
河北菜用大豆田昆虫群落功能团组成及多样性研究
2022-06-22 02:54高慧敏巴义彬冉红凡田玉孙英焘郝玲玉赵思佳曹岩坡
西南大学学报(自然科学版) 2022年6期
高慧敏, 巴义彬, 冉红凡, 田玉,孙英焘, 郝玲玉, 赵思佳, 曹岩坡
1.河北省农林科学院 经济作物研究所/河北省蔬菜工程技术研究中心,石家庄 050051;2.河北大学 博物馆,河北 保定 071000;3.河北省农林科学院 植物保护研究所/河北省农业有害生物综合防治工程技术研究中心/农业部华北北部作物有害生物综合治理重点实验室,河北 保定 071000
菜用大豆(Glycinemax)又称毛豆,以其营养丰富、味道鲜美以及医药保健作用深受广大消费者的喜爱.近年来随着市场需求的增加,菜用大豆产业迅猛发展,我国已成为世界上最大的菜用大豆生产与出口国之一,主要分布在福建、浙江、江苏以及安徽、河北等地[1].然而,规模化生产与连年种植常造成害虫的大发生,威胁菜用大豆产业的发展.多年来,我国生态学家和学者们对不同作物田的节肢动物群落进行了研究[2-5].目前关于菜用大豆田昆虫群落的文献报道仅有节肢动物群落的种类组成[6-7]、菜用大豆田害虫天敌资源调查[8-9]等,而关于北方地区菜用大豆田昆虫群落组成及功能团方面的研究还未见报道.本文利用群落中功能团的多样性替代群落中物种的多样性,通过对河北省菜用大豆田昆虫群落营养结构的多样性变化进行调查,探明其昆虫群落的生物资源现状、群落营养结构及多样性变化,为进一步探讨北方地区菜用大豆田的科学管理与虫害综合防控提供科学依据.
1 材料与方法
1.1 试验地概况
田间调查在河北省农林科学院经济作物研究所大河试验园区菜用大豆田进行(38°05′N,114°18′E),面积2 000 m2.菜用大豆于2018年5月20日播种,10月20日收获.地块前茬为芦笋,同期周边地块种植有大葱、韭菜、甘蓝等蔬菜.
1.2 调查方法
2018年5月15日至10月31日采用马来氏网开展昆虫诱集.将马来氏网安装于菜用大豆田核心区域,要求通风良好且阳光充足.拉紧固定绳索,使其各网面展开抻平;中间的黑色阻隔网下部与地面接触,不留缝隙,以便收集爬行类节肢动物.收集瓶中酒精(95%)持有量保持在50%以上,每隔15 d换1次收集瓶,将收集瓶中的昆虫标本逐一编号进行分类鉴定及统计.在菜用大豆播种后立即挂网开始诱集,5月29日第1次收样,更换收集瓶,此后每隔15 d收样1次.
1.3 功能团划分
按照营养和取食习性将昆虫样本分为吸收式害虫、咀嚼式害虫、天敌昆虫和中性昆虫4个功能团.
个体数量占捕获总量大于10%为优势类群,1%~10%为常见类群,不足1%为稀有类群[10].
1.4 数据处理
数据采用SPSS 23.0进行统计分析,运用R(V3.6.1)软件计算分析以下参数:
1) 物种相对丰盛度(N),为物种个体数占总个体数的比例.
2) 物种丰富度(S),即群落中物种数目的多少.
3) Shannon-Wiener多样性指数(H′)
H′=-ΣPiln(Pi)
式中Pi为第i个科的个体占总个体数的比例.
4) 均匀性指数(E)
E=H/Hmax
式中H为实际观察的物种多样性指数,Hmax为最大的物种多样性指数,Hmax=LnS(S为群落中的总物种数)[11].
5) Gini-Simpson指数(D)
D=Σ(n/N)2
式中n为某一物种个体数,N为总的个体数[12].
群落结构相似性分析:将物种个体数量原始数据进行对数转换,计算Bray-Curtis相似指数,建立各处理间相似性矩阵,采用非参数的二因子相似性分析(ANOSIM)和非度量多维标度(NMDS)排序法分析各功能团的结构差异[13].
