
麦类全谷物中生物标记物质的研究进展
2022-06-23 07:26:18钱海峰齐希光
中国粮油学报 2022年5期
王 犇, 李 言, 钱海峰, 张 晖, 齐希光, 王 立
(江南大学食品学院,无锡 214122)
谷物是人类食物的重要来源之一,谷物籽粒包含了我们人体所需的营养成分(蛋白质、脂肪和碳水化合物),同时在谷物皮层、胚芽中也富含膳食纤维、维生素、矿物质和多种植物化学物等功能性成分(酚酸、黄酮类、植酸、木脂素和植物甾醇等)[1, 2]。目前,大多数谷物都是经过碾磨精制后食用,造成谷物外层营养物质严重流失,因此,保留有胚芽和麸皮的全谷物越来越受人们重视[3, 4]。研究表明,食用全谷物食品有降低心血管疾病[5]、调节Ⅱ型糖尿病[6, 7]以及降低一些癌症风险的作用[8, 9]。美国谷物化学家协会(AACC)在1999年对全谷物的定义为:全谷物应由完整、碾碎、破碎或压片的颖果组成,其主要组成部分包括淀粉胚乳、胚芽和麸皮,各部分的相对比例与完整颖果相同[10]。近年来,全谷物食品发展迅速,据统计[11],截至2021年1月,全球63个国家总共有超过13 000种带有全谷物标签的产品,相比2011年的3 378种,增长超过350%。然而,由于全球缺乏统一的全谷物标准,尤其是起步较晚的发展中国家,造成全谷物食品生产商各自为营,导致产品质量良莠不齐[12, 13]。此外,目前也缺乏单独能准确衡量不同谷物摄入量的方法,导致难以评估全谷物对人体健康的作用[14]。传统的饮食摄入量的评估通常采取饮食记录、饮食问卷和采访的方式进行,这种自我评价方法具有一定的主观性,很可能削弱了全谷物与疾病之间的关联[15, 16]。因此,寻找出能反映全谷物或其特定部位精准摄入量的成分显得至关重要。
谷物中的生物标记物是谷物麸皮或胚芽中特有的物质,能较好反映麸皮或胚芽的含量进而用来监测脱壳、脱麸等生产过程以及鉴定全谷物产品是否有掺假;可以反映谷物的摄入量,有助于研究特定谷物食品对健康影响的潜在机制[14, 17]。欧美等国已经进行了许多全谷物生物标记物质在食品标准和人体代谢方面的研究,我国全谷物产品行业也推荐以烷基间苯二酚作为全麦粉品质的鉴定指标[13, 17, 18]。作为全谷物的生物标记物一般应满足以下条件[19]:只存在于某种或某几种特定谷物的麸皮或胚芽中(精制谷物中不存在或含量极低);基本不受加工的影响,加工后的全谷物及其产品中能检测到该生物标记物;人体摄入后,能在血浆、尿液或组织中检测到该物质或其衍生物,且摄入量有较好的正相关性。研究发现,谷物中具有作为生物标记物潜力的物质有烷基间苯二酚[20-22]、苯并恶唑嗪酮[23, 24]、蒽酰胺[25, 26]、甾体皂苷[27, 28]、大麦芽胍硷[29, 30]和酚酸[22, 31, 32]等(表1)。本文主要介绍了小麦、大麦、黑麦和燕麦等麦类全谷物中具有代表性的潜在生物标记物的结构、存在部位、加工方式对其影响以及人体摄入后的代谢与检测,以期为麦类全谷物生物标记物的评估以及麦类全谷物食品品质提供保障。

表1 全谷物中潜在的生物标记物
1 烷基间苯二酚
1.1 结构及存在部位
