
一个矮穗位自交系的主基因+多基因混合遗传模型
2022-06-21 09:30颜勇刚陆江江瑞林李鸿钰程俐
四川农业科技 2022年5期
颜勇刚,陆江,江瑞林,李鸿钰,程俐
(四川省乐山市农业科学研究院,四川 乐山 614000)
玉米不仅是我国主要的粮食作物、饲料作物,还是淀粉和乙醇生产企业的重要原料[1]。随着玉米种植密度的提高,株型改良已成为玉米遗传育种工作者研究的重点内容之一[2]。穗位高是玉米理想株型育种的一个重要性状[3]。穗位高性状为多基因控制的数量性状,近年来借助分子标记连锁图谱应用数量性状位点(QTL)分析方法对玉米穗位高性状的遗传进行了较多研究,但是研究结果不尽一致[4]。穗位高对玉米茎和根生长有重要影响,穗位过高迫使根和茎承受更大的压力,容易造成植株倒伏[5]。经典数量遗传学在分析遗传规律时存在一定的局限性,近些年植物数量性状主基因和多基因混合分析逐渐被人们认识并且广泛的应用于各种作物,可检测和鉴定数量性状主基因和多基因的存在,并可对基因效应和方差等遗传参数进行估计[6]。本文应用植物数量性状主基因+多基因混合遗传模型分析方法对一个矮穗位自交系ds1的穗位高进行了遗传分析,为进一步利用分子标记对穗位高进行有针对性的选择和改良提供理论依据。
