
寻找鸟类的姊妹群
2022-06-23 10:37李菲凡
化石 2022年2期
李菲凡
在20世纪60年代的科学运动——恐龙文艺复兴中,沉寂近4 0年的鸟类起源于兽脚类(Theropoda)恐龙的假说得到恢复,并获得普遍支持。兽脚类的哪一类群与鸟类亲缘关系最近,即作为鸟类的“姊妹群”成为了随之而来的一个问题。随着化石材料的丰富和系统发育分析方法的改进,对鸟类姊妹群的认识也不断进步,这个位置先后被赋予恐爪龙类(Deinonychosauria)、伤齿龙科(Troodontidae)、擅攀鸟龙科(Scansoriopterygidae)、近鸟龙科(Anchiornithidae)等类群,目前仍未结束相关的争议。

Paul(1984)提出的主龙类分支图,其中兽脚类主要类群的系统发育关系与现代基本相同。兽脚类部分的缩略语:T:兽脚类;m:巨齿龙;pr:原美颌龙;cl:腔骨龙;di:双脊龙;ce:角鼻龙;eu:美扭椎龙;y,永川龙;co:美颌龙科;or:嗜鸟龙;pr:原角鼻龙;al:异特龙;ty:霸王龙科;a:始祖鸟科;d:驰龙;sa:蜥鸟龙;b,鸟类;ov:窃蛋龙科;su:似鸵龙科(现为似鸟龙科的次异名)
早期的研究
自Hennig于20世纪50年代提出系统发育学理论以来,这一理论已成为指导生物分类和重建演化关系的重要依据。在恐龙方面,1984年,Paul首次提出了与现代观点基本一致的兽脚类系统发育树。然而,这一研究将驰龙科(Dromaeosauridae)的伶盗龙(Velociraptor)和伤齿龙科(Troodontidae)的蜥鸟龙(Saurornithoides)置于比始祖鸟科(Archaeopterygidae)更为衍化的位置,使得它们被归入了原始鸟类,而与鸟类最接近的恐龙则是霸王龙科(Tyrannosaurid)。
恐爪龙类作为鸟类的姊妹群
恐爪龙类(Deinonychosauria)通常包括驰龙科和伤齿龙科,是大众所较为熟知的类群,将该类群作为鸟类的姊妹群也是1990-2000年代影响力最大的一种分类法。一个重要的原因是:对恐爪龙(Deinonychus)自身的研究,尤其是将其与始祖鸟对比的研究,直接引发了恐龙文艺复兴和兽脚类起源说的复兴。

Gauthier(1986)提出的兽脚类分支图

顾 氏 小 盗 龙(Microraptor g u i)模式标本(引自Xu et al.,2003)
1986年,Gauthier利用系统发育分析探究了鸟类的起源,提出了重要的定义手盗龙类(Ma n i r apt or a),即所有接近鸟类而远离似鸟龙科(O r n it hom i m id ae)的虚骨龙类(Coelurosauria)。该研究发现,鸟翼类(Avialae,在该研究中与目前最常用的鸟类定义范围相同)和恐爪龙类互为姊妹群,而手盗龙类的其它类群间的亲缘关系不明。1990年代的多项系统发育研究在梳理其它手盗龙类关系的同时,进一步支持了恐爪龙类作为鸟类姊妹群的观点。