烷基间苯二酚(Alkylresorcinols,ARs)是一种1,3二羟基苯(间苯二酚)衍生物,具有两亲性,在苯环的五号位上有长度为14~27个碳原子,0~4个双键的烷基链,它们大多为饱和的奇数烷基链[41](图1)。ARs几乎只存在于黑麦、小麦和大麦类谷物的麸皮中[34],质量分数分别为360~3 200、317~1 010、44~500 μg/g[33]。黑麦中有85%的烷基链为饱和烷基链,主要是AR 16∶0~AR 25∶0,有15%的烷基链为AR n∶1~AR n∶3的不饱和烃链(烯基间苯二酚)[41, 42]。小麦中的AR 19:0和AR 21∶0的质量分数最丰富,其次是AR 25∶0、AR 23∶0和AR 17∶0[43]。大麦中AR 25∶0的质量分数最多(占总AR的35.6%),其次是AR 21∶0和AR 23∶0(分别占总AR的29.2%和21%)[44]。

注:n1为17、19、21、23、25,n2为0、1、2、3。图1 主要的烷基间苯二酚及其衍生物的分子结构
1.2 加工对其影响
1.2.1 碾磨处理
由于ARs几乎都存在于麸皮中,因此精制小麦粉的总ARs含量基本为零[34]。回添式全麦粉的制作通常是先将麦粒碾磨精制成面粉后再将其与处理后的麸皮和胚芽按比例混合,Carcea等[45]采用石磨和辊磨的方式碾磨软质小麦,再将碾磨后的麸皮按谷粒原始比例混合,发现ARs含量与原始谷粒含量相同,说明碾磨对ARs的影响非常小。Ciccoritti等[46]建立了一种基于部分去糠的微粉化和气流分级小麦粉加工工艺,用于提高面食中的麸皮含量,用此方法加工小麦粉制备的意大利面中ARs质量分数增加了121%。
1.2.2 其他加工处理
谷物产品在食用过程中都需要经过热加工过程,因此研究不同热加工方式对ARs的影响非常重要。Tian等[47]比较了烘烤、微波、挤压和高压热处理对小麦麸皮中ARs的影响,结果表明ARs损失量为烘烤处理>高压热处理、挤压处理>微波处理,其中,烘烤处理温度在110 ℃内ARs含量下降缓缓慢。此外,在刚开始的高压热处理时,由于麸皮含水量高而传热快,加速了酶对ARs的降解,而随后温度升高钝化了酶,从而使ARs趋于稳定。同样,Michniewicz等[48]发现在120 ℃下高压灭菌条件下对黑麦处理30 min,ARs含量变化较小,说明控制加热条件可以减少ARs的损失。Ciccoritti等[49]对2种硬粒小麦进行水热、热风和冷冻干燥3种预处理,发现热风干燥和水热干燥处理均提高了ARs的提取量,两种硬粒小麦在130 ℃处理条件下提取量分别提高了19%和14%。Ciccoritti等[50]的另一项研究中,将硬粒小麦在水中浸泡4 h后再进行水热处理(121 ℃,10 min),使ARs的可提取含量增加了48%,这可能是由于细胞内受热形成水蒸气导致细胞破裂,封闭ARs的细胞膜被破坏,使得ARs的含量增加[49]。因此,合理控制热处理条件可以减少ARs的损失而使其保持稳定,适当的水热处理甚至可以提高AR的提取能力。此外,Katinna等[51]研究表明天然黑麦麸皮在发酵过程中其ARs含量略有增加,因为在天然麸皮发酵过程中谷物内源性酶和微生物产生的酶可以降解释放结合在细胞壁上的酚类化合物。
1.2.3 加工成品食物
不同谷物食品中ARs含量也有不同,Andersson等[52]测定了挪威25种面包和饼干中的ARs含量,结果表明不同食品中ARs的质量分数有很大差异(21~548 μg/g),但这些食品中ARs的计算量与检测量高度相关。Gunenc等[53]比较了相同烘焙条件下不同配方面包中的ARs,发现所有测试的面包均能检测到ARs,其中,含小麦麸皮和ARs提取物面包中的ARs含量和同系物组成没有显著变化。Andersson等[52]将面包中ARs含量换算为全谷物含量后,发现所测得的结果与工厂提供的信息较为接近,表明了ARs在整个食品加工过程中保持稳定。Menzel等[54]测定了瑞典159种产品中的ARs含量和同系物组成,发现可以通过ARs含量和AR 17∶0/AR 21∶0比值来预测产品中全谷物小麦和黑麦的含量。