1 材料和方法
1.1 供试材料
自交系ds1、As、Bs、Cs均为乐山市农业科学研究院的玉米自选系。2019年春季,于四川乐山配置3个杂交组合As×ds1、 Bs×ds1及Cs×ds1。2020年春季,利用3个F1与亲本回交获得三个组合的BC1和BC2,3个F1自交获得F2。2020年冬季和2021年春季将各组合用于遗传分析的6个世代种植于四川乐山和海南三亚,常规大田管理。
1.2 性状调查
待雄穗停止生长,测定所有群体所有植株的穗位高,即地面到第一果穗柄着生节的高度。
1.3 数据分析
应用章元明等[7]和盖钧镒等[6]提出的植物数量性状“主基因+多基因混合模型”分析方法,对各组合6世代穗位高进行分析。通过AIC(Akaike.s information criterion)值的判别和一组适合性测验选择最优遗传模型,并估计主基因和多基因的效应值、方差等遗传参数。
2 结果与分析
2.1 各世代穗位高的次数分布
应用多世代联合分析数量性状主基因和多基因混合遗传的统计方法,分析了玉米3个杂交组合6个世代穗位高的遗传效应。四川乐山,母本自交系As、Bs、Cs的平均穗位高分别为为41.05、39.44、54.75cm,父本ds1的平均穗位高为35.45cm,Cs、As、Bs F1代的平均穗位高分别为64.54、59.05、73.31cm。海南三亚Cs的平均穗位高为84.38cm,ds1的平均穗位高为45.96cm,其F1代的平均穗位高为110.75cm。各组合的F2穗位高平均值分别为53.36、49.09、67.43、91.29cm。
从3个组合4个世代的穗位高分布和拟合曲线(图1)可以看出,3个组合的BC1、F2群体的穗位高呈双峰分布,有明显的主基因存在特征。
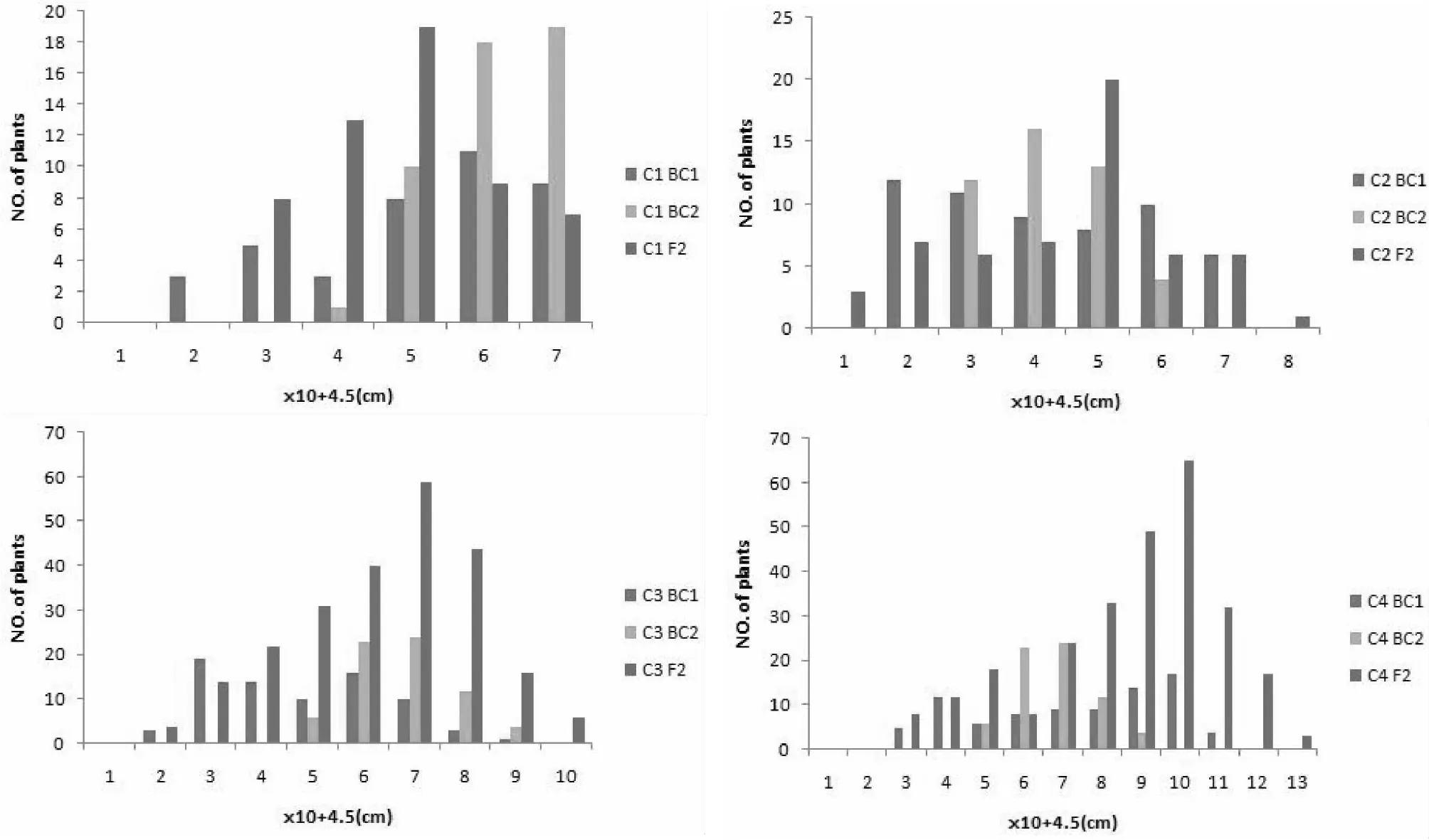
C1 = As×ds1 (乐山),C2 = Bs×ds1 (乐山),C3= CS×ds1 (乐山),C4 = Cs×ds1(三亚)图1 各组合BC1、BC2、F2穗位高的次数分布注:下文均用C1、C2、C3、C4代替各组合
2.2 遗传分析
