别突增至23.9、26.0和34.0 mg/L,且其浓度显著高于沉积物中单宁酸浓度.有氧马尾松组S1M、S2M、S3M中单宁酸浓度均较低,其上覆水浓度分别为4.29、4.75、1.25 mg/L,均显著小于空白组单宁酸浓度.

表1 树叶浸泡下上覆水和沉积物中DOC和单宁酸浓度分布
2.3 树叶浸泡对沉积物Fe、Mn、S分布的影响
空白组沉积物中的DGT有效态Fe浓度均随深度的增加而增加,缺氧空白组S1Q、S2Q、S3Q浓度范围分别为4.77~21.92、9.11~28.85和8.42~38.78 mg/L,有氧空白组S1X、S2X、S3X浓度范围分别为1.70~16.70、4.46~20.75和1.41~28.29 mg/L,有氧空白组DGT有效态Fe浓度整体低于缺氧空白组.有氧桉树组S1A、S2A、S3A中DGT有效态Fe浓度在深度上总体呈先升后降的趋势,分别在-20、0、-10 mm处达到峰值18.37、15.30、18.84 mg/L.有氧马尾松组中S1M和S2M的DGT有效态Fe浓度总体随深度的增加呈上升趋势,浓度范围分别为3.48~24.17、11.71~26.43 mg/L,相较而言,S3M的DGT有效态Fe浓度变化波动较大,浓度范围为4.98~19.87 mg/L(图5).
缺氧空白组S1Q、S2Q、S3Q的DGT有效态Mn浓度均随深度的增加而增加,浓度范围分别为0.22~0.99、0.49~1.76、0.72~2.05 mg/L(图5).有氧桉树组S1A、S2A、S3A上覆水中的DGT有效态Mn浓度相较于缺氧空白组分别增加至0.68、1.18、0.98 mg/L,说明沉积物中不溶性的Mn(Ⅳ)还原为溶解性Mn(Ⅱ),释放到上覆水中.
缺氧空白组S1Q、S2Q、S3Q沉积物中的DGT有效态S浓度较低,范围分别为0.024~0.035、0.020~0.026和0.023~0.029 mg/L(图5),且加入桉树叶与马尾松叶后,其沉积物中的DGT有效态S浓度较缺氧空白组均有所下降.
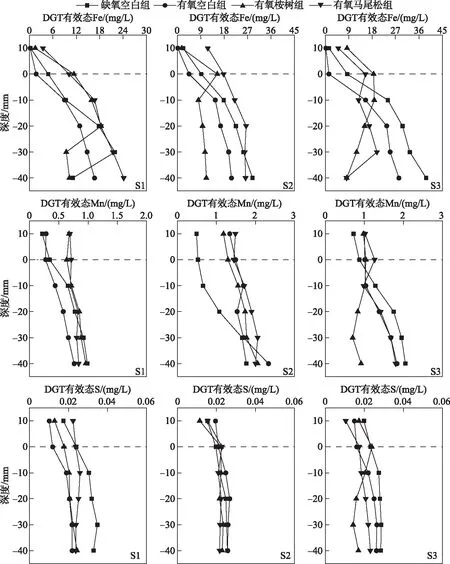
图5 树叶浸泡下沉积物中DGT有效态Fe、Mn、S浓度的垂向分布
2.4 沉积物-水界面处Fe2+、Mn2+、S2-表观扩散通量
负值和正值分别代表到沉积物和水的通量[23].本次浸泡过程中Fe2+、Mn2+、S2-的扩散通量范围分别为12.416~31.806、0.111~1.807、0.003~0.023 mg/(m2·d),均为正值,表明沉积物是各金属离子通量的源(图6).有氧桉树组S1A、S2A、S3A的各金属扩散通量(特别是Mn2+)均低于缺氧空白组S1Q、S2Q、S3Q的各金属扩散通量,因此可以推测,在桉树叶浸泡过程中沉积物中不溶性的Mn(Ⅳ)还原为溶解性Mn(Ⅱ),不溶性的Fe(Ⅲ)还原为可溶性Fe(Ⅱ),并与还原态硫反应,生成硫化亚铁(FeS)和硫化锰(MnS)沉淀[25].此外,S3A的Fe2+扩散通量明显高于S1A、S2A,这可能是由于S3A的强还原性条件(Eh<0 mV)能够促进更多的Fe2+进入上覆水中[26].各样柱S2-通量值较小,反映沉积物和上覆水之间的S扩散几乎达到平衡.