2 结果与分析
2.1 昆虫群落组成结构
在菜用大豆的整个生长期共采集到6 888头昆虫,隶属于11目94科255种.根据营养关系与食性分为吸收式害虫、咀嚼式害虫、天敌昆虫及中性昆虫4个功能团(表1).从物种丰富度上看,咀嚼式害虫的物种丰富度最高(104种),占总数的40.79%,其次为吸收式害虫(65种)和天敌昆虫(62种),中性昆虫种类最少(24种).从个体数量上看,吸收式害虫最多,占总数65.82%,其中丝光小长蝽Nysiusthymi个体总数高达1 489头,占吸收式害虫总数的32.84%,占所有诱集昆虫个体数的21.62%.其次,烟蓟马Thripstabaci,蚜虫Aphididaesp.,假眼小绿叶蝉Empoascavitis及条沙叶蝉Psammotettixstriatus等,个体数量在200~700头之间.咀嚼式害虫种类多,但除隆背花薪甲Cortinicaragibbosa外,个体数均较少.中性昆虫物种数与个体数在所有采集到的种类与个体数占比上均低于10%.
此外调查发现62种昆虫为优势种群,个体数占诱集到昆虫总数的91.08%,其余193种昆虫个体数仅占诱集到昆虫总数的8.92%,其中77种昆虫仅诱集到1头,55种昆虫诱集到2~3头,61种昆虫诱集到4~5头.
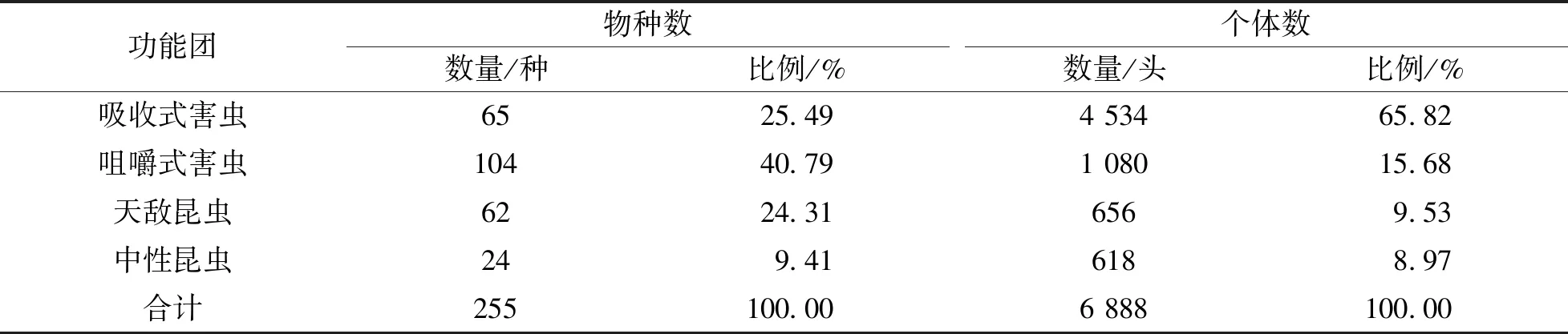
表1 菜用大豆田昆虫功能团物种组成结构
2.2 菜用大豆主要害虫与天敌发生动态分析
在菜用大豆整个生长期有6种主要害虫,包括5种吸收式害虫(丝光小长蝽、烟蓟马、蚜虫、假眼小绿叶蝉、条沙叶蝉)与1种咀嚼式害虫(隆背花薪甲),6种害虫的个体数之和占捕获昆虫总数的51.03%.丝光小长蝽种群数量在6月底达到最高峰,之后下降,至7月底数量有所回升,但之后逐渐递减.烟蓟马与条沙叶蝉自5月底开始至6月底数量逐渐增多,之后虫口数量一直保持在较低水平,至10月中旬近收获季烟蓟马的数量又有较大幅度升高.咀嚼式害虫中,隆背花薪甲数量最多,但与吸收式害虫相比,数量显著低于吸收式害虫(图1).