2.2.1 遗传模型 利用植物“主基因+多基因混合遗传模型”分析C1、C2、C3、C4组合F2群体的穗位高,获得了5类模型的极大似然值和AIC值。其中3个组合的穗位高均是D2、D4、D1模型的AIC值最小(表1)。通过对D2、D4、D1模型的适合性检验(表2)和AIC值分析,结果显示3个组合的3个模型的所有分布参数与理论分布一致(P>0.05),而D2模型的AIC值最小,即D2模型为最优,比较符合F2群体的实际分布,即3个组合的穗位高均为“1对加性主基因+加性-显性多基因混合”遗传模式。
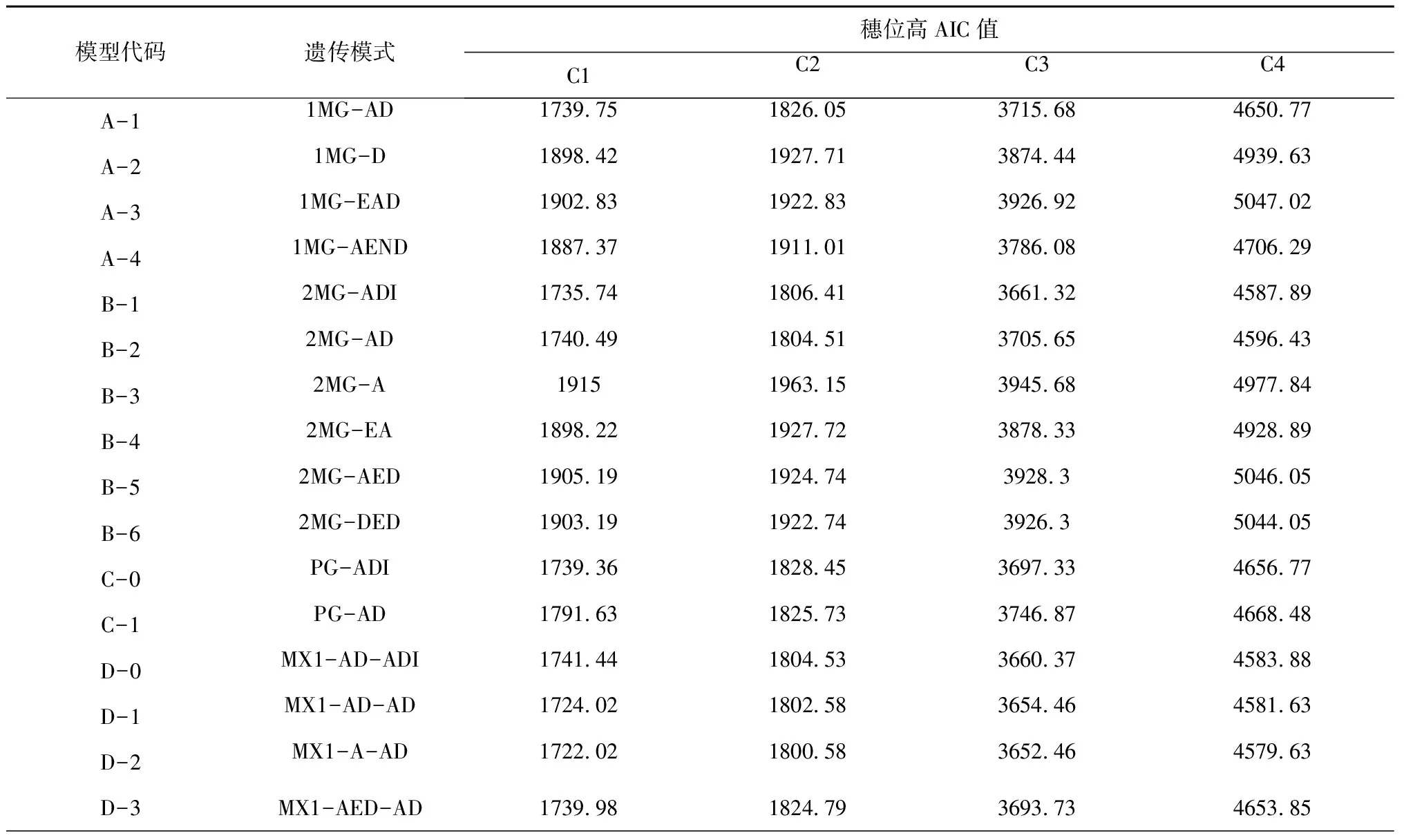
表1 各组合后代穗位高遗传模型的AIC值
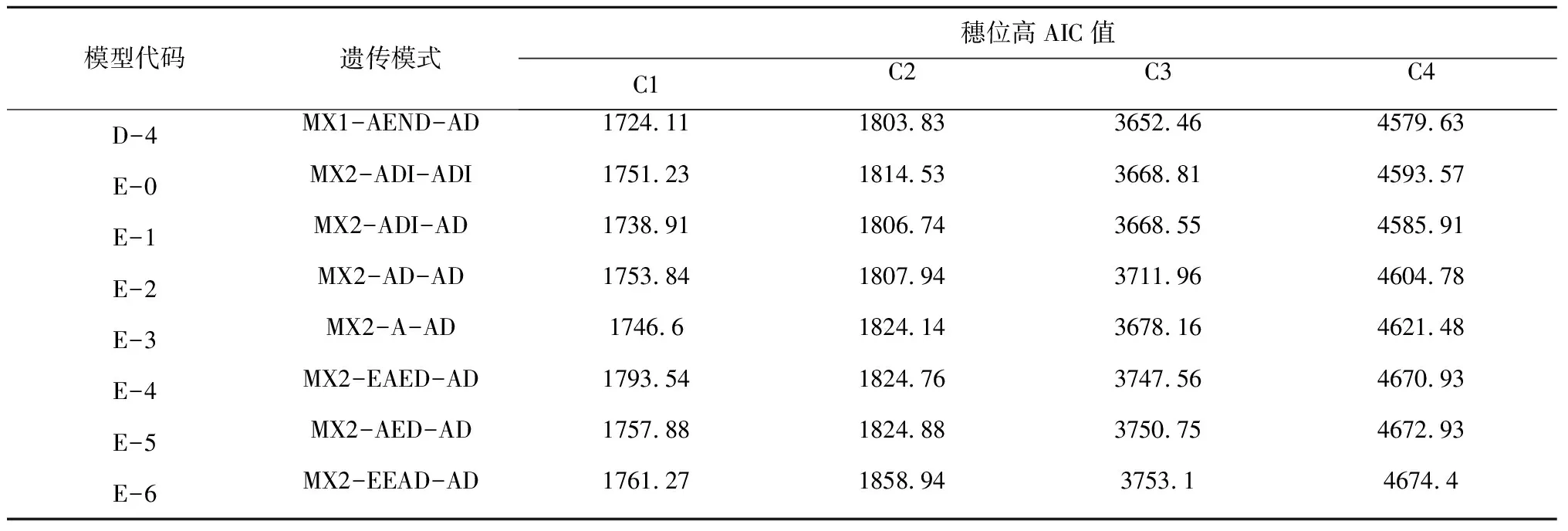
续表1
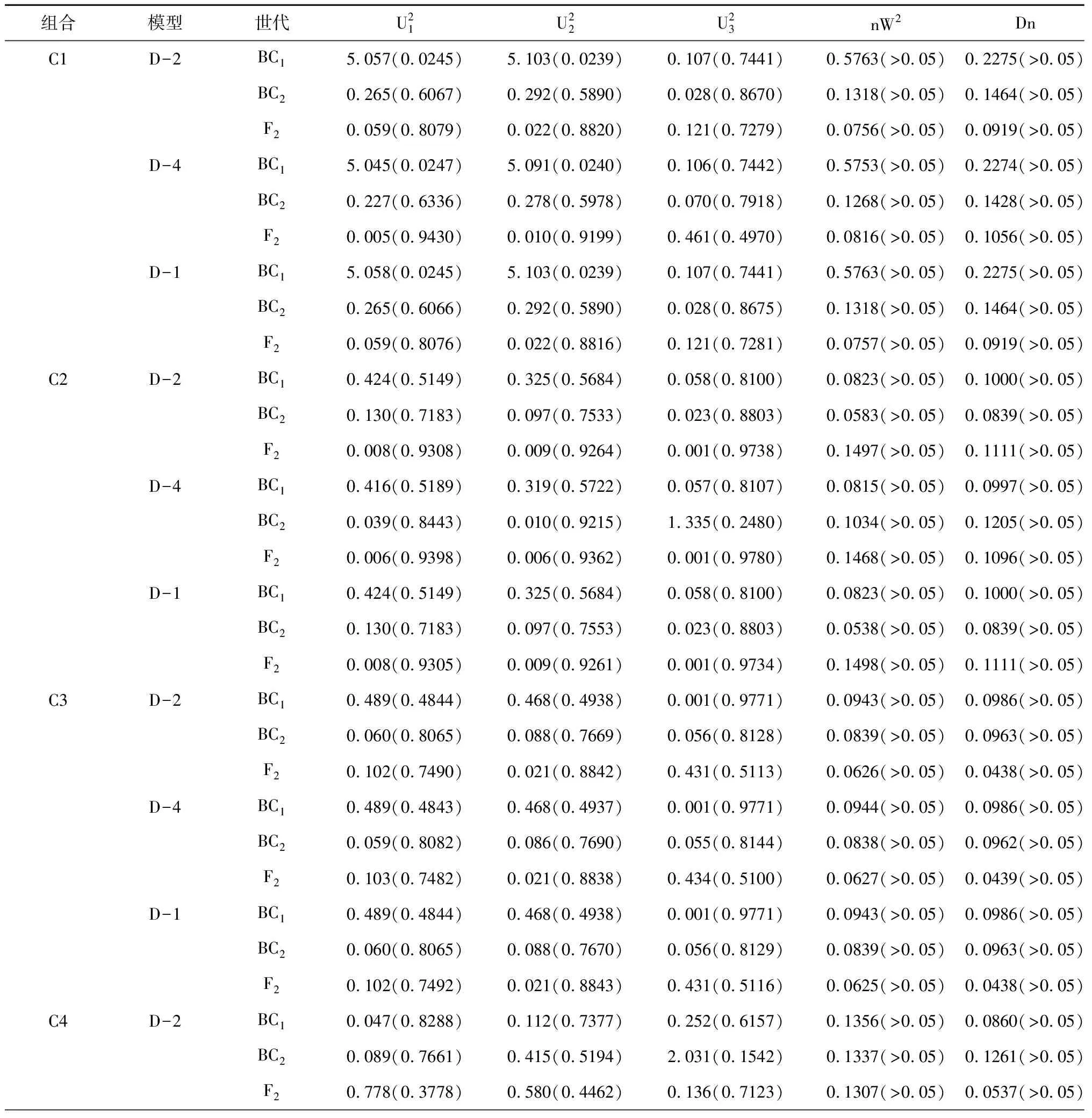
表2 各组合后代穗位高备选遗传模型的适合性检验
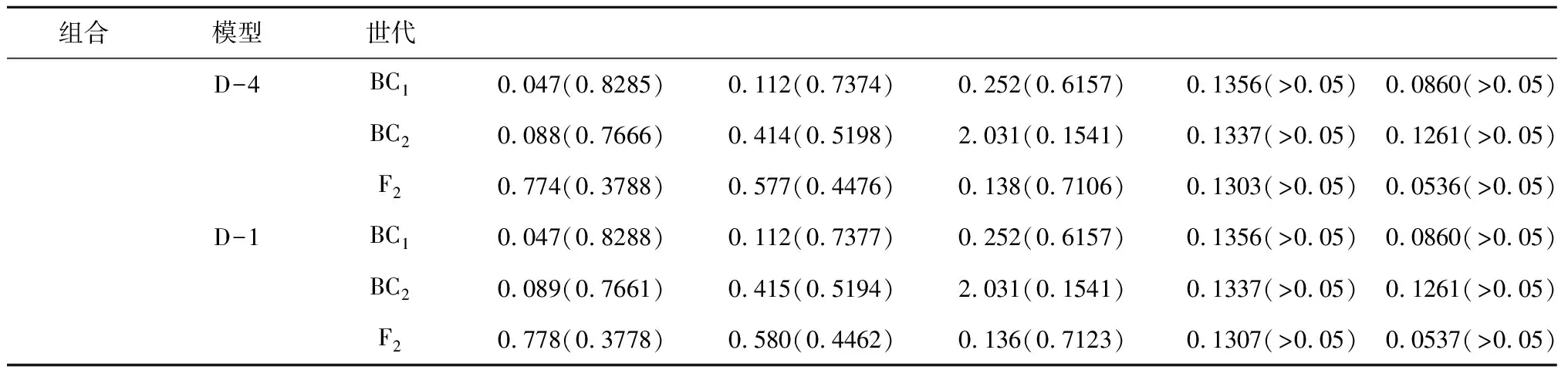
续表2
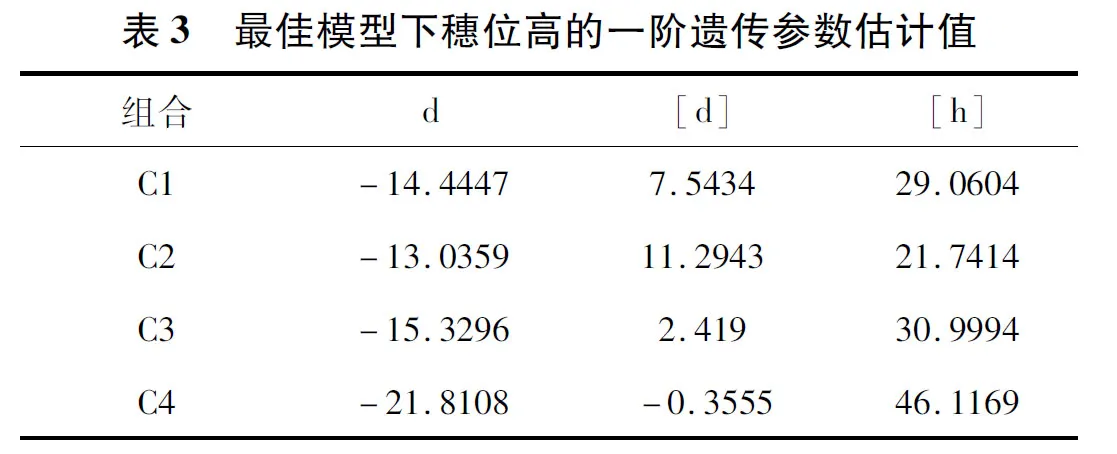