因此,ARs并不会因为加工而大量损失,而是有很好的稳定性且在成品食物中保持一定的含量,食品中的ARs含量与特定的全谷物或麸皮的消耗量相关。
1.3 摄入人体后的代谢及检测
人体摄入全谷物食品后可以在血浆、脂肪组织和红细胞膜中检测到ARs,在血浆和尿液中检测到ARs的代谢物[14, 35]。Landberg等[21]在一项干预研究中,30名志愿者在6周内食用全谷物食品,发现ARs摄入量与血浆中的ARs含量有明显正相关关系(r=0.59)。不同全谷物摄入人体后,ARs的同系物含量也有所不同,如全谷物黑麦摄入后血浆中AR 17∶0/AR 21∶0的比例高于摄入小麦的比例(黑麦中约为1.0,而小麦约为0.1),因此这一比例可用于区分摄入的全谷物种类[55]。Kyryo等[56]分析了10个欧洲国家共2 845名受试者血浆中的ARs,发现主要食用全小麦国家的受试者中,其AR 17∶0/AR 21∶0的比例显示较低,而主要食用全黑麦国家的受试者中,其比例则较高。Aubertin-Leheudre等[57]还发现血浆中的ARs与尿液中的ARs代谢物3,5-二羟基苯甲酸(DHBA)和3-(3,5-二羟基苯基)-1-丙酸(DHPPA)也显著相关(r分别为0.372和0.408),说明尿液中ARs代谢物来源于血浆中ARs。此外,Wu等[58]进行12周的全谷物饮食干预研究发现,全谷物摄入量与脂肪组织中的ARs浓度高度相关(r=0.60~0.84),表明脂肪组织中的ARs有作为全谷物摄入量生物标记物的潜力。
ARs在体内的代谢依赖于细胞色素P450(CYP450),后者使ARs的烷基链末端插入羟基,随后被氧化成羧基,然后该衍生物发生β氧化而被逐步降解,进而形成2种主要的代谢产物:DHBA和DHPPA[59]。ARs在代谢过程中能够与葡萄糖醛酸苷或硫酸基团偶联,之后会解偶联并进入尿液[20]。有研究报道在非谷物类食品中也有少量存在DHBA和DHPPA,并在尿液中检测到了其进一步的衍生物:2-(3,5-二羟基苯甲酰胺)乙酸(DHBA-glycine)和5-(3,5-二羟基苯基)戊酸(DHPPTA),且后者目前只被认为是ARs的独特代谢物[59]。血浆ARs表观半衰期相当短(4~6 h)[19, 61],而其4种代谢物(DHBA、DHPPA、DHBA-glycine和DHPPTA)则较长(10~16 h)[19, 60],因此具有反映全谷物小麦/黑麦摄入量的潜力。
ARs具有很高的生物标记潜力,能较好反映全谷物小麦/黑麦的摄入量,且能根据同系物的比例来区分小麦和黑麦。对ARs代谢物的综合分析比单独分析其中一种能更好地反映全谷物摄入量,但对于饮食不稳定、个体差异大的人群仍不能精确反映其全谷物摄入量[14, 59]。
2 苯并恶唑嗪酮
2.1 结构及存在部位
苯并恶唑嗪酮(Benzoxazinoids,Bx)是一种吲哚衍生类化合物,根据其化学结构可分为三类:苯并恶唑啉酮类、内酰胺类和羟肟酸类,常见的苯并恶嗪类化合物包括有苯并恶唑啉酮类的1,3-苯并恶唑-2-酮(BOA)和6-甲氧基-1,3-苯并恶唑-2-酮(MBOA),内酰胺类的2-羟基-1,4-苯并恶嗪-3-酮(HBOA)和2-羟基-7-甲氧基-1,4-苯并恶嗪-3-酮(HMBOA),羟肟酸类的2,4-二羟基1,4-苯并恶嗪-3-酮(DIBOA)和2,4-二羟基-7-甲氧基-1,4-苯并恶嗪-3-酮(DIMBOA)[62](图2)。Bx类化合物存在于黑麦、玉米和小麦的根、茎和叶片等部位,在小麦和黑麦麦粒中的平均含量分别为4.8、95 μg/g[62, 63]。Bx主要存在于小麦和黑麦的胚芽和麸皮中,胚芽中Bx含量高于麸皮,其中主要的Bx是DIBOA和HBOA的双己糖衍生物(DIBOA-Glc-Hex和HBOA-Glc-Hex),其次是DIBOA-Glc 和DIBOA[63]。
