
不同底质及光照条件对轮叶黑藻组织培养及种植的影响
2022-06-18 09:15:42张靖华
生物学杂志 2022年3期
吕 喆,刘 杰,张靖华,万 云,冷 欣
(1.南京大学 生命科学学院,南京 210023;2.安徽建筑大学 环境与能源工程学院,合肥 230601)
沉水植物作为水生态系统的重要生产者,具有重要的生态学功能与意义。已有研究发现,沉水植物能有效吸收储存水环境中的营养物质,影响水域中的物质循环[1-2];缓解水体富营养化,抑制浮游植物生长,减少藻类大暴发,为其他水生植物的生存和生长提供条件[3-4];还能有效富集重金属及悬浮颗粒[5];从而支撑整个水域的生物群落和食物链。大量研究表明,恢复重建沉水植物,可改善水质,提高生态多样性。
轮叶黑藻(Hydrillaverticillate)是水鳖科(Hydrocharitaceae)的一种多年生沉水植物,普遍生长于世界各处水域中,具有极强的耐污性,同时自身的净化能力又使其成为水生植物恢复过程中的先锋物种[6]。众所周知,植物组织培养作为一项被广泛使用的技术,能够不受时间和空间的限制,通过筛选优质种苗,快速产出大量同样高品质的种苗,目前国内已有轮叶黑藻组培的相关方法,从培养基配比、外植体处理、芽诱导增殖及驯化等方面进行了系统的研究,当下的培养方法是使用MS培养基,置于光照度为1 000 lx的白光下,每天照射10 h[7]。而光照这一关键环境因素可分为光强、光质和光周期3个方面,对植物的组织分化、生长发育、形态构成、物质积累等有重要影响。
本实验设置不同光质和光周期进行轮叶黑藻的组织培养,探究不同光照条件对轮叶黑藻组织培养的影响,寻找最适培养光照条件。为了验证培养出的轮叶黑藻种苗质量,使其能够在环境各异的河流湖泊中因地制宜地种植,还设置不同的遮光程度模拟不同水深,搭配不同的底质类型探讨其对轮叶黑藻生长的影响。针对研究中关于底质和水深对植物生长影响结论不一[8],本文通过探究轮叶黑藻组培苗在不同底质与水深下生长表现,探寻不同条件下的最佳种植方式。
1 材料与方法
1.1 材料
为探究光质及光周期对轮叶黑藻组织培养的影响,选用在实验室中继代培养一年的成熟健壮轮叶黑藻作为母本,在超净工作台上剪取健壮的片段置于盛有MS培养液的锥形瓶内,封上封口膜后置于组培室培养,光照周期8 h/d,室内温度(25±1)℃,培养期间随时剔除污染以及长势不佳的植株,培养一个月后的植株留作本次实验的种苗。
为探究底质类型和遮光程度对轮叶黑藻组培苗野外生长的影响,选取在实验室中同批培养的成熟健壮组培苗剪为长(2±0.2)cm,重(0.2±0.01)g的片段,置于营养液中备用。泥土和沙子均取自常熟市南湖湿地公园,种植用水是南湖水,水质指标为氨氮0.05 mg/L,总氮1.1 mg/L,COD 17 mg/L,总磷0.25 mg/L。