菜用大豆田主要的天敌以捕食性天敌为主(图2),主要包括龟纹瓢虫Propyleajaponica,印度细腹食蚜蝇Sphaerophoriaindiana,东亚小花蝽Oriussauteri,十二斑褐菌瓢虫Vibidiaduodecimguttata,日本通草蛉Chrysoperlanippoensis,深点食螨瓢虫Stethoruspunctillum.其中龟纹瓢虫、印度细腹食蚜蝇和东亚小花蝽数量最多.龟纹瓢虫在整个生长季节的发生数量比较稳定,收获后其数量减至0;印度细腹食蚜蝇的发生高峰期出现在6月中旬;东亚小花蝽的发生高峰期出现在6月下旬;十二斑褐菌瓢虫主要发生在菜用大豆生长中后期(8月中旬至9月中旬);深点食螨瓢虫主要发生在菜用大豆生长中期(7月中旬至9月中旬);日本通草蛉在菜用大豆生长前期(6月中、下旬)出现1个小高峰期,而盛发期则出现在中后期(8月下旬).
寄生性天敌白毛长腹土蜂Campsomeriellaannulata和蚜茧蜂Aphidiussp.发生量相对较大,分别活跃在8月和7月中上旬.
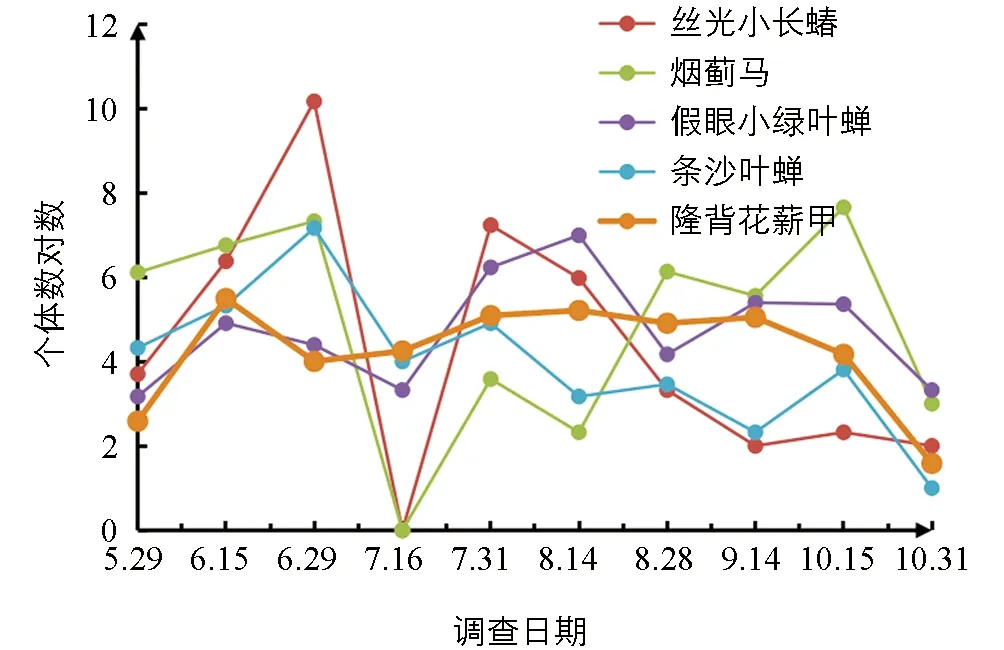
图1 菜用大豆主要害虫发生动态

图2 菜用大豆主要天敌发生动态
2.3 菜用大豆田昆虫各功能团物种相对丰盛度与物种丰富度的时间动态
在菜用大豆整个生长期内,除8月底至9月中旬外,其他各时期吸收式害虫的相对丰盛度均最高,分别在6月中旬至7月底和10月中旬出现两个高峰.咀嚼式害虫相对丰盛度在8月底至9月中旬最高.天敌昆虫的相对丰盛度稳定在较低水平.中性昆虫除在7月中旬出现1个高峰外,其余时间均维持在较低水平(图3).
