
辽宁仙人洞国家级自然保护区森林群落稳定性评价
2022-06-28 06:41:40宋文璐杜维新苑知言赵海涵
浙江农林大学学报 2022年3期
宋文璐,张 华,伏 捷,张 俊,杜维新,苑知言,赵海涵
(1. 辽宁师范大学 地理科学学院,辽宁 大连 116029;2. 辽宁师范大学 海洋可持续发展研究院,辽宁 大连116029)
森林是陆地生态系统的主体,具有涵养水源、保育土壤、固碳制氧、保护生物多样性等多种功能[1],森林生态系统的稳定性程度直接关系到其生态服务功能的发挥。森林生态系统稳定性评价是森林生态系统研究的重要领域。稳定性是植物群落结构与功能的综合特征[2]。自群落稳定性概念首次被提出后,至今已有70多种描述生态系统不同性质的稳定性概念,其内涵可归结为系统对外界干扰的抵抗力(resistance)和受扰动后返回初始状态的恢复力(resilience)等2个方面[3-4]。稳定性维持机制同样引起了国内外学者的深入探讨,已有理论中较完善的有多样性或复杂性理论、食物网理论、冗余理论等[5]。由于森林群落结构和功能的复杂性以及稳定性与研究区域的气候、种源和干扰等外界因素有关[2],所以至今尚没有统一的稳定性测度方法。学者们根据研究对象所选取的测度方法和指标体系不尽相同。如郑元润[6]在对大青沟森林植物群落的研究中,提出了群落演替与比较分析相结合的方法,并对Godron稳定性测定方法作出改进,近年的稳定性研究中也广泛运用到改进后的Godron法[7-8]。除此之外,许多研究利用野外调查和实验数据,包括群落组成、更新潜力、物种多样性、土壤理化性质、人为干扰等,建立稳定性评价指标体系;运用主成分分析法[9]、模糊数学中隶属函数值法[10-11]、综合指数法[12]等对森林群落稳定性进行评价。生态系统结构决定其功能。有研究证明:林分空间结构指标也可以较好地反映森林群落的稳定性[13]。综上所述,稳定性是一个比较复杂的问题[2],在研究方法上应综合考虑森林生态系统的群落特征、空间结构、生物量以及外部环境等因素,构建全面且客观的稳定性评价指标体系,并选用合适的方法,以对森林群落稳定性作出定量评价[13]。辽宁仙人洞国家级自然保护区是以森林生态系统为保护对象的综合性自然保护区,地处华北植物区系与长白植物区系的交错地带,植物地理成分复杂。区内保存有目前亚洲面积最大的原生型赤松Pinus densiflora-栎Quercus林顶级群落,同时还有银杏Ginkgo biloba、东北红豆杉 Taxus cuspidata、杜仲 Eucomnia ulmoides、野大豆 Glycine soja 和三桠乌药 Lindera obtusiloba等国家级保护植物以及东北罕见的亚热带植物[14]。该地区森林生态系统在涵养水源、保持水土,调节小峪河、英那河水量水质,促进生态旅游开发和周边第三产业发展等方面具有重大作用。以往的研究多关注保护区内植物区系[15]、林下土壤[16]等,对其森林生态系统稳定性尚未有相关研究。因此,本研究以辽宁仙人洞国家级自然保护区内原生的典型森林群落为研究对象,集合种群更新潜力、群落基本特征、物种多样性、种群生态位重叠和凋落物特性等群落自身特征,以及森林土壤肥力等指标,运用模糊综合评判的隶属函数值法对各森林群落的稳定性进行综合评价,以期为该保护区森林生态系统的有效保护及管理提供科学依据。
1 研究区概况