1.2 方法
为探究光质及光周期对轮叶黑藻组织培养的影响,采用双因素交叉设计,设置白光(W)、红光(R)、蓝光(B)、红蓝1∶1混合光(R&B)4种光质(白光为全光谱,红光波长658 nm,蓝光波长444 nm),8 h光照/16 h黑暗、12 h光照/12 h黑暗、16 h光照/8 h黑暗3种光照周期(以下简称8 h/d、12 h/d、16 h/d),共计12种光照处理,设置光量子通量均为50 μmol/m2·s;在无菌条件下将种苗植株均匀剪为2 cm左右的片段,每个片段上带有6片左右的叶片,每瓶培养液中接入一个片段,置于不同的光照处理下培养。
为探究底质类型和遮光程度对轮叶黑藻组培苗种植的影响,设置泥土、沙子、泥沙混合(1∶1)3种底质类型,装入一次性种植袋中,底质厚度10 cm,每个种植袋中种入3株轮叶黑藻组培苗片段,用遮阳网设置不遮光、40%遮光和80%遮光,共计9种条件,每种条件设置27组重复,分别置于3个1 m×1 m×1 m的大水箱中,水深保持1 m,每隔10天换水一次。
1.3 测定指标及方法
从培养的第15天起,每隔10 d每种处理随机选取两瓶组培苗,一瓶记录茎长后在烘箱内80 ℃烘至恒重,测定干重;一瓶采用比色法测定叶绿素含量,每个处理设置3个重复。至第55天时,多选取两瓶组培苗,一瓶采用蒽酮法测定植物可溶性总糖含量,采用考马斯亮蓝比色法测定植物可溶性蛋白含量;一瓶使用北京索莱宝科技有限公司的试剂盒测定SOD、POD、CAT活性及MDA含量。酶活力单位定义如下:SOD:在黄嘌呤氧化酶偶联反应体系中样本的抑制百分率为50%时,反应体系中的SOD酶活力定义为一个酶活力单位;POD:每g组织在每mL 反应体系中每分钟470 nm下吸光值变化0.01定义为一个酶活力单位;CAT:每g组织每分钟催化1 μmol H2O2降解定义为一个酶活力单位。
从种植的第10天起,每隔10 d每种处理随机取出9个种植袋,每袋中随机选取一株轮叶黑藻测量其株高、根数、芽数,而后分成根、茎、叶置于烘箱中80 ℃烘至恒重,并计算根、茎、叶的干重占比,至第30天共计3次。
1.4 数据处理
数据处理使用SPSS、R语言等统计软件进行双因素协方差分析和Bonferroni多重比较法。
2 结果与分析
2.1 光质及光周期对轮叶黑藻组织培养的影响
2.1.1 光质和光周期对轮叶黑藻组培苗干重和茎长的影响