从20世纪末起,我国辽西地区的热河生物群出产了一系列保存精美的驰龙科小盗龙类(Microraptoria)的化石,它们拥有与鸟类几乎相同的羽毛,有些呈“四翼”形态,可能具有飞行或滑翔能力。这使得驰龙科相对于伤齿龙科,更加成为鸟类起源研究的焦点。此外,南半球的驰龙科半鸟亚科(Unenlagiinae)也经常被用于与鸟类进行比较,但它们有时被认为比其它驰龙科更接近鸟类,甚至自身就是鸟类的姊妹群。
近年来,随着更原始的驰龙科、有羽毛的伤齿龙科及基干鸟翼类的不断发现,以及系统发育数据集的扩大,以驰龙科作为研究重心的思路受到挑战。首先,小盗龙类若干类似鸟类的特征,包括不对称飞羽、飞行或滑翔的能力,可能是独立演化的树栖适应性特征,和鸟类的飞行起源并不是同一事件。新发现的热河生物群的小盗龙类——天宇盗龙(Tianyuraptor)和振元龙(Zhenyuanlong)反而大型化,前肢短,后肢适合奔跑,体现了和树栖类群的生态位分化。另一类更原始的驰龙科——哈兹卡盗龙亚科(Halszkaraptorinae)也具有截然不同的适应性特征,形态类似游禽,适应水栖。其次,发现了若干羽毛保存良好的伤齿龙科或原始鸟翼类标本,其正羽散乱、呈丝状或无明显分层,羽枝缺乏羽小枝,羽片不完全封闭,可能无法飞行或滑翔,也说明鸟类的现代羽毛形态和飞行能力可能另有起源。原本被认为前肢可像现代鸟类那样拍打的半鸟,被认为是肩胛骨方向的鉴定有误:实际与其它兽脚类一样,肩胛骨贴近胸廓侧面而非背侧,而肩臼朝向肩带的后腹侧。这也和基干鸟类的前肢仍不能像现代鸟类那样拍打的研究相符。而将半鸟亚科独立于驰龙科的观点,被认为过于强调各特征的独立性而忽视其总体性,且对许多特征的存在状态判断有误,因而几乎没有获得支持。
2005年,第一种保存了羽毛的伤齿龙科金凤鸟(Jinfengopteryx)被报道,其最早被归入始祖鸟科,这提示了伤齿龙科与鸟类的关联。2010年以来,恐爪龙类的有效性受到了诸多质疑,伤齿龙科在许多大型的系统发育分析中比驰龙科更加接近鸟类,这使得恐爪龙类成为驰龙科的次异名。驰龙科和伤齿龙科究竟能否维持单系性,还有待于未来的进一步研究。

擅攀鸟龙模式标本的线图及化石照片(引自Feduccia,2013)
擅攀鸟龙科作为鸟类的姊妹群
作为手盗龙类的一支,擅攀鸟龙科(Scansoriopterygidae)是2002年建立的,其每个属都具有显著区别于其它手盗龙类的特征,我们对其至今知之甚少。早期仅有的2件化石标本被命名为擅攀鸟龙(Scansoriopteryx)和树息龙(Epidendrosaurus),皆为幼体,是最小的非鸟类恐龙标本之一。现在通常将树息龙视为擅攀鸟龙的次异名。擅攀鸟龙的第三指格外延长,长于第二指,被认为是树栖特征。全身仅覆盖着结构简单的绒羽,缺乏包括飞羽在内的正羽,这对其系统发育位置而言很不寻常,因而被认为可能是幼体特征。2008年,张福成等报道了一属新的擅攀鸟龙科:耀龙(Epidexipteryx)。它与具有较长尾骨的擅攀鸟龙不同,具有短尾骨和类似尾综骨的愈合末端。耀龙最引人注目的特征是它的全长只有44.5cm,其中却有近20 c m是4条带状的尾羽,而其余体表也只有绒羽。此外,耀龙具有一些类似窃蛋龙类(Oviraptorosauria)的特征。