辽宁仙人洞国家级自然保护区位于辽宁省大连市庄河境内的北部山区 (39°54′00″~40°03′00″N,122°53′24″~123°03′30″E),总面积 3 574.7 hm2。地貌为侵蚀、剥蚀低山和丘陵。地带性土壤为石灰岩棕壤性土棕壤(60%)和石灰岩典型棕壤(30%)。年平均气温8.9 ℃,年平均降水量799.0 mm,属于暖温带湿润季风气候,四季温和,雨热同季,并具有一定的海洋性气候特点[14]。研究区内植物种类丰富,森林植被主要以赤松为主,此外还广泛分布有蒙古栎Quercus mongolica、栓皮栎Q. variabilis、麻栎Q.acutissima、槲树Q. dentata等;灌木层主要有青花椒Zanthoxylum schinifolium、榛Corylus heterophylla、牛 叠 肚Rubus crataegifolius、 白 檀Symplocos paniculata、 照 山 白Rhododendron micranthum、 胡 枝子Lespedeza bicolor等;草本层主要有矮丛薹草Carex callitrichosvar.nana、内折香茶菜Isodon inflexus、变色白前Cynanchum versicolor、辽宁堇菜Viola rossii、京芒草Achnatherum pekinense、菴闾Artemisia keiskeana等。
2 研究方法
2.1 样地设置及调查
辽宁仙人洞国家级自然保护区的典型地带性植被为赤松林、赤松栎林和多种栎组成的落叶阔叶林,集中分布于核心区内,受林窗干扰较小[14]。于2018年6—7月,在辽宁仙人洞国家级自然保护区核心区内选取9个典型森林群落(赤松林、赤松栎林和落叶阔叶林3种森林植被类型的代表性群落各3个)为研究对象。在每个森林群落内设置20 m×30 m的调查样地1块(表1),其经纬度、海拔高度、坡度、坡向等信息利用易测宝地理信息系统(GIS)和手持罗盘测定。对样地内所有胸径≥3 cm的树木进行每木检尺,记录其树种名称、树高、胸径、冠幅、树龄等;在每块样地的中心和四角共设置5 m×5 m的灌木样方5个,调查记录其种类组成、基径、丛高、冠幅和总覆盖度等;在每个灌木样方中心各设置1个1 m×1 m的草本样方,调查记录草本层植物的总覆盖度、种类组成及各植物种的盖度、株丛数和平均高度。在每块20 m×30 m样地内沿对角线等分选取3个土样、凋落物采集点,采集0~20 cm层扰动土样和表层原状土样(容积100 cm3的环刀),其中扰动土样用于测定土壤比重、有机质质量分数和主要养分质量分数,原状土样用于测定土壤容重;同时采用1 m×1 m样方收获法区分未分解层和半分解层,采集凋落物样品,记录其鲜质量和风干质量,以风干质量测算其蓄积量,并采用浸泡实验法测定了其最大持水率和最大持水量。

表1 森林群落调查样地基本情况Table 1 Basic situation of the sample plots in the forest communities
2.2 评价指标体系
2.2.1 种群更新潜力 选取乔木树种的幼苗、幼树占乔木总植株数的比例作为评价因子,规定胸径DBH≤1 cm 的个体为幼苗,1<DBH<3 cm 为幼树,DBH≥3 cm 为成树[10]。
2.2.2 群落基本特征 选取郁闭度、平均胸径、平均树高、平均年龄和物种种数作为评价因子,其中林下物种种数包括林下灌木层和草本层的所有植物种类。


2.2.5 凋落物特性 选取未分解层和半分解层凋落物的蓄积量、最大持水量、最大持水率,以及凋落物总蓄积量和总最大持水量作为评价因子。以风干质量估算不同林分凋落物的蓄积量,采用室内浸泡法测定凋落物的最大持水量及其吸水速率。