各功能团的物种丰富度显示(图4),吸收式害虫和咀嚼式害虫的物种丰富度高于天敌昆虫与中性昆虫.吸收式害虫的物种丰富度在6月保持较高水平,之后略有降低并维持在中等水平,至9月中旬后迅速降低.咀嚼式害虫的丰富度在6月中旬和9月中旬出现两个峰值,之后锐减.天敌昆虫在整个生长前中期波动都不大,在9月中旬之后迅速降低.中性昆虫仅在7月中旬出现了1个高峰,其余时间均维持在较低水平.
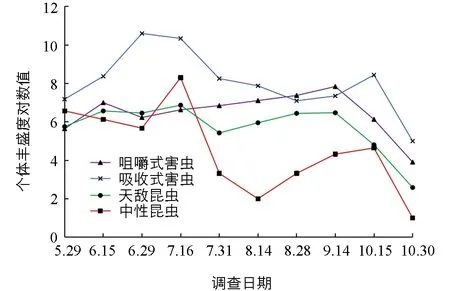
图3 菜用大豆田昆虫各功能团物种相对丰盛度时间动态

图4 菜用大豆田昆虫各功能团物种丰富度指数时间动态
各功能团的物种Shannon-Wiener多样性指数显示(图5),在菜用大豆生长期,咀嚼式害虫的多样性多数时间高于其他3个功能团,保持在1.6~3.0之间.吸收式害虫在生长前期多样性指数较高(1.9),之后呈现出下降—升高—下降的趋势.天敌昆虫在菜用大豆生长前中期呈波浪式上升,维持在1.6~2.5之间,10月中旬之后迅速下降.中性昆虫多样性指数高峰出现在7月中旬,之后迅速降低,8月中旬为0,之后迅速升至1.4,9月至10月持续下降.