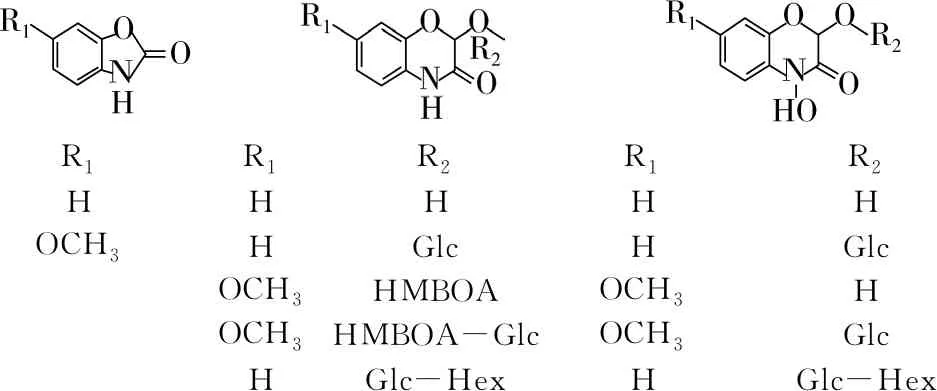
R1R1R2R1R2HHHHHOCH3HGlcHGlcOCH3HMBOAOCH3HOCH3HMBOA-GlcOCH3GlcHGlc-HexHGlc-Hex
2.2 加工对其影响
浸泡处理能促使双糖基化羟肟酸和内酰胺水解,水热处理能够促进酶促水解使Bx浓度增加。Tanwir等[63]发现浸泡6 h促使黑麦中DIBOA-Glc-Hex和HBOA-Glc-Hex的浓度下降, DIBOA和DIBOA-Glc浓度分别增加了约20倍和4倍;同时发现,煮沸过程中只有少量的DIBOA-Glc-Hex转化,DIBOA-Glc浓度增加了2倍,而HBOA-Glc-Hex的浓度没有下降。Pedersen等[64]对1种小麦和2种黑麦进行水热处理再分别磨成小麦粉和黑麦粉,结果显示3种样品中总Bx质量摩尔浓度分别从0(未检测到)、377.2、159.6 nmol/g分别上升到4 077.4、6 079.0、3 542.6 nmol/g;此外还发现,当用对应的小麦粉和黑麦粉制作成面包后,除苯并恶唑啉酮外的Bx含量均显著降低,这是因为羟肟酸和内酰胺类化合物可能在焙烤过程中降解为苯并恶唑啉酮类化合物。
面包的发酵过程也会影响Bx的含量,Savolainen等[65]研究了发酵对麸皮中Bx的影响,发现添加酶和酵母菌促使DIBOA、DIMBOA和HBOA的双糖苷缀合物含量升高,添加乳酸菌可促进了双糖苷缀合物的进一步转化。此外,Dihm等[66]检测了北欧20种商业面包和3种传统面包中Bx的含量,小麦和黑麦面包中Bx质量分数分别为14~403、152~3 340 μg/g,甚至2种芬兰黑麦面包中的Bx质量分数高达2.3~3.3 mg/g,说明发酵和烘焙过程会显著影响其含量。
总之,浸泡、水热处理和发酵处理均能提高全谷物(小麦和黑麦)Bx的含量,且主要是通过酶的作用促使谷物中缀合的及结合的Bx化合物转化和释放。此外,加工对Bx含量影响不大,但其中机理不甚明确。
2.3 摄入人体后的代谢及检测
研究表明,人体在摄入全谷物食品(小麦和黑麦)后,血浆、尿液和组织中的Bx类物质的含量会增加[24, 67]。Adhikari等[67]研究发现,20名健康的受试者在摄入1周含Bx的膳食后,HBOA-Glc占尿液中总Bx的80%,而膳食中HBOA-Glc仅占总Bx的1%,因此HBOA-Glc有望作为Bx摄入的主要代谢标记物。Jensen等[68]研究发现Bx代谢较为快速,摄入仅1 h后血浆中就可检测到Bx并在3 h达到峰值,大部分Bx在12 h内被清除;尿液中的Bx在早餐后5 h检测到,甚至在36 h后还能检测到,在尿液中有延缓现象表明Bx可能会被存储于组织中并达到一定的稳定性。一些Bx化合物的葡萄糖醛酸化和硫酸化、羟肟酸的还原以及一些糖苷类化合物(如内酰胺)被肠道直接吸收是Bx吸收和代谢的主要机制[67]。