2.2.6 土壤肥力 选取土壤容重、土壤比重和有机质、有效钾、碱解氮、有效磷质量分数作为评价因子。土壤容重和比重分别采用环刀法和比重计法测定,土壤有机质质量分数采用重铬酸钾氧化-外加热法测定,土壤有效钾、有效氮和有效磷质量分数采用X荧光光谱法测定。
2.3 研究方法
采用主成分分析法比较6项评价指标对研究区9个典型森林群落稳定性的影响程度[20]。基于以上6项指标应用模糊数学中隶属函数值法进行稳定性综合评价。参与评价因子原始数据的标准化处理方法参照文献[10]。隶属函数值的计算公式如下其中:(U)Xij为第i个群落第j项评价指标的隶属函数值,且(U)Xij∈[0,1];Xij为第i个群落第j项评价指标所有因子标准化数据的平均值;Xjmin和Xjmax分别为所有参与评价群落第j项评价指标所有因子标准化数据平均值的最小值和最大值[10]。通过计算每个森林群落各项指标隶属函数值的平均值,得出各森林群落稳定性的高低排名。采用单因素方差分析(one-way ANOVA)和最小显著差法(LSD)对不同森林群落中物种多样性、凋落物特性和土壤肥力指标进行显著性检验。显著性水平设定α=0.05。
3 结果与分析
3.1 种群更新潜力
由表2可知:9个森林群落的幼苗、幼树比例存在明显差异。赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落幼树比例最高、幼苗比例较高,树种丰富,以花曲柳Fraxinus rhynchophylla、栓皮栎和槲栎为主,种群更新潜力最好。麻栎+栓皮栎-青花椒+榛-矮丛薹草群落幼苗比例最高(55.97%),但幼树很少,种群更新潜力次之。槲树+赤松-榛-矮丛薹草群落和赤松-青花椒-矮丛薹草群落幼苗、幼树比例中等,但组成存在差异;槲树+赤松-榛-矮丛薹草群落的幼树以槲树为主,赤松-青花椒-矮丛薹草群落的幼树种类相对丰富,如赤松、花曲柳和核桃楸Juglans mandshurica等。两者种群更新潜力相近,其他群落仅有少量幼苗或幼树,且种类简单,种群更新潜力较低。
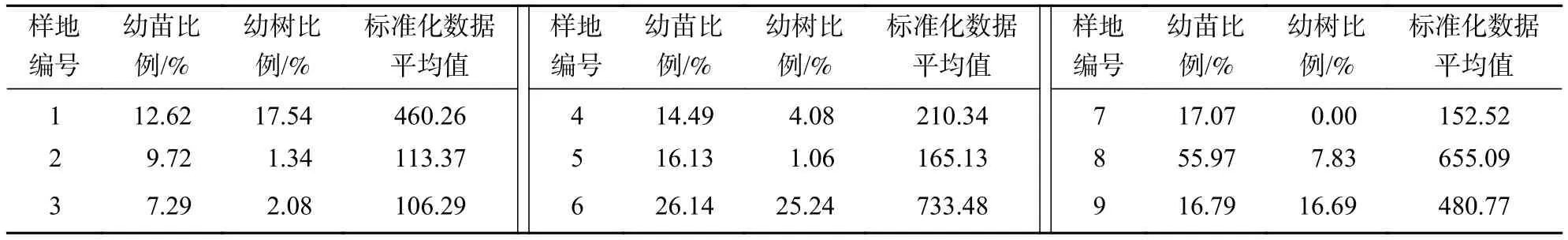
表2 森林群落种群更新潜力Table 2 Regenerative potential of the forest population
3.2 群落基本特征
由表3可知:蒙古栎+赤松-牛叠肚+照山白-矮丛薹草群落、蒙古栎-牛叠肚-矮丛薹草群落和赤松-牛叠肚+青花椒-矮丛薹草群落平均树高均大于10 m,说明群落内乔木的生存空间处于较上层,但由于蒙古栎+赤松-牛叠肚+照山白-矮丛薹草群落和蒙古栎-牛叠肚-矮丛薹草群落均以蒙古栎为优势林层且株数多,蒙古栎分枝多叶片大,树冠覆盖范围广,故两者林冠郁闭度均大于0.80,而赤松-牛叠肚+青花椒-矮丛薹草群落的优势树种赤松株数较少,故林冠郁闭度仅0.68;赤松-青花椒-矮丛薹草群落和赤松-三桠乌药+照山白-矮丛薹草群落郁闭度、平均树高等与其他群落差异不大,但草本层植物种类较丰富,如尾叶香茶菜Isodon excisus、甘菊Chrysanthemum lavandulifolium和长萼鸡眼草Kummerowia stipulacea等仅发现于赤松-青花椒-矮丛薹草林群落,毛缘宽叶薹草Carex siderostictavar.pilosa、中华蹄盖蕨Thyrium sinense和紫草Lithospermum erythrorhizon等仅发现于赤松-三桠乌药+照山白-矮丛薹草群落,说明林下植被的种类除受林冠层影响,还可能与土壤、坡度等环境因素有关。