菜用大豆昆虫各功能团Gini-Simpson集中性指数分析显示(图6),咀嚼式害虫集中性指数在5月底至10月底均较高(0.7~0.9).吸收式害虫6月中旬优势集中性最高,之后迅速降低,7月中下旬有所回升并维持至9月中旬.天敌昆虫则呈波浪式上升,维持在0.6~0.9之间,10月中旬之后迅速降低.中性昆虫波动幅度较大,7月中旬之后迅速降低,8月中旬为0,8月底反弹至0.7,之后逐渐降低,10月底降至0.5.
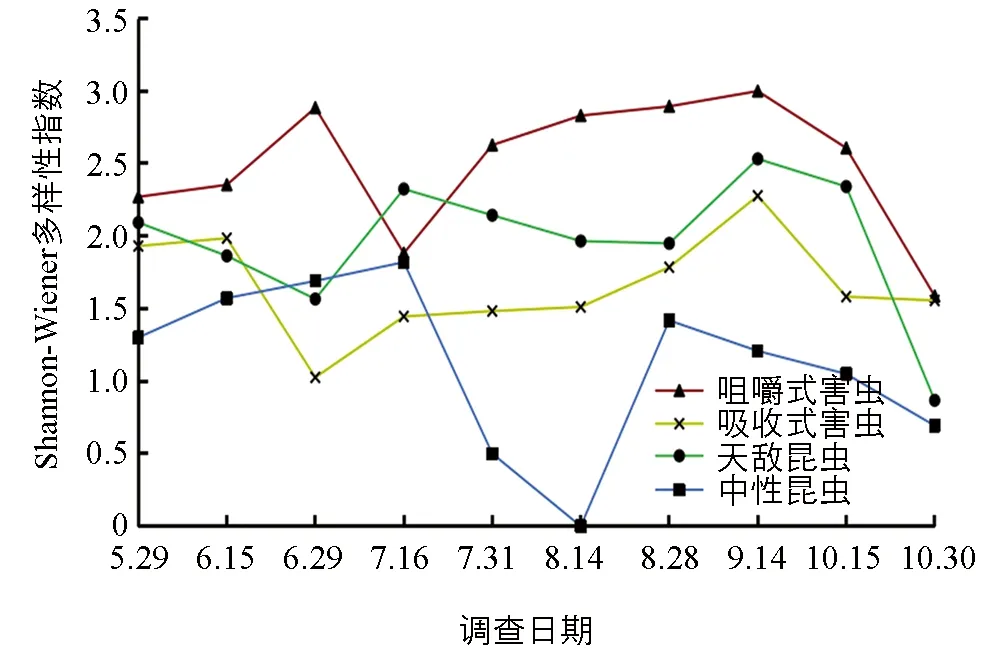
图5 菜用大豆田昆虫各功能团Shannon-Wiener多样性指数时间动态
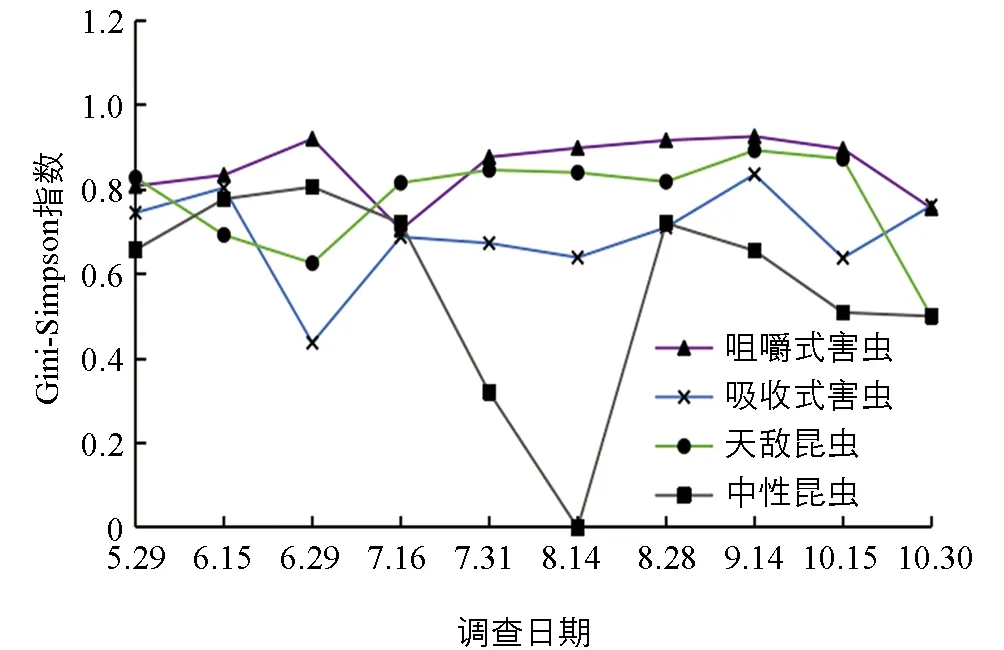
图6 菜用大豆田昆虫各功能团Gini-Simpson集中性指数时间动态
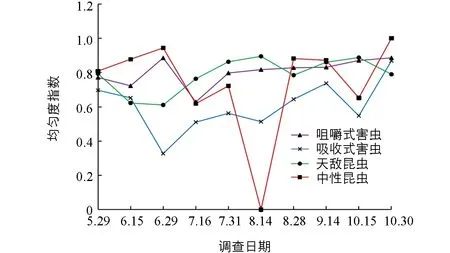
图7 菜用大豆田昆虫各功能团均匀度指数(Pielou)时间动态
菜用大豆田昆虫各功能团均匀度指数时间动态分析显示(图7),咀嚼式害虫的均匀度指数在整个生长期维持在较高水平(0.7~0.9).吸收式害虫的均匀度指数较低,且呈波浪式上升.天敌昆虫在5月底至10月中旬维持在较高水平(0.6~0.9).中性昆虫波动较大,在7月底之前均维持在较高水平(0.7~0.9),之后急剧下降,8月中旬至0,之后迅速上升—下降—上升,10月底上升至最高水平(1.0).