张福成等(2008)对耀龙进行的系统发育分析。擅攀鸟龙科属于基干鸟翼类,是鸟类的姊妹群
2000年代的系统发育分析常将擅攀鸟龙科置于比恐爪龙类更衍化的位置,作为鸟类的姊妹群。此时,鸟翼类的范围与鸟类不再相同,擅攀鸟龙科成为唯一属于鸟翼类的非鸟类恐龙。擅攀鸟龙的腰带又具有一些原始特征,例如缺乏完全穿孔的髋臼,比其它兽脚类都要原始,因此反而偶尔被用来反对鸟类的兽脚类起源说。其他研究者则将这些视为幼体特征,认为它们会随着发育而改变。
从2010年代起,随着化石材料的增加和系统发育数据集的扩大,擅攀鸟龙科的位置也发生了改变,更多地被发现位于基干近鸟类(Paraves,即所有接近新鸟亚纲而远离窃蛋龙的手盗龙类),比驰龙科和伤齿龙科更基础。近年,两属新的擅攀鸟龙科奇翼龙(Yi)和混元龙(Ambopteryx)被报道,其研究均支持该科属于基干近鸟类。前者是长约60cm的成年个体,而后者是长约32cm的亚成年或成年个体。它们最突出的特征是腕部具有一延长的骨棍,能在前肢支撑起皮膜,说明其采取了不同于其它恐龙用正羽飞行或滑翔的策略,这可能也是其它擅攀鸟龙科第三指延长和缺乏正羽的原因。关于擅攀鸟龙科的位置还有另一种较常见的观点,认为它们属于基础窃蛋龙类,或者是窃蛋龙类的姊妹群。
伤齿龙科、近鸟龙科与鸟类
在20世纪90年代的系统发育研究中,伤齿龙科偶尔作为鸟类的姊妹群,而一般被认为和驰龙科一起构成恐爪龙类。21世纪初,热河生物群出产了一些原始伤齿龙科,尤其是具有后倾耻骨的中国猎龙(Sinovenator),进一步支持了恐爪龙类的单系性。
2009年,徐星等根据一件产自辽宁建昌的不完整标本命名了近鸟龙(Anchiornis),归入基干鸟翼类,其出产地层的具体年代不明。同年,胡东宇等报道了一件更完整的近鸟龙标本,确认其年代及层位为晚侏罗世早期的髫髻山组,距今约1.61-1.51亿年,早于约1.5亿年前的始祖鸟和约1.25亿年前的热河生物群。这解决了兽脚类起源说的“时间悖论”,即没有确凿地发现过年代早于始祖鸟的有羽毛恐龙。该研究将近鸟龙归入伤齿龙科。
2 010 年代以来,髫髻山组又陆续出产了一些类似近鸟龙的有羽毛恐龙,如晓廷龙(Xiaotingia)和彩虹龙(Caihong),它们的加入使得近鸟类的系统发育关系发生了巨大的动荡。这些恐龙既有类似伤齿龙科的特征,例如齿骨前部的牙齿比中部的更小、更密集,又有类似鸟翼类的特征,例如肩胛骨肩峰的前缘向外弯曲。它们很多也呈现“四翼”形态,但羽毛较小盗龙类的散乱,无明显分层或呈丝状。目前通常认为它们构成单系群,属于近鸟龙科(Anchiornithidae),是鸟类的姊妹群;而伤齿龙科是鸟翼类的姊妹群,比驰龙科更接近鸟类。