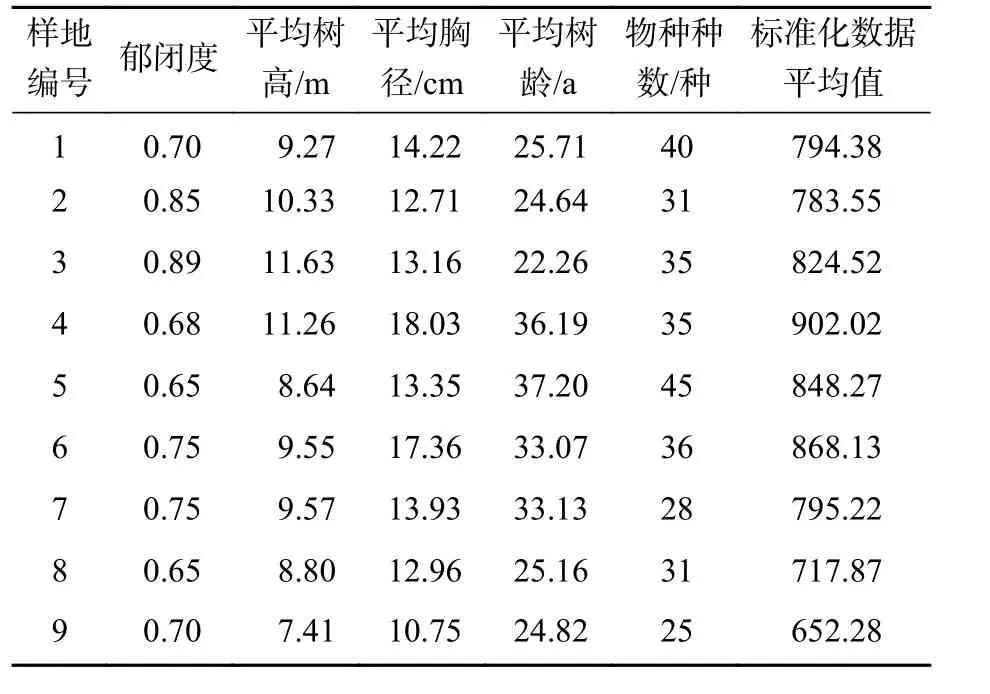
表3 森林群落基本特征Table 3 General characteristics of the forest communities
3.3 物种多样性
森林群落具有明显的垂直分层结构。由表4可以看出:研究区各森林群落不同功能层的物种多样性表现不一致。各群落乔木层物种多样性差异明显,槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落、赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落和槲树+赤松-榛-矮丛薹草群落的Shannon-Wiener指数和Simpson指数较大,最小的为蒙古栎-牛叠肚-矮丛薹草群落;Pielou指数和Alatalo指数的平均值最大的为槲树+赤松-榛-矮丛薹草群落,蒙古栎-牛叠肚-矮丛薹草群落和赤松-牛叠肚+青花椒-矮丛薹草群落的均匀度指数较小;赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落的Margalef指数最大,蒙古栎+赤松-牛叠肚+照山白-矮丛薹草群落和蒙古栎-牛叠肚-矮丛薹草群落乔木层仅有蒙古栎和赤松2个种群,Margalef指数较小。不同森林群落间灌木层Shannon-Wiener指数差异极显著(P<0.01),Margalef指数差异不显著(P>0.05);草本层仅Pielou指数差异显著(P<0.05)。将森林群落各层次综合来看,9个森林群落物种多样性指标差异从大到小依次表现为赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落、赤松-三桠乌药+照山白-矮丛薹草群落、槲树+赤松-榛-矮丛薹草群落、槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落、麻栎+栓皮栎-青花椒+榛-矮丛薹草群落、赤松-青花椒-矮丛薹草林群落、蒙古栎+赤松-牛叠肚+照山白-矮丛薹草群落、赤松-牛叠肚+青花椒-矮丛薹草群落、蒙古栎-牛叠肚-矮丛薹草群落。这与各功能层物种多样性变化在一定程度上具有一致性。

表4 森林群落物种多样性指数Table 4 Species diversity indexes of the forest communities
3.4 种群生态位重叠
由表5可知:多数群落灌木层种群生态位重叠值为0的种对占全部种对的比例低于草本层,说明灌木层种对重叠情况多于草本层。草本层存在一些生态位完全重叠的种对,说明草本层部分植物对生态因子的需求相似性较高。各群落种群生态位重叠值主要集中在0~0.6,说明群落内种间生态位分化程度低,种群生态位重叠现象普遍存在但程度较弱,不同物种共同利用或占有某一资源的竞争较缓和。从总体看,9个森林群落种群生态位重叠分布格局相似,且重叠程度不明显。

表5 森林群落种群生态位重叠指数分布格局Table 5 Distribution pattern of niche overlap index of the forest communities
3.5 凋落物特性