在培养前期(15 d和25 d)生长速度缓慢,人工接种初始片段的误差对干重结果干扰严重,从第35天开始趋向正常,故在干重分析中15 d和25 d数据不纳入分析。从第35天开始,表示红光的红色散点和表示12 h/d光照周期的三角形散点处于顶部位置,表明其干重较大;同样,表示蓝光的蓝色散点和表示8 h/d光照周期的圆形散点基本位于底部,表明其干重较小(图1)。

图1 光照条件对轮叶黑藻组培苗干重对数的影响
为满足正态性和方差齐性的要求,对干重进行以10为底的对数变换。根据方差分析结果可知:轮叶黑藻组培苗干重在不同光质及光周期下有显著性差异(P<0.01);茎长受光周期影响极显著(P<0.01),而受光质影响不显著,光质和光周期的协同作用影响不显著(表1)。

表1 不同光照条件下轮叶黑藻组培苗干重和茎长的方差分析结果
红光处理下轮叶黑藻的干重均显著大于其他光质,其他光质之间没有差异;光周期对干重的影响在不同时间、不同光质下表现不一,在第35天,干重在白光和红光下为12 h/d>16 h/d>8 h/d,在蓝光和红蓝混合光下则为16 h/d>12 h/d>8 h/d;在第45天,白光、红光和蓝光下12 h/d的干重显著高于其他两组,红蓝混合光下3种光周期之间没有显著差异;在第55天,白光下12 h/d的干重大于其他两组,蓝光下3种光周期之间没有显著差异,红光和红蓝混合光下12 h/d和16 h/d之间没有差异,均远大于8 h/d(图2)。

不同字母表示存在显著差异,显著水平(P<0.05)。
12 h/d光周期下轮叶黑藻茎长要显著大于其他两组,在第15天到第45天,8 h/d和16 h/d光周期下茎长没有显著差异,但16 h/d光周期呈现反超8 h/d的趋势,至第55天,16 h/d光周期下茎长显著大于8 h/d(图3)。
2.1.2 光质和光周期对轮叶黑藻组培苗叶绿素含量的影响
方差分析结果显示,光质和光周期均显著影响叶绿素a和叶绿素b的含量;而光质和光周期及其交互作用显著影响了叶绿素a和叶绿素b的比值。轮叶黑藻组培苗叶绿素含量在不同光照条件下的差异主要是由光周期引起的(表2)。
12 h/d光周期下叶绿素a、叶绿素b的含量远高于其他两组。对于叶绿素a来说,从第35天开始,蓝光下的叶绿素a含量均最高,而光周期则基本呈现12 h/d>8 h/d>16 h/d;而对于叶绿素b来说,在第15天到第35天,白光照射下含量高于其他光照,而从第45天开始各组光质之间均没有显著差异,8 h/d和16 h/d光周期下,叶绿素b含量无显著差异。蓝光照射下叶绿素a/b最大,红蓝混合光次之;对于光周期而言,12 h/d的叶绿素a/b最低,而8 h/d和16 h/d的叶绿素a/b没有显著性差异(图4)。
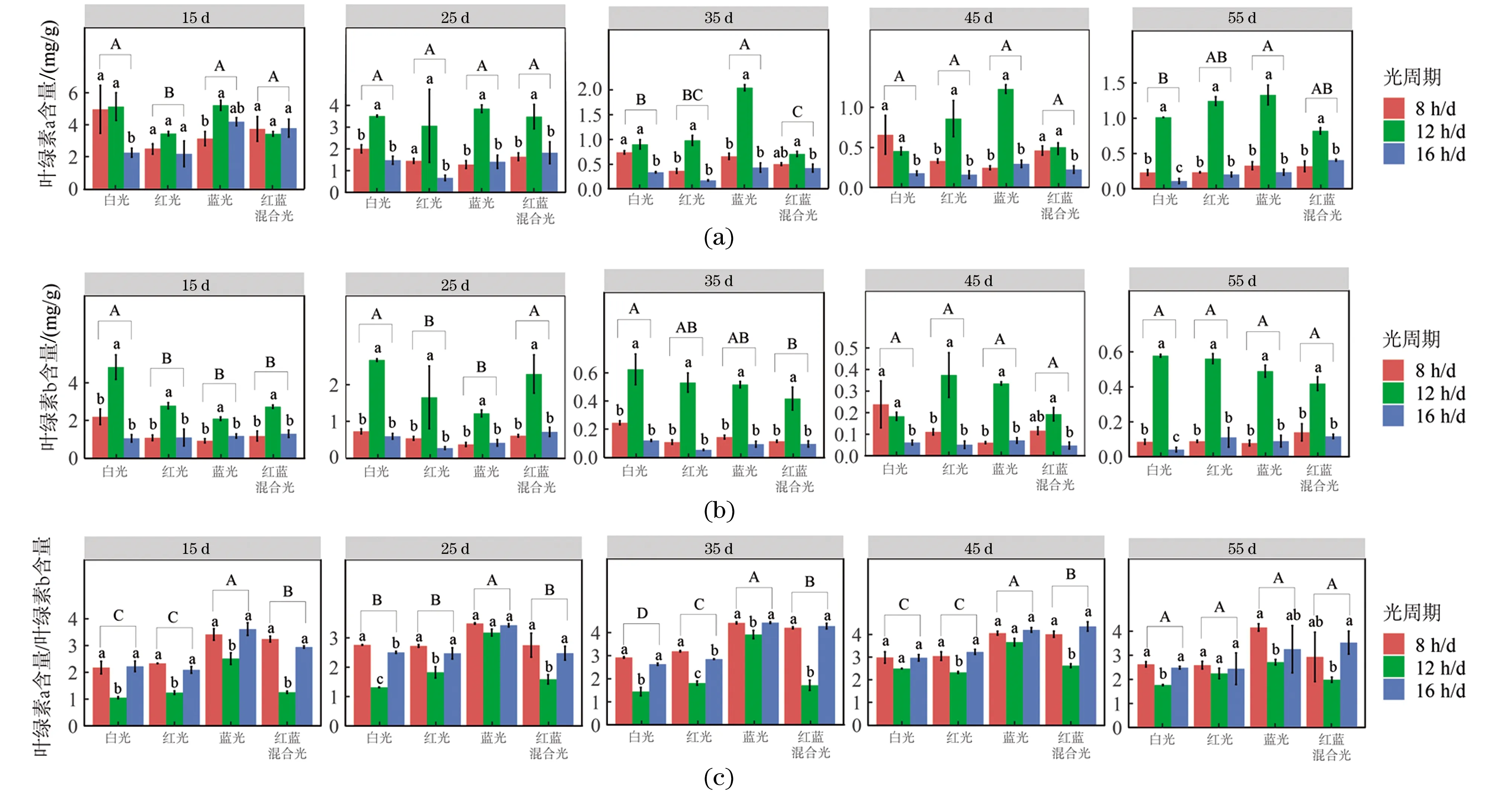