血浆和尿液中的一些Bx衍生物也逐渐显示其生物标记的潜力,Hanhineva等[24]在12名摄入黑麦面包志愿者的血浆中检测到5种Bx和4种Bx衍生物,其中2-羟基-N-(2-羟基苯基)乙酰胺(HHPAA)和N-(2-羟基苯基)乙酰胺(HPAA)在所有受试者中都能检测到,推测DIBOA和BOA会在体内转化为HHPPA和HPPA。Zhu等[32]通过非靶向代谢组学的方法鉴定出了尿液中5种Bx衍生物,发现其随全谷物摄入有相应的增长,尤其是在摄入6~9 h时间段;动力学曲线表明,Bx衍生物在摄入后的6 ~12 h左右可在尿液中检出,并且达到峰值,说明其在尿液中比在血浆中存在时间更长,有望为中长期尿液生物标记物来反映全谷物摄入,但Bx及其衍生物与全谷物摄入量的相关性的详细研究报道还较少。
3 蒽酰胺和皂苷
3.1 结构及存在部位
燕麦蒽酰胺(avenanthramides,AVAs)是燕麦中独有的酚类生物碱,根据结构中的双键数量可将AVAs分为C型和A型。C型AVAs含有单个双键,是由羟基肉桂酸与邻氨基苯甲酸通过酰胺键连接而成,A型AVAs是由含有2个双键的羟基苯烯酸与邻氨基苯甲酸通过酰胺键连接而成[25, 30, 69]。根据AVAs上取代基的不同,目前已发现至少27种AVAs,其中主要的3种是AVA-A(2p)、AVA-B(2f)和AVA-C(2c),它们是由5-羟基邻氨基苯甲酸分别与对香豆酸、阿魏酸和咖啡酸通过酰胺键链接而成的蒽酰胺[25, 30](图2)。AVAs存在于燕麦的叶片、谷壳和籽粒中,燕麦籽粒中的质量分数为2~323.7 μg/g[25, 70],燕麦籽粒中的AVAs主要存在于麸皮中,含量会随籽粒的逐渐成熟而增加,同时受到品种、年份、地点、栽培条件的影响而有不同[71]。
燕麦皂苷(Avenacosides,AVE)是燕麦中独有的甾体皂苷类物质,是1种纽替型甾体皂苷,主要分为两大类:AVE-A和AVE-B,AVE-A是1个纽替皂苷元在C-3处连接了1个鼠李糖和2个葡萄糖,在C-26处连接了一个葡萄糖的甾体皂苷;AVE-B是1个纽替皂苷元在C-3处与1个鼠李糖和3个葡萄糖连接,在C-26处与1个葡萄糖连接的甾体皂苷(图4)[25, 27]。燕麦中的AVE-A和AVE-B质量分数分别为360、300 μg/g,AVE-A占谷物总皂苷含量的41.9%~60.6%, AVE-B占谷物总皂苷含量的35.8%~55.2%[72]。

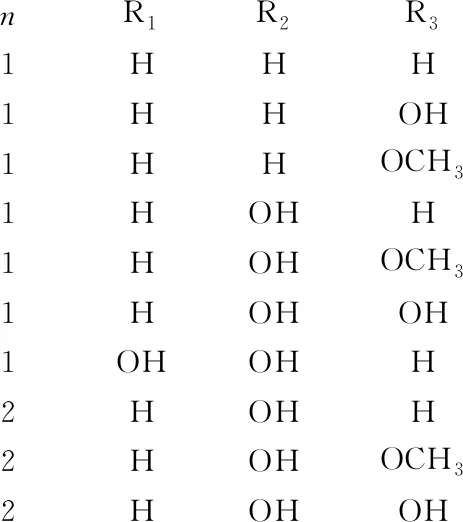
nR1R2R31HHH1HHOH1HHOCH31HOHH1HOHOCH31HOHOH1OHOHH2HOHH2HOHOCH32HOHOH

图4 2种主要燕麦皂苷分子结构
3.2 加工对其影响
3.2.1 加工对燕麦蒽酰胺的影响
Antonini等[73]将2个不同基因型燕麦进行工业脱壳,发现燕麦粒碾磨成全燕麦粉后,AVA 2p、AVA 2f和AVA 2c的比例不变,总质量分数有略微上升(23.9~42.5 μg/g),说明碾磨不会造成全谷物燕麦中的AVAs的损失。
对燕麦进行热加工会导致AVAs的浓度下降,有研究表明超高温(145 ℃,8 s)处理会导致AVAs质量分数下降42%~50%[74]。不同AVAs的热稳定性也不同,Bryngelsson等[78]比较了蒸煮和高压灭菌对燕麦中3种AVAs的影响,蒸煮导致AVA 2p质量分数下降了36%,而AVA 2c和AVA 2f含量没有显著变化;高压灭菌(100~120 ℃,16 min)使AVA 2c和AVA 2p均降低了37%而AVA 2f没有显著变化,说明AVA 2p对热最敏感,AVA 2f的热稳定性最高。