由表6可知:9个森林群落的凋落物蓄积总量为2.87~16.33 t·hm-2,差异极显著(P<0.01)。蒙古栎-牛叠肚-矮丛薹草群落的半分解层凋落物蓄积量最高,槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落未分解层凋落物蓄积量和总蓄积量最高,赤松-三桠乌药+照山白-矮丛薹草群落和赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落的凋落物总蓄积量也较高。凋落物蓄积量是影响凋落物层持水性能的特征之一[21],最大持水量决定最大持水率。9个森林群落凋落物总最大持水量为70.90~589.04 t·hm-2,差异极显著(P<0.01),槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落凋落物各层最大持水量均为最高,赤松-青花椒-矮丛薹草群落各层最大持水率和最大持水量均较低,而各群落内部未分解层和半分解层的最大持水量和持水率相差不大。综合比较,槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落凋落物层表现最好,在拦蓄降水、改良土壤肥力方面作用显著。

表6 森林群落凋落物特性Table 6 Litter characteristics of the forest communities
3.6 土壤肥力
由表7可知:9个森林群落的土壤比重差异不显著(P>0.05),容重差异极显著(P<0.01),反映出各群落土壤固相组成物质的种类及含量相似但紧实程度不同。槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落容重较大,孔隙较少,其储水性能较好,透水性差。不同森林群落土壤有机质、有效钾、碱解氮和有效磷质量分数差异极显著(P<0.01),赤松-三桠乌药+照山白-矮丛薹草群落和赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落的有机质质量分数较高且显著高于其他群落;槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落的有效钾质量分数最高,达2.65%;赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落和赤松-三桠乌药+照山白-矮丛薹草群落的碱解氮和有效磷质量分数较高,但与其他群落相差不大。综合比较,赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落的土壤肥力指标排名最高。

表7 森林群落土壤肥力Table 7 Soil fertilities of the forest communities
3.7 群落稳定性评价结果
综合比较各森林群落6项指标隶属函数值平均值的大小可以评判森林群落稳定性状况。从评价结果可知(表8):9个森林群落稳定性从大到小依次为赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落、槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落、麻栎+栓皮栎-青花椒+榛-矮丛薹草群落、赤松-三桠乌药+照山白-矮丛薹草群落、赤松-青花椒-矮丛薹草群落、蒙古栎+赤松-牛叠肚+照山白-矮丛薹草群落、蒙古栎-牛叠肚-矮丛薹草群落、赤松-牛叠肚+青花椒-矮丛薹草群落、槲树+赤松-榛-矮丛薹草群落。