菜用大豆昆虫各功能团NMDS二维点图胁迫系数(stress)均小于0.2.由NMDS二维点图可知,吸收式害虫、咀嚼式害虫及中性昆虫种类呈离散式分布,而天敌昆虫则有较明显的聚群划分(图8).
3 结论与讨论
生物多样性是指在一定地区所有生物物种及其遗传变异和生态系统的复杂性总称.通过分析群落结构多样性,比较该地区害虫群落结构及天敌种类与数量,昆虫多样性研究已从物种多样性拓展至功能多样性[14].利用昆虫群落中功能团多样性替代物种多样性,可以使复杂的网络结构简单化,不同类群间的关系更加清晰[15-16].本研究采用马来氏网诱集,调查分析了河北石家庄菜用大豆田主要害虫、天敌类群与发生动态.共采集到6 888头昆虫标本,分属11目94科255种,分为吸收式害虫、咀嚼式害虫、天敌昆虫与中性昆虫4个功能团.吸收式害虫中以丝光小长蝽、烟蓟马、蚜虫、假眼小绿叶蝉与条沙叶蝉为害最为严重.咀嚼式害虫种类最多,但仅隆背花薪甲发生数量较多,可能引起较大为害.天敌昆虫则以捕食性的龟纹瓢虫、印度细腹食蚜蝇、东亚小花蝽、十二斑褐菌瓢虫、日本通草蛉、深点食螨瓢虫等为主,其中龟纹瓢虫、印度细腹食蚜蝇与东亚小花蝽数量总体最多;寄生性天敌蚜茧蜂与白毛长腹土蜂数量也相对较多.与其他研究相比,本研究获得的节肢动物物种数更多,这可能与试验地周围的植被更加丰富有关[9,17].

图8 菜用大豆田昆虫各功能团NMDS二维点图
本研究首次发现丝光小长蝽为菜用大豆上的重要害虫,在6月中下旬种群数量急剧增加,这与此前仅有零星分布的报道存在一定差异[18],其对菜用大豆的危害与防治有待进一步研究.此外,烟粉虱在河北与福州均为菜用大豆重要害虫,而在福州危害严重的斜纹夜蛾ProdenialituraFabricius和豆银纹夜蛾AutographanigrisignaWalker等鳞翅目害虫在河北地区仅为零星发生[8].本研究中蚜虫、假眼小绿叶蝉、烟蓟马、烟粉虱种群数量较多,这与山东、长春等地的菜用大豆上的主要害虫种类报道相一致[19-23].此前已报道的菜用大豆田天敌中瓢虫科、食蚜蝇科、小蜂总科等的一些种类为优势种[8],本研究中获得的天敌仅为昆虫纲5目62种,以捕食性的龟纹瓢虫、印度细腹食蚜蝇、东亚小花蝽、十二斑褐菌瓢虫、日本通草蛉、深点食螨瓢虫等种类为主,蛛形纲天敌有待调查.龟纹瓢虫、东亚小花蝽、日本通草蛉等捕食性天敌为全国广布种,其他种类仅在河北为优势种.
由于气候条件、耕作制度以及周边田块植被不同,导致南北方菜用大豆田主要害虫与天敌类群存在较大差异.石家庄菜用大豆田主要害虫种类较少,这可能是因为周边植被丰富、整体物种多样性较高、群落结构稳定等环境条件决定的.本研究明确了石家庄菜用大豆主要害虫与天敌类群,为当地有针对性地制定有效的害虫防治策略提供了理论与指导依据.
猜你喜欢
煤气与热力(2021年5期)2021-07-22
红蜻蜓(2020年9期)2020-09-26
机电信息(2020年19期)2020-08-31
发电技术(2020年3期)2020-06-29
落叶果树(2020年2期)2020-04-20
小学生作文(低年级适用)(2018年4期)2018-11-29
小朋友·快乐手工(2015年10期)2015-11-02
中国新闻周刊(2015年31期)2015-08-28
小朋友·快乐手工(2015年2期)2015-03-13
科技创新与品牌(2012年7期)2012-08-06