(a)叶绿素a含量多重比较;(b)叶绿素b含量多重比较;(c)叶绿素a、b含量比值多重比较。不同字母表示存在显著差异,显著水平(P<0.05)。
2.1.3 光质和光周期对轮叶黑藻组培苗可溶性糖和可溶性蛋白含量的影响
方差分析结果显示:光质和光周期及其交互作用显著影响轮叶黑藻组培苗可溶性糖和可溶性蛋白的含量。轮叶黑藻组培苗可溶性糖和可溶性蛋白含量在不同光照条件下的差异主要是由光周期引起的(表3)。
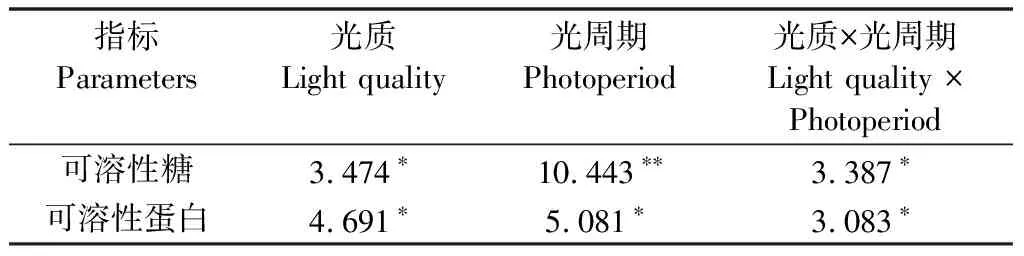
表3 不同光照条件下轮叶黑藻组培苗可溶性糖和蛋白含量的方差分析结果
红光照射下可溶性糖的含量稍高于其他光照,其他光质之间没有差异,光周期在不同光质下影响也不一致,白光照射下16 h/d光周期可溶性糖含量远高于其他两组,其他光照下光周期之间可溶性糖含量虽没有太大差异,但8 h/d光周期均为最低;而红光处理下可溶性蛋白含量最低,白光处理下最高,光质的影响同样不同,除白光下8 h/d光周期显著低于其他两组,剩余光照处理下不同光周期间没有显著差异(图5)。

(a)可溶性糖含量多重比较;(b)可溶性蛋白含量多重比较。不同字母表示存在显著差异,显著水平(P<0.05)。
2.1.4 光质和光周期对轮叶黑藻组培苗抗氧化酶体系的影响
方差分析结果显示:轮叶黑藻组培苗的整个抗氧化酶体系均受光质和光周期及其交互作用的显著影响。轮叶黑藻组培苗抗氧化酶体系在不同光照条件下的差异主要是由光周期引起的(表4)。

表4 不同光照条件下轮叶黑藻组培苗抗氧化酶体系的方差分析结果
光周期对POD活性的影响在不同光质下表现不同,8 h/d光周期下POD活性均最低,蓝光中16 h/d高于12 h/d,而红蓝混合光中则是12 h/d高于16 h/d;对SOD活性来说,白光下不同光周期之间没有显著性差异,红光和红蓝混合光下,16 h/d光周期下SOD活性最高,蓝光下则是8 h/d最高;而对CAT活性来说,白光下不同光周期之间同样没有显著性差异,其他3种光质下8 h/d光周期下基本最低,12 h/d光周期下最高;而MDA含量除红蓝混合光外,其他3种光质均为12 h/d>16 h/d>8 h/d,而光质之间的比较则是白光>红光>蓝光>红蓝混合光(图6)。

(a)POD活性多重比较;(b)SOD活性多重比较;(c)CAT活性多重比较;(d)MDA含量多重比较。不同字母表示存在显著差异,显著水平(P<0.05)。
2.2 底质及遮光程度对轮叶黑藻组培苗生长的影响
2.2.1 底质及遮光程度对轮叶黑藻形态指标和生物量的影响