浸泡处理能够促进结合形式AVAs的水解和释放而使燕麦内源性AVAs的含量增加[76, 77]。Jasmeet等[77]将超高温处理后的燕麦粒与超纯水混合并在室温下(22 ℃)储藏12周,发现AVAs质量分数从起初的2.96 μg/g上升到13.88 μg/g。Bryngelsson等[78]也发现燕麦粒在80 ℃下浸泡15 min后AVAs上升了50%,在20 ℃下浸泡10 min后达到最大值(提高了100%)。因此,可以通过控制热加工强度和适当的预处理(如浸泡)能减少全谷物燕麦中AVAs的热损失。
Dimberg等[79]分析了5种含燕麦成分的食品(面包、茶饼、松饼、通心粉和新鲜意面)AVAs含量,发现除新鲜意面外其他4个燕麦食品的游离AVAs浓度均高于原料,推测原因可能是加工过程中 AVAs的重新合成和或结合态AVAs的释放。不同加工成品食物中的AVAs含量差异较大,Soycan等[80]测定了市售的22种燕麦产品中AVAs质量分数,质量分数最高的是燕麦麸皮和燕麦片(分别为49.6、48.8 μg/g),其次是燕麦饼(43.8 μg/g),燕麦卷的质量分数最低(28.3 μg/g)。
3.2.2 加工对燕麦皂苷的影响
关于燕麦皂苷在加工中的变化研究不多,Oenning等[81]研究了不同温度和pH条件下加热对AVE-A和AVE-B提取物的影响,结果发现AVE在100 ℃条件下能稳定3 h,在140 ℃下加热会降解,主要降解产物为去鼠李糖苷的AVE-A和AVE-B;同时还发现,在pH为4~6时,AVE-A的浓度随加热时间的延长而减少,且浓度与时间成线性相关(r=0.98~0.99)。Yang等[27]对市售的15种不同燕麦产品中AVE-A和AVE-B的质量分数进行了测定,AVE-A的质量分数为37.9~377.5 μg/g,AVE-B的质量分数为11.7~89.2 μg/g,不同产品中燕麦皂苷的质量分数差异较大,燕麦麸皮的AVE质量分数最高(300.4~443.0 μg/g),其次为燕麦片(100.9~293.9 μg/g),冷燕麦谷物产品中AVE的质量分数最低(49.6~90.7 μg/g)。因此,AVAs能够以一定的含量存在于一些燕麦产品中,但加工对燕麦皂苷的影响还处于初步阶段。
3.3 摄入人体后的代谢及检测
3.3.1 燕麦蒽酰胺的代谢及检测
AVAs被摄入人体后,能够在血浆和尿液等部位检测到[37, 82]。AVAs的吸收具有剂量依赖性,Zhang等[26]研究发现,摄入高质量分数AVAs(229.6 μg/g)燕麦饼干的受试者相比于低AVAs摄入量(32.7 μg/g)的受试者,其血浆中AVAs浓度要更高(质量浓度分别为4.26~8.39、1.33~1.98 ng/mL),达到峰值的时间更长(分别为2~3 h和1~2 h)。同样的,Chen等[82]报道,健康老人摄入1g富含AVA 2p、AVA 2f和AVA 2c的混合物(AEM)后,3种AVAs的浓度在2 h达到峰值。研究表明,AVAs的半衰期在5 h以内,不同AVAs的半衰期也不同, AVA 2p与AVA 2c半衰期相近(2~3 h),AVA 2f半衰期最长(2~5 h),可能是因为其具有更疏水的羟基肉桂酸部分,所以吸收和消除速度慢[26, 82]。
AVAs在代谢过程中会发生甲基化转化, AVA 2c在甲基化转移酶的作用下发生甲基化生成AVA 2f,两者均可还原生成二氢AVA-2c(DH-2c)和二氢AVA-2f(DH-2f),其中DH-AVA 2c是人体中AVA 2c的主要代谢物;这些物质还能通过酰胺键的断裂生成其他低分子代谢物,因此AVAs与其代谢物可能共同作为全谷物燕麦摄入的生物标记物[37, 83]。