表8 森林群落各指标隶属函数值及平均值Table 8 Subordinate function values of community stability
3.8 森林群落稳定性影响因素主成分分析
对种群更新潜力等6项森林群落稳定性评价指标的主成分分析结果显示(表9):前3个主成分的累积贡献率达到81.17%,其中第1主成分特征值为1.90,贡献率为31.66%。由表10可见:第1主成分中种群更新潜力、物种多样性和土壤肥力的荷载系数最大,分别为0.79、0.82和0.75,表明这3项指标对森林群落稳定性的影响程度较大。
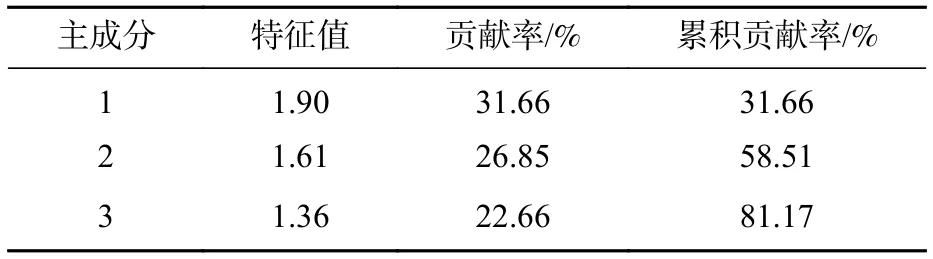
表9 主成分分析结果Table 9 Results of principal components analysis

表10 森林群落稳定性影响因素载荷矩阵Table 10 Component matrix for forest community stability factors
4 讨论与结论
4.1 讨论
4.1.1 种群更新潜力对群落稳定性的影响 森林群落的稳定性由群落内外多种因素共同决定,群落内的种群差异可以看作稳定性差异的表现。乔木层作为森林群落的优势层,对群落具有支配作用[22]。不仅林冠郁闭度会影响林下植被的种类与数量,同时乔木幼苗和幼树是林分天然更新的基础,因此群落的种群更新潜力高,森林群落持续存在且健康发展的能力好,在一定程度上提高了群落的稳定性。本研究中,赤松和麻栎群落的种群更新潜力相对最高,更新层中栎属Quercus乔木如栓皮栎和槲栎的幼苗、幼树较多,其根系较发达且耐荫性强,能够较好地适应林下的荫蔽环境,逐渐生长并进入优势林层,因此就种群更新潜力而言,该群落稳定性最高。这与张梦弢等[23]的研究结果一致,即云杉Picea jezoensis-冷杉Abies nephrolepis针阔混交林更新潜力明显好于其他2种森林群落,稳定性也最好。
4.1.2 物种多样性对群落稳定性的影响 物种多样性是描述群落结构和功能的数量性指标,多样性和稳定性的关系也一直备受关注。研究者们通过生物多样性实验、数学模型和理论综述等方法对其展开研究,但结论并不完全一致[24]。大量实证研究表明:物种多样性通过功能多样性和功能冗余的增加实现对群落稳定性的正效应[25]。物种多样性高的森林群落,不仅包含的种群数量多,而且各种群对生态因子的需求存在差异,因此面临干扰时,群落抵抗波动的能力强,受到扰动后的恢复力也强[26],有利于群落的稳定。但部分学者结合数学模型得出结论,多样性和稳定性呈负相关关系或者两者之间并不存在明显的联系[27-28]。本研究中,各群落的稳定性高低与物种多样性大小不一致,如赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落、槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落和麻栎+栓皮栎-青花椒+榛-矮丛薹草群落的物种多样性递减,其稳定性也逐渐降低;而蒙古栎-牛叠肚-矮丛薹草群落物种多样性最低,稳定性却高于赤松-牛叠肚+青花椒-矮丛薹草群落和槲树+赤松-榛-矮丛薹草群落。该研究结果与于晓文等[2]对冀北辽河源河岸林群落的研究结果相似。这表明物种多样性是群落稳定的必要条件,但物种多样性与群落稳定性并非一定是正相关关系。