近鸟龙科是手盗龙类中最新建立的一个科级演化支,该类群通常还包括产自内蒙古道虎沟的足羽龙(Pedopenna),其属于中侏罗世的海房沟组,与髫髻山组的近鸟龙科同属燕辽生物群。热河生物群的义县龙(Yixianosaurus)和原先被归入始祖鸟的奥斯特伦姆龙(Ostromia)可能也属于这个类群。近鸟龙科被推测起源于亚洲。
此外还存在两种不同的观点,认为髫髻山组的近鸟龙科成员属于伤齿龙科或始祖鸟科。这两种观点通常维持恐爪龙类的单系性,后者常将始祖鸟科也归入恐爪龙类。也有部分研究认为近鸟龙科的成员不构成单系群,而是散布在鸟翼类的不同演化阶段。

较完整的近鸟龙化石标本照片及线图(引自Hu et al.,2009)
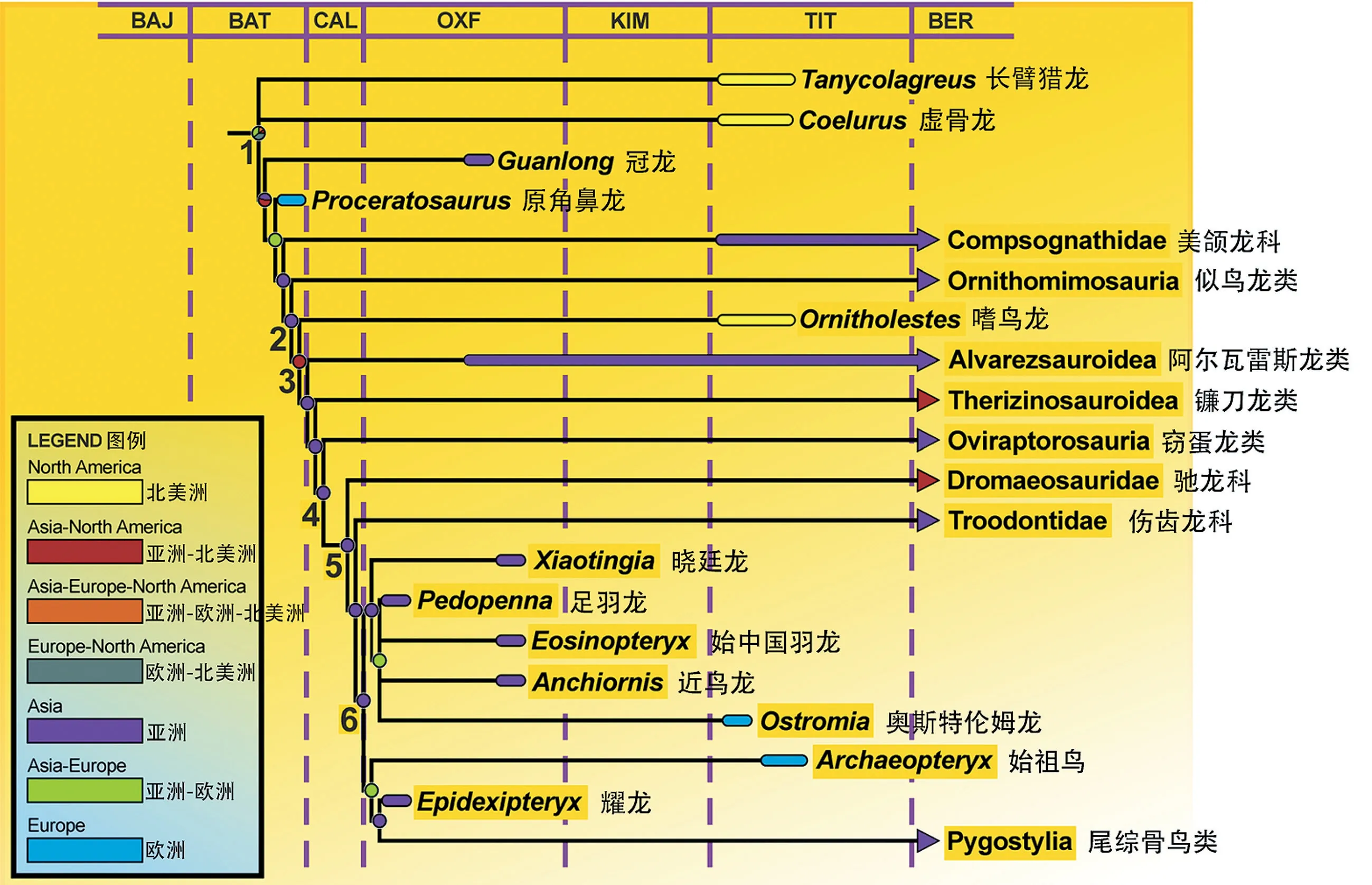
含有近鸟龙科的虚骨龙类分支图(改自Foth & Rauhut,2017)。左下角的图例代表不同支系的起源地。数字代表的节点为:1.虚骨龙类;2.手盗龙形类(Maniraptoriformes);3.手盗龙类;4.廓羽盗龙类(Pennaraptora);5.近鸟类;6.鸟翼类
总结与展望
长久以来,鸟类起源是国内外非常重视的研究课题,是进化论的模式证据之一,在进化生物学中具有重要意义。经历了20世纪后半叶的复兴之后,在最近25年,鸟类的兽脚类起源说由于我国的众多发现获得了爆发性的进展。鸟类的直接祖先从原则上是难以确认的,也无法依靠系统发育分析找到直系的演化关系。但是,我们可以找到鸟类的“姊妹群”,并根据姊妹群的特征来设想鸟类的“祖先类型”。哪种恐龙最接近鸟类也是爱好者很感兴趣的问题。近十年来,在手盗龙类之中,距离鸟类最近的几个类群——擅攀鸟龙科、驰龙科、伤齿龙科的关系经常变动,位置很不稳定。在鸟类姊妹群的候选中,这个位置最近有给予近鸟龙科的倾向,但是还远远未到能给出肯定的答案之时。燕辽生物群是目前唯一出产年代早于始祖鸟的有羽毛恐龙的生物群,是擅攀鸟龙科和大多数近鸟龙科的产地,充满了新发现的潜力。希望将来,这一生物群能带来更多的新发现,令我们更加精确和深入地认识由非鸟恐龙到鸟类的演化过程。
猜你喜欢
学与玩(2022年9期)2022-10-31
文苑(2020年12期)2020-04-13
传播与制作(2019年10期)2019-12-02
中华诗词(2019年7期)2019-11-25
小太阳画报(2019年1期)2019-06-11
小学阅读指南·低年级版(2016年11期)2017-02-06
科技知识动漫(2016年12期)2016-12-23
漫画月刊·哈版(2016年1期)2016-02-22
小学科学(2015年11期)2015-12-01
电影新作(2014年1期)2014-02-27