方差分析结果显示:株高受底质类型和遮光程度及其交互作用的影响,而根数、芽数及各部分生物量则由底质类型和遮光程度分别影响。轮叶黑藻形态指标和各部分生物量在不同底质及遮光程度下的差异主要是由不同底质类型引起的(表5)。

表5 不同底质及遮光程度下轮叶黑藻形态指标和各部分生物量的方差分析结果
在种植前期,80%遮光下株高显著增长,而中后期除沙子底质外,不同遮光程度下株高无显著差异,沙子底质中不遮光处理株高最小;对不同底质来说,泥土底质和泥沙混合底质下的株高无差异,沙子底质株高最小。不遮光处理下轮叶黑藻生根最多,40%遮光和80%遮光下无显著差异,而泥土底质和泥沙混合底质中的轮叶黑藻生根数远多于沙子底质。芽数基本呈现不遮光>40%遮光>80%遮光,但这种差异在种植早期和后期不明显,对底质来说,前中期泥土和泥沙混合底质差异不明显,均远多于沙子底质,种植后期,泥土底质超过了泥沙混合底质(图7)。

(a)株高多重比较;(b)根数多重比较;(c)芽数多重比较。不同字母表示存在显著差异,显著水平(P<0.05)。
根部的干重基本呈现不遮光>40%遮光>80%遮光,但这种差异在种植前期和后期不明显,泥土和泥沙混合底质均远多于沙子底质。在种植的前期和后期,遮光程度对茎部干重的影响不显著,在中期则基本呈现不遮光>40%遮光>80%遮光的趋势;同时,在种植的中后期,泥土底质下的轮叶黑藻茎部干重最大,沙子底质下最小。除了泥沙混合底质外,在种植的前期和后期,遮光程度对叶片部分的干重影响不显著,而在中期,除了泥沙混合底质外,基本呈现80%遮光>不遮光>40%遮光,而底质的影响则为泥土>泥沙混合>沙子(图8)。

(a)根部干重多重比较;(b)茎部干重多重比较;(c)叶片干重多重比较。不同字母表示存在显著差异,显著水平(P<0.05)。
2.2.2 底质及遮光程度对轮叶黑藻生物量分配的影响
方差分析结果显示:根部的生物量占比与个体生长差异无关,只受底质类型和遮光程度影响,茎部的生物量占比只受遮光程度的影响,而个体间的生长差异影响了叶片生物量的占比,同时底质类型和遮光程度也影响叶片生物量的占比(表6)。
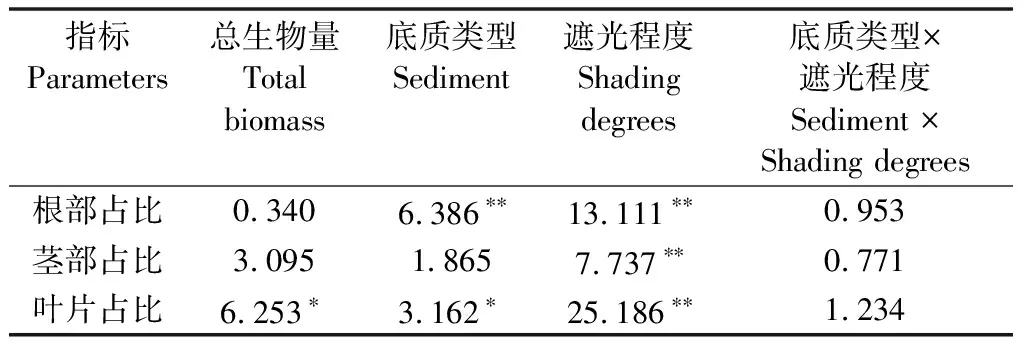
表6 不同底质及遮光程度对轮叶黑藻各部分生物量分配的影响
相同底质下,不同遮光程度下轮叶黑藻根部生物量占比差异不大,尤其是种植后期基本持平;在种植前期,泥沙混合底质下根部生物量占比略高于其他两组,在种植后期,沙子底质中的根生物量反超领先。种植前期和后期不同遮光处理下叶片生物量占比差异不显著,中期时80%遮光下叶片生物量占比最大,而对底质来说,不同底质下叶片生物量占比差异也不显著(图9)。

(a)根部占比多重比较;(b)叶片占比多重比较。不同字母表示存在显著差异,显著水平(P<0.05)。