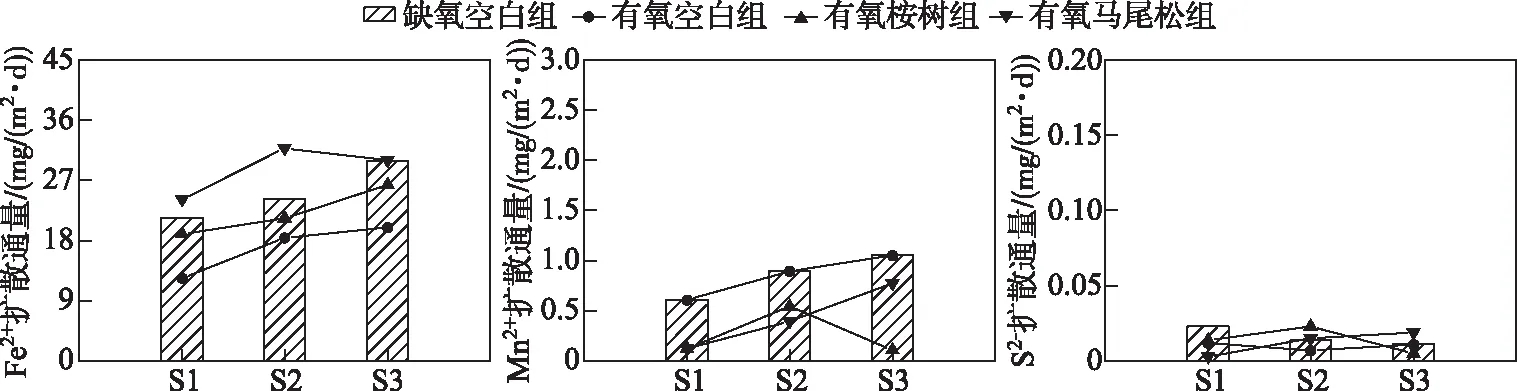
图6 沉积物-水界面处Fe2+、Mn2+、S2-扩散通量
3 讨论
树叶在水体中的分解过程可以概括为两个阶段:第一阶段是物理作用,易溶物质的快速溶解,第二阶段是分解作用,树叶所含物质的慢速分解[27].本实验时长小于3 d,故以树叶的快速淋溶作用为主.本次浸泡实验发现,在样柱中加入桉树叶浸泡1 h后,未与桉树叶接触的沉积物-水界面处水体色度开始明显变化,最终在浸泡24 h后样柱水体完全不透光(图7),这与罗凡等[17]的浸泡实验结果一致.此外罗凡等[15]的研究表明,在有氧条件下,桉树叶浸泡于水库底层水中后水体呈现黄棕色,并未出现黑色.由此可推测黄棕色是由桉树叶浸泡释放出的高浓度有机质和单宁酸导致的,而黑水的形成则是高浓度有机质、单宁酸与沉积物向上覆水体中释放铁、锰等金属离子共同反应的结果.

图7 树叶浸泡54 h后的水体颜色对比
为了进一步论证,本研究对桉树叶和马尾松叶浸泡过程理化指标进行相关性分析.分析发现,有氧桉树组(S1A、S2A、S3A)的透光度均与DO浓度存在显著正相关性(R=0.978、0.767、0.947)(表2),说明黑水形成过程需要消耗大量DO.此外,S1A透光度还与Eh值存在显著相关性,而S2A和S3A相关性较弱,这可能是由于S2和S3存在热分层现象,沉积物间隙水中Eh较低,对水体Eh值的影响较大,导致桉树叶浸泡对水体Eh值的影响相对较弱.