此外,肠道微生物可能会影响AVAs的代谢和生物利用度,AVAs主要通过肠道菌群代谢而还原生成DH-AVAs,然而个体之间差异很大,一些受试者仅产生很少或不产生DH-AVAs,因此人体肠道菌群是十分重要的影响因素,还需进一步的研究[14, 25, 37]。
3.3.2 燕麦皂苷的代谢及检测
关于AVE的代谢研究非常少,Johansson等[84]研究发现,受试者在饮用燕麦麸皮饮料后,能在其血浆中检测到甾体皂苷骨架,这可能是AVE降解产生的纽替皂苷元,并推测纽替皂苷元可以人体被吸收并在肝脏中羟基化,然后进行葡萄糖醛酸化。在Wang等[28]的一项研究中,12名健康受试者在服用单剂量燕麦麸早餐后,尿液中AVE-A和AVE-B浓度迅速上升,并且分别在4.33、4.17 h达到峰值,半衰期分别为6.22、4.55 h;2种AVE在尿液中的含量分别为摄入量的2.26%和2.52%,这表明AVE有可能作为燕麦产品摄入的生物标记物。此外,研究还发现了AVE-A的3种代谢物和AVE-B的5种代谢物,AVE的去糖基化被认为是小鼠和人的主要代谢途径,肠道微生物在AVE的代谢中起主要作用[28]。目前还需更多的研究验证AVE作为全谷物燕麦生物标记物的潜力。
4 总结与展望
烷基间苯二酚、苯并恶唑嗪酮、燕麦蒽酰胺和燕麦皂苷等物质仅存在于少数麦类谷物的麸皮或胚芽中,经过加工后,在相应的全谷物食品中仍有一定的含量,可以作为鉴定全谷物品质的生物标记物质。同时,在摄入人体后,能够达到人体的血浆、尿液或脂肪等部位并且能够被检测到。但综合来看仍然存在以下问题:几种潜在的生物标记物质同系物种类多,结构复杂,且其含量受相应谷物的品种和生长环境等条件的影响而有很大的差别;不同加工过程对全谷物中生物标记物的影响的研究较少,还缺少其在更多全谷物食品中的信息;目前人体摄入全谷物中生物标记物的研究数量少,规模小,人体生物样本中的生物标记物含量与全谷物食品摄入量的相关性仍有待研究;能代表特定全谷物的生物标记物仍有限,除麦类全谷物外,因此还缺少能代表其他全谷物的新生物标记物。因此,在今后的研究中可以侧重以下方面:深入了解全谷物生物标记物的结构与其在谷物中分布情况,挖掘更多潜在的全谷物生物标记物;广泛研究多种加工方式对全谷物生物标记物的影响,确定更多全谷物产品中生物标记物信息以鉴定全谷物产品的品质;深入全谷物生物标记物在人群中的流行病学研究,适当扩大研究规模以更加精确评估全谷物生物标记物的标记能力和代谢机制,使未来深入了解全谷物摄入量与人体健康的关系成为可能。
猜你喜欢
中国农学通报(2022年21期)2022-08-22 01:12:30
河南畜牧兽医(2021年9期)2021-12-10 10:43:50
今日农业(2021年17期)2021-11-26 23:38:44
今日农业(2021年9期)2021-07-28 07:08:26
军事文摘(2020年20期)2020-11-16 00:32:10
现代家庭·生活版(2019年1期)2019-01-24 01:29:56
今日农业(2017年11期)2017-01-31 14:53:50
三联生活周刊(2015年21期)2015-05-21 16:59:46
食品工业科技(2014年15期)2014-03-11 18:17:49
食品工业科技(2014年15期)2014-03-11 18:17:20