40%遮光处理下茎部生物量占比最高,前中期不同遮光间存在显著差异,至种植后期差异缩小至不显著,而底质类型对根部生物量占比没有影响(图10)。

不同字母表示存在显著差异;显著水平(P<0.05)。
3 讨论
3.1 光质及光周期对轮叶黑藻组织培养的影响
光质、光周期对植物的形态建成、生理生化特性、产量和品质等众多方面均有调控作用[9]。在本实验中,红光显著提升组培苗干重,这与杜建芳等[10]对油菜的研究结果一致。研究表明蓝光会抑制植物的生长,而本实验中蓝光下组培苗干重和茎长与其他光色不存在显著差异,其原因可能为接种片段偏小,组培苗生长缓慢,初始片段造成的长度相对误差较大,抵消了蓝光的抑制作用。光周期对组培苗的干重和茎长有显著影响,随着光周期的延长,干重和茎长均先增大再减小,这与刘庆[11]对草莓的研究结果相同。
在一定范围内,光照可以促进原质体或黄化体向叶绿体的转化[12]。本实验中,12 h/d光周期大幅促进组培苗叶绿素合成,张欢等[13]在对油葵芽苗菜的研究中也有类似的结论,在生长前期(第15天~第35天)蓝光促进了叶绿素a的合成,抑制了叶绿素b的合成,故蓝光下叶绿素a/b最大,红蓝混合光次之,陈祥伟等[14]对乌塌菜的研究也得到了相同的结论。
光质对植物碳氮代谢有着重要作用,可溶性蛋白作为一种调控物质,在植物生理代谢过程中发挥着重要作用,其含量是衡量植物生长水平的重要指标[15];而可溶性糖则是一种营养物质,为植物代谢提供碳和能力,并促进细胞壁的形成[16],在一定程度上能反映植物的生长。本实验中红光显著提升了组培苗可溶性糖的积累,同时降低了可溶性蛋白的含量,白光照射的可溶性蛋白含量最高;一般来说,光周期延长会增加植物的物质积累[17],虽然本实验中8 h/d下物质积累量最低,但不同光周期之间没有显著差异,原因可能是16 h/d下叶绿素含量太低,光合作用产物整体减少,而植物的氮代谢又和碳代谢相互依存,所以整体可溶性糖和可溶性蛋白在不同光周期间并没有明显差异和趋势[18]。
光质和光周期对抗氧化酶体系的影响,不同植物结果不同[19]。抗氧化酶主要参与活性氧的清除,对植物生长发育有重要的影响。SOD能催化植物体内超氧自由基转化为过氧化氢,随后CAT和POD会立即将其分解为水,而MDA则是抗氧化酶体系代谢后的产物,抗氧化酶活性水平能反映植物抵抗环境胁迫的能力。本实验中,光周期的影响因光质的不同而改变,8 h/d下POD活性最低;而红光和红蓝混合光下,16 h/d光周期提高了SOD的活性,蓝光下8 h/d最高;而CAT活性是12 h/d>16 h/d>8 h/d,因此,光周期延长提高轮叶黑藻组培苗抗氧化酶活性。白光下MDA的含量最高,氧化程度较大,主要是因为组内12 h/d的光周期带来了巨大差异。除红蓝混合光外,剩余3种光质下MDA含量呈现为12 h/d>16 h/d>8 h/d。