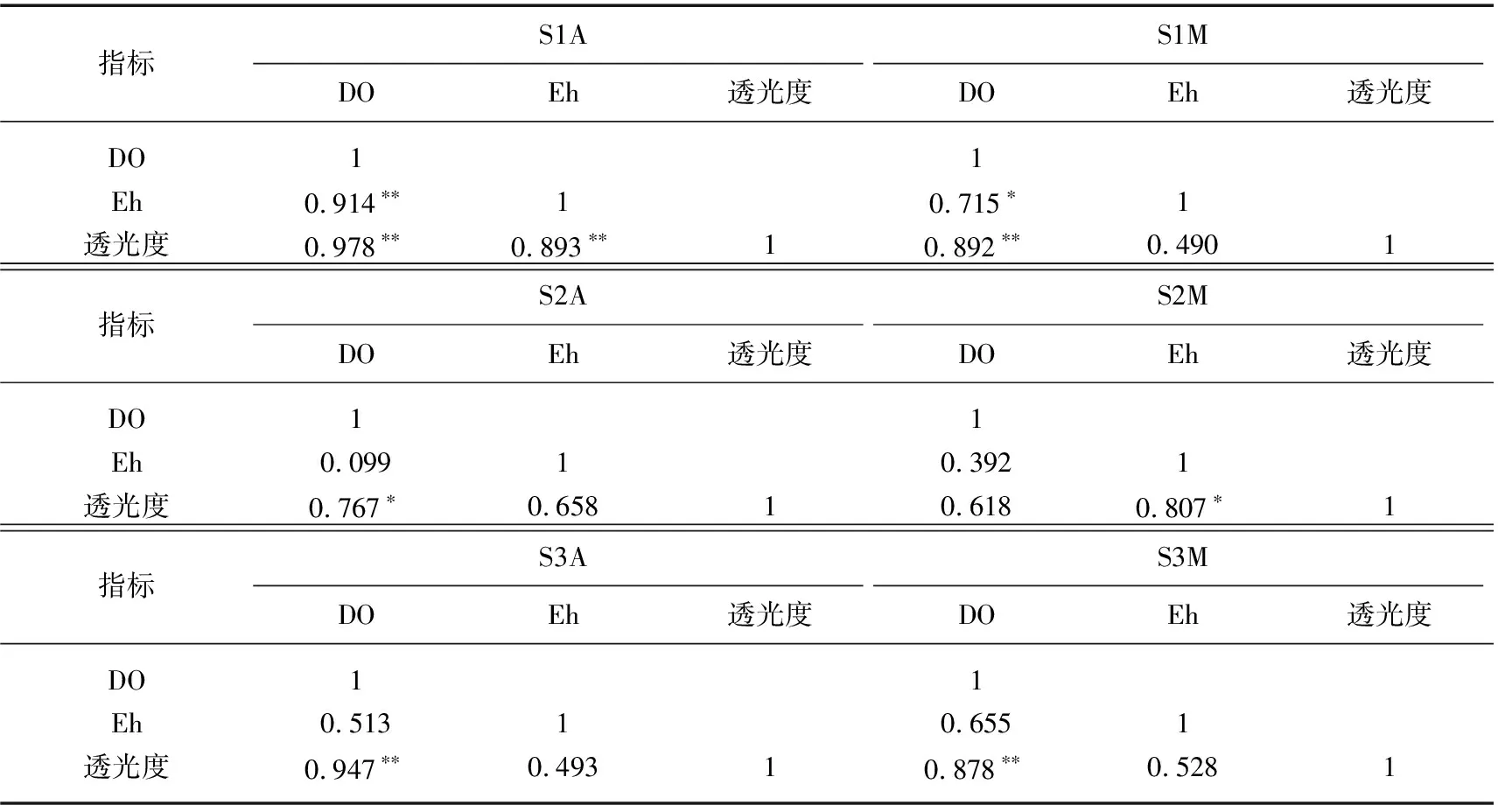
表2 沉积物上覆水DO、Eh和透光度的相关性分析
铁、锰和硫是沉积物氧化-还原体系的敏感元素[28],桉树人工林区水库沉积物中铁和锰含量丰富,其中铁含量约为太湖沉积物的1.6倍[11, 29].桉树叶浸泡和马尾松叶浸泡后上覆水中DGT有效态Fe浓度均明显增加,但马尾松叶浸泡后沉积物间隙水中DGT有效态Fe浓度波动无明显规律,而桉树叶浸泡后表层沉积物的DGT有效态Fe浓度明显增加,深层沉积物间隙水中DGT有效态Fe浓度明显下降,说明深层沉积物中Fe向沉积物-水界面迁移,为黑水形成提供了重要的Fe来源.此外,桉树叶浸泡和马尾松叶浸泡均在水体中释放大量有机质,导致上覆水中DOC含量突增.而桉树叶浸泡后释放出大量单宁酸,其上覆水中浓度分别达到23.9、26.0和34.0 mg/L,分别是马尾松叶浸泡后的5.6、5.5、27.2倍.可以推测,热分层期库底沉积物处于厌氧环境,沉积物中不溶性的Mn(Ⅳ)还原为溶解性Mn(Ⅱ),不溶性的Fe(Ⅲ)还原为可溶性Fe(Ⅱ),硫酸盐还原产生还原态硫,并进一步与还原态铁反应生成硫化亚铁(FeS)沉淀[25].有氧环境下,Fe2+在沉积物-水界面附近被氧化为Fe3+,一旦接触DOC和单宁酸,立即络合生成黑色沉淀,随着浸泡时间增长,DOC和单宁酸浓度不断上升,越来越多的Fe2+被络合而消耗,黑水现象也愈发严重.浸泡实验样柱中可能发生的反应如下(图8):
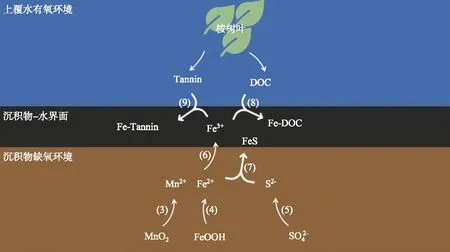
图8 桉树叶浸泡下样柱沉积物-水界面处物质反应
236MnO2+(CH2O)106(NH3)16(H3PO4)+472H+→ 106CO2+236Mn2++8N2+H3PO4+366H2O
(3)
424FeOOH+(CH2O)106(NH3)16(H3PO4)+848H+→ 106CO2+424Fe2++H3PO4+16NH3+742H2O
(4)
(5)
4Fe2++O2+4H+→ 4Fe3++2H2O
(6)
Fe2++S2-→ FeS
(7)
Fe3++DOC → Fe-DOC
(8)
Fe3++Tannin → Fe-Tannin
(9)