2.2.2 遗传参数的估算 根据选择的最优遗传模型估计穗位高最优遗传模型一阶遗传参数(表3)。组合C1、C2、C3、C4控制穗位高的主基因加性效应分别为-14.4447、-13.0359、-15.3296、-21.8108,多基因加性效应为7.5434、11.2943、2.419、-0.3555,多基因显性效应为29.0604、21.7414、30.9994、46.1169。
根据选择的最优遗传模型估计穗位高最优遗传模型二阶遗传参数(表4)。C1组合BC1、BC2和F2群体的主基因遗传力分别为20.68、76.52、76.74,而多基因遗传力则分别为4.89、0、0,表明BC2和F2群体主基因分别决定了穗位高表型变异的76.52%、76.74%,而多基因决定了穗位高表型变异的0%、0%。C2组合BC1、BC2和F2群体的主基因遗传力分别为10.6、31.62、67.03,而多基因遗传力则分别为23.85、31.84、40.79,表明BC2和F2群体主基因分别决定了穗位高表型变异的31.62%、67.03%,而多基因决定了穗位高表型变异的31.84%、40.79%。C3组合BC1、BC2和F2群体的主基因遗传力分别为20.15、41.05、39.83,而多基因遗传力则分别为6.8、29.11、27.7,表明BC2和F2群体主基因分别决定了穗位高表型变异的41.05%、39.83%,而多基因决定了穗位高表型变异的29.11%、27.7%。C4组合BC1、BC2和F2群体的主基因遗传力分别为20.55、54.96、44.4,而多基因遗传力则分别为7.36、0、17.35,表明BC2和F2群体主基因分别决定了穗位高表型变异的54.96%、44.4%,而多基因决定了穗位高表型变异的0%、17.35%。由此可见,各组合的主基因表现相似,而多基因表现差异较大,环境可能对穗位高的影响较大。
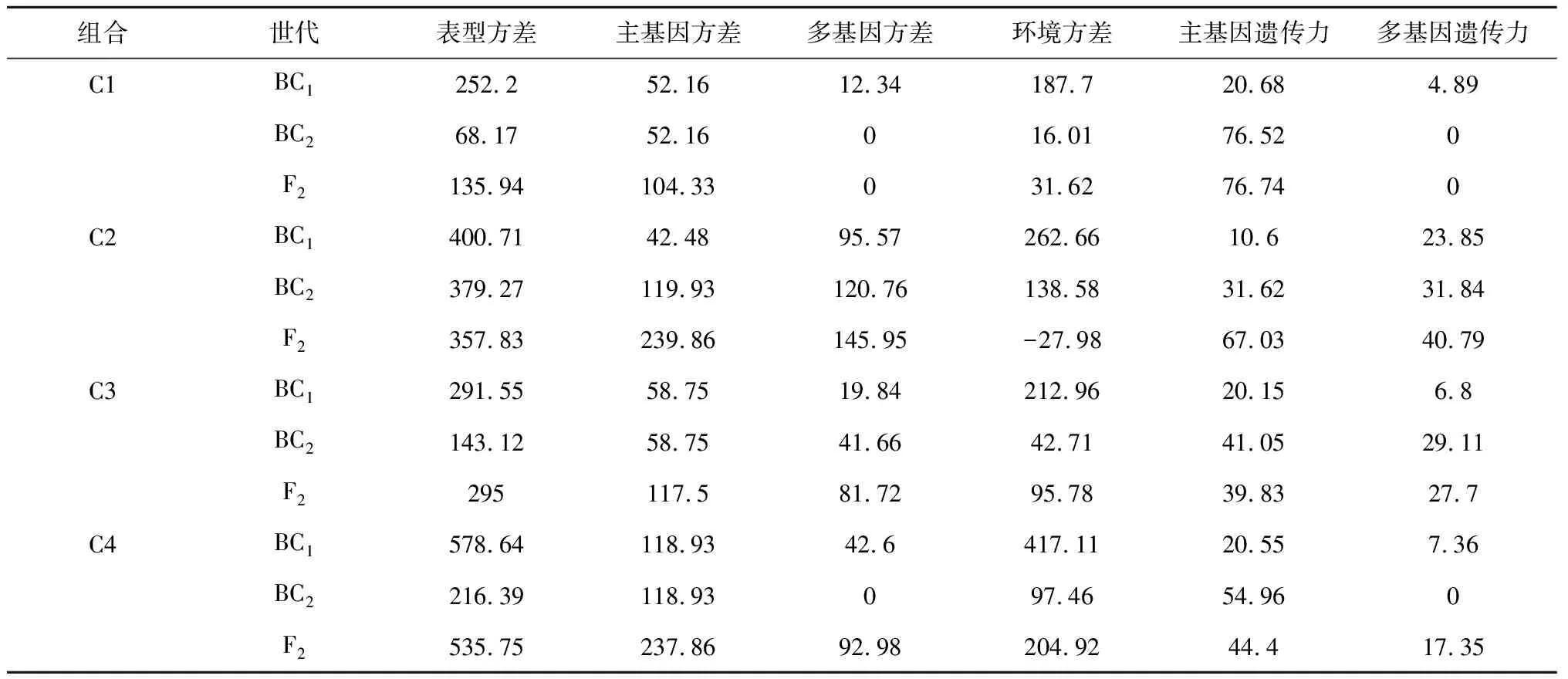
表4 最佳模型下穗位高的二阶遗传参数估计值
3 讨论
经典数量遗传学的研究方法不能检测主基因的存在[8]。本文采用数量性状的主基因+多基因混合遗传模型的方法分析了3个玉米组合穗位高的遗s传,发现穗位高在各组合中均表现为1对加性主基因+加性-显性多基因遗传模型。本研究结果显示,如果对穗位高性状改良,可利用ds1主基因遗传率相对较高的特点,选用所需的穗位高性状,并通过杂交、回交转移主基因,利用主基因和多基因的加性效应来对穗位高材料进行选育。自交系ds1和不同的自交系杂交的组合主基因遗传力和多基因遗传力差距较大,各组合穗位高变异除受主基因和多基因遗传力控制外,还有部分未解释的变异,因此育种过程中对穗位进行选择还需考虑组合、环境等的影响。后期可通过扩大种植群体获得更精确的数据,并利用分子标记和QTL定位进一步研究玉米穗位高的遗传为育种提供参考。
猜你喜欢
作物学报(2022年2期)2022-11-06
西南农业学报(2022年7期)2022-09-30
中国现代医生(2022年21期)2022-08-22
中国种业(2022年8期)2022-08-19
江苏农业科学(2022年6期)2022-04-15
瞭望东方周刊(2021年6期)2021-03-30
支部建设(2020年15期)2020-07-08
三农资讯半月报(2020年2期)2020-03-09
农民致富之友(2016年5期)2016-10-21
百科知识(2015年18期)2015-09-10