3.2 底质及遮光程度对轮叶黑藻组培苗生长的影响
底质对植物不仅有固定作用,还能提供营养物质,不同的底质类型对植物生长产生不同影响[20]。而遮光所带来的光照差别也是重要影响因素。
本实验中,沙子底质中轮叶黑藻株高最低,芽数最少,生根数最多,这是由于沙子所含营养物质少,且结构松散,不易扎根,同时,沙子底质中虽然根数最多,但各组织干重最小,说明沙子底质无法为轮叶黑藻生长提供充足营养。同时沙子底质中根部生物量占比最高,说明在沙子底质中轮叶黑藻同比分配了更多的物质和能量给根部。泥土底质和泥沙混合底质之间各项指标差异较小,泥土底质中的株高、芽数、茎部干重和叶片干重稍高,而泥沙混合底质则有更高的根部干重和根部生物量占比,说明泥土底质更有助于轮叶黑藻的生长,泥沙底质中同样需要分配更多物质能量用于扎根。
植物的趋光性使得在80%遮光下轮叶黑藻提升株高并增加叶片生物量占比以补偿光照不足的影响来获取光照,分配更多的物质能量生长叶片进行光合作用。不遮光处理则提升了轮叶黑藻的根数、芽数、根部干重、茎部干重及根部生物量占比,说明轮叶黑藻在充足光照下优先进行根茎的生长。40%的遮光下则茎部生物量占比提高。
总之,光质和光周期对轮叶黑藻组培苗生长影响显著,红光显著提高了组培苗的生长速度和品质,而光周期的适当延长,也有利于轮叶黑藻组培苗生长,因此应考虑选用红色光照进行轮叶黑藻培养,光周期控制在12 h/d左右。同时,底质类型和遮光程度对轮叶黑藻种植影响显著,泥土底质显著提升了轮叶黑藻生长水平,而适度的遮光,也可以增加其生物量及各项形态指标,在实际应用中,在沙子底质种植轮叶黑藻则应选择较浅的水域种植,而在泥土或泥沙混合底质中可以选择较深的水域。
猜你喜欢
遗传(2023年9期)2023-09-25 09:31:44
农业科技通讯(2023年1期)2023-02-12 07:08:50
科学养鱼(2022年1期)2022-02-17 02:12:14
生物学通报(2021年7期)2021-07-01 03:24:52
生态科学(2020年3期)2020-06-08 12:09:54
当代水产(2019年10期)2019-12-18 05:28:56
农机化研究(2019年9期)2019-05-24 09:53:42
少儿科学周刊·少年版(2017年7期)2017-09-29 08:02:41
少儿科学周刊·儿童版(2017年7期)2017-09-29 02:22:58
中国酿造(2017年4期)2017-04-27 07:06:28