对这一情况的解释主要有以下2种:①群落各功能层对稳定性的影响程度不同,导致不同层次物种多样性与群落总体稳定性的关系表现并不一致。例如张明霞等[9]对秦岭松栎混交林的研究结果显示:松栎混交林的稳定性与乔木层的物种多样性呈负相关,与林下植被层的物种多样性呈正相关;②可能存在着1个多样性阈值,在该值以下,物种多样性表现为调节控制和种间协调功能,因此物种多样性与稳定性呈正相关关系;高于该值后,物种多样性变化对群落结构和功能的影响不大,群落稳定性差异不明显[2]。另外,由于稳定性存在多种定义,同一系统可能表现出因不同稳定性定义而产生不同的多样性—稳定性关系。
4.1.3 土壤肥力对群落稳定性的影响 土壤物理因素和化学因素等共同决定了土壤肥力大小。本研究中,槲树+赤松-榛-矮丛薹草群落土壤容重较大,土壤有机质质量分数仅2.85%,同时群落土壤碱解氮、有效磷、有效钾质量分数低,土壤肥力较差,制约着森林植被长期持续发展,导致群落稳定性较低。土壤作为森林群落必不可少的组成部分,为植被的存在和生长提供水、肥、气、热等必需条件。土壤容重与土壤质地、结构和有机质质量分数等有关,影响着土壤中水分、空气和养分的运输;土壤有机质作为土壤肥力的重要指标之一,是植物营养元素的给源,也是制约土壤理化性质的关键因素;土壤中氮、磷、钾是植物生长最主要的三大营养物质[29],它们的含量与有机质含量显著正相关。凋落物层作为森林群落中联结土壤与植物的纽带,是森林土壤有机碳的主要来源,影响土壤微生物群落活性和功能发挥[30],不同分解和积累程度凋落物下,土壤的养分组成和腐殖质性质都具有明显的不同。因此,积累较厚且分解程度高的凋落物层能有效减弱土壤淋溶作用,利于氮磷富集和有机质积累,土壤肥力得到改善[31],有利于植被生长和群落的稳定;反之,若林地内凋落物层稀薄,则导致土壤肥力较低,不利于森林群落的持续发展。
4.2 结论
本研究综合考虑森林群落自身特征和土壤肥力,应用模糊综合评判的隶属函数值法,对辽宁仙人洞国家级自然保护区9个森林群落的稳定性进行了综合评价,取得以下结论:①辽宁仙人洞国家级自然保护区的9个典型森林群落稳定性差异显著,由高到低依次为赤松+麻栎-地锦+青花椒-两型豆+白花碎米荠群落、槲树+麻栎-花木蓝+榛-菴闾+矮丛薹草群落、麻栎+栓皮栎-青花椒+榛-矮丛薹草群落、赤松-三桠乌药+照山白-矮丛薹草群落、赤松-青花椒-矮丛薹草群落、蒙古栎+赤松-牛叠肚+照山白-矮丛薹草群落、蒙古栎-牛叠肚-矮丛薹草群落、赤松-牛叠肚+青花椒-矮丛薹草群落、槲树+赤松-榛-矮丛薹草群落。②种群更新潜力影响群落的恢复力,物种多样性提高了群落的抵抗力,土壤肥力为群落发展提供良好基础。三者是影响森林群落稳定性的主要因素。
猜你喜欢
广东蚕业(2022年4期)2022-11-23 13:31:55
基层中医药(2021年8期)2021-11-02 06:25:06
今日农业(2020年15期)2020-12-15 10:16:11
湖北农机化(2020年16期)2020-10-21 01:29:08
江西农业(2020年13期)2020-09-08 07:44:54
作文通讯·高中版(2017年11期)2017-12-20 08:09:43
海峡姐妹(2017年9期)2017-11-06 08:39:39
汉语世界(2017年5期)2017-09-21 07:44:37
花木盆景·盆景赏石(2010年7期)2010-07-28 05:39:30
花木盆景·盆景赏石(2010年6期)2010-06-19 05:53:06