天雹水库桉树人工林区砍伐期一般在秋冬季.砍伐期内,大量桉树落叶携带着富含铁锰的土壤颗粒在降雨的作用下随径流进入库区水体[12],导致水库底部表层沉积物中的铁锰含量进一步增加[11].已有研究表明,相较于其它树种(如杂交相思、马尾松),桉树的单宁酸含量高且易于分解[9].在桉树叶浸泡分解的过程中,其释放的单宁酸与底层沉积物中的铁锰离子反应,产生单宁酸铁等致黑物质[30].但由于夏季水库水体处于分层期,温跃层阻止上下水体交换,导致致黑物质处于恒温层,无法进入上覆水体[11, 31].秋冬季,水体分层结构被破坏,温跃层消失,上下水体开始混合,底层致黑物质向上覆水移动,泛黑现象发生.这种由气象因子变化导致水体热分层结构失稳,影响上下水体交换与沉积物释放,进而导致水体泛黑的现象在红枫湖和百花湖也有发现,与桉树人工林区泛黑成因不同的是,其致黑物质为金属硫化物[32-34].
未来可进一步研究气象水文条件驱动下桉树人工林区水体泛黑过程的变化机理,为指导我国桉树人工林科学种植及保障水库生态环境安全提供重要科学支撑.
4 结论
1)桉树叶浸泡明显消耗水体中的DO,且显著影响水体Eh与透光度.此外,透光度与DO之间呈显著正相关(相关系数为0.618~0.978).
2)桉树叶浸泡释放大量DOC和单宁酸,其中单宁酸浓度远超有氧马尾松组,其上覆水中浓度分别达到23.9、26.0和34.0 mg/L,说明桉树浸泡为黑水形成提供了重要的DOC和单宁酸来源.
3)桉树叶浸泡后上覆水和表层沉积物中DGT有效态Fe浓度均明显增加,深层沉积物间隙水中DGT有效态Fe浓度明显下降,且其表观扩散通量均为正值,说明深层沉积物中Fe向沉积物-水界面迁移,为黑水形成提供了重要的Fe来源.
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
中国海洋大学学报(自然科学版)(2022年10期)2022-09-30
中国调味品(2022年8期)2022-08-05
环境工程技术学报(2022年3期)2022-06-05
浙江农业科学(2022年4期)2022-03-31
新农业(2021年12期)2021-11-29
养猪(2021年1期)2021-11-26
农家科技中旬版(2020年10期)2020-07-12
三峡大学学报(自然科学版)(2020年4期)2020-07-09
中国饲料(2020年3期)2020-03-09
