
Music stimulus has a positive effect on survival and development of the larvae in swimming crab P ortunus trituberculatus*
2022-06-16 02:55ZhimingRENJiechiWANGChunlinWANGChangkaoMUYangfangYECeSHI
Zhiming REN , Jiechi WANG , Chunlin WANG ,**, Changkao MU , Yangfang YE ,Ce SHI 1, 2, **
1 Key Laboratory of Applied Marine Biotechnology, Ningbo University, Chinese Ministry of Education, Ningbo 315211, China
2 Collaborative Innovation Center for Zhejiang Marine High-efficiency and Healthy Aquaculture, Ningbo 315211, China
Abstract The sound stimulus has a significant impact on marine animals. However, many studies have focused on the negative impacts of sound on marine vertebrates. The positive impacts of sound on marine animals, especially on marine invertebrates, were rarely studied. In the present study, we evaluated the effects of music on the survival and development of an economically important marine invertebrate, swimming crab Portunus trituberculatus. Three pieces of classical music and a control group, i.e., Eine Kleine Nachtmusik by Mozart (EK), Bach-Violin Concerto No. 1 (BV), Anonymous Romanza by Anonymous (AR), and environmental sound (EN) were used. The sound pressure received were 163-dB root mean square (RMS)re 1 μPa for EK, 166-dB RMS re 1 μPa for BV, 165-dB RMS re 1 μPa for AR, 44-dB RMS re 1 μPa for EN, respectively. The larvae were subject to 3-h classical music transmission every day during the entire larval development period. The result showed that BV and AR significantly elevated the cumulative survival rate of the larva from 3.42%±1.77% (EN) to 10.65%±1.69% (BV) and 7.12%±1.37% (AR) respectively( P <0.05). Though music did not affect the cumulative development time of larvae from Zoea I to Zoea Ⅳ,it significantly increased the molting rate of megalopa from 24.00%±11.78% (EN) to 44.00%±13.06% (AR)and 46.67%±10.50% (BV) ( P <0.05). Musical exposure during critical growth intervals may contribute to the survival and development of larval crustaceans. From an application point of view, music intervention is much economically feasible. The observed music effects imply that music could further enhance growth,welfare, and production.
Keyword: swimming crab; Portunus trituberculatus; sound; classical music; oxygen consumption
1 INTRODUCTION
There is growing concern about the influence of sound from human activities on aquatic animals(Popper and Hastings, 2010). However, we are still far from understanding how sound affects aquatic animals either at the individual or population levels,for the knowledge of sound effects comes mainly from experiments with terrestrial animals. In addition to this, the data on sound stimuli effects other than noise has been rarely studied (Papoutsoglou et al.,2008b). Nevertheless, evidence showed that the other types of sound, music, for example, also affect aquatic animals (Papoutsoglou, 2012).
The soothing effects of music on human psychology and behavior are well known (Rykov et al., 2008).Although it is difficult to assess the perception of animals to music, researchers showed music could influence animals’ behavior, including fish (Rickard et al., 2005). Previous studies showed that Mozart music could improve the feeding rate of goldfish(Imanpoor et al., 2011), promote the growth performance of gilthead seabream (Sparusaurata)(Papoutsoglou et al., 2008b, 2015), carp (CyprinuscarpioL.) (Papoutsoglou et al., 2007, 2010), and rainbow trout (Oncorhynchusmykiss) (Papoutsoglou et al., 2013). Even though music has positively affected vertebrates like fish, much less is known about the effect of music on the early developmental stages of aquatic invertebrates (De Soto et al., 2013).
The swimming crabPortunustrituberculatusis an economically important crab species in China, of which the gross production reached 113 810 t in 2019(Fisheries Department of Agriculture, Ministry of China, 2020). Successful larval production is the premise of subsequent successful pond culture.However, suboptimal rearing condition, including nutrition (Shi et al., 2019c), background color (Shi et al., 2019a; Wang et al., 2019), light intensity (Dou et al., 2021) was reported to compromise the survival rate of the swimming crab larvae. Furthermore,evidence indicates that detrimental larvae-microbiota interactions can be a significant explanation for the poor performance and viability of the larvae (Vadstein et al., 2018), as little information is available regarding the interplay between microbiota and hostP.trituberculatus(Shi et al., 2019b). Consequently, only 4%-10% Zoea I larva could develop to juvenile crab.Interestingly, recent studies have found that sound can significantly influence the development of the crabs. The larval development of five species of crabs in New Zealand was accelerated when treated with habitat sounds (coral reef sounds) (Stanley et al.,2011), indicating that appropriate sounds may positively affect larval crustaceans.
Therefore, here we tested the effect of three pieces of music on the survival, development, and oxygen consumption rate ofP.trituberculatuslarvae. To the best of our knowledge, this is the first attempt to evaluate the effect of music on marine larval invertebrates.
2 MATERIAL AND METHOD
2.1 The source of the larvae and culture
The experiment was carried out in an aquacultural cooperative, Ningbo city, Zhejiang province, from April to June 2019. Once the larvae hatched, they were moved to indoor hatching tanks filled with seawater with temperature 22-25 °C, dissolved oxygen (DO) >6 mg/L, and salinity 22-24. The healthy, actively swimming Zoea I larvae were used in Experiment 1. Zoea I, Zoea Ⅱ, Zoea Ⅲ, Zoea Ⅳ,and Megalopa were used in Experiment 2.
2.2 Sound processing
In the present study, three pieces of classical music,i.e., Eine Kleine Nachtmusik by Mozart (EK), Bach-Violin Concerto No. 1 (BV), and Anonymous Romanza by Anonymous (AR) were used. Previous research showed that the three classical music positively influenced fishes’ growth performance and development (Papoutsoglou et al., 2007, 2015). Each piece of music (3 min) was re-recorded and calibrated with a hydrophone (HiTech HTI-96-MIN built-in amplifier) and recorder (wildlife SM4, sampling rate 96 kHz, USA) in an experimental tank without larvae.The waveform analysis is given in Fig.1. The highest energy frequency of different treatments was around 100 Hz, which is in line with underwater sound.Acoustic module analysis performed by MATLAB software was used to obtain the image of a frequency power spectral density (unit standard to 10 Hz,rectangular window, fast Fourier transform (FFT)length of 960 000, using the average of the previous 60 s) (Fig.1) (Voellmy et al., 2014). The Adobe Audition CC was used to analyze the waveform of the music. The processed music was stored in Windows Wave (WAV) format and played with a WAV player(ASUS FX63D7700 player, China, playing frequency 20-20 kHz), power amplifier (Langyun, LY-301K/BT, China, maximum power was 24 V), and an underwater speaker (PABGM, SX-450, China,diameter 220 mm, playing frequency 80-180 kHz,maximum power 90 W). The sound pressure received were 163-dB RMS re 1 μPa for EK, 166-dB RMS re 1 μPa for BV, 165-dB root mean square (RMS) re 1 μPa for AR, and 44-dB RMS re 1 μPa for environmental sound (EN).
2.3 Experiment 1: the effect of music on survival rate and development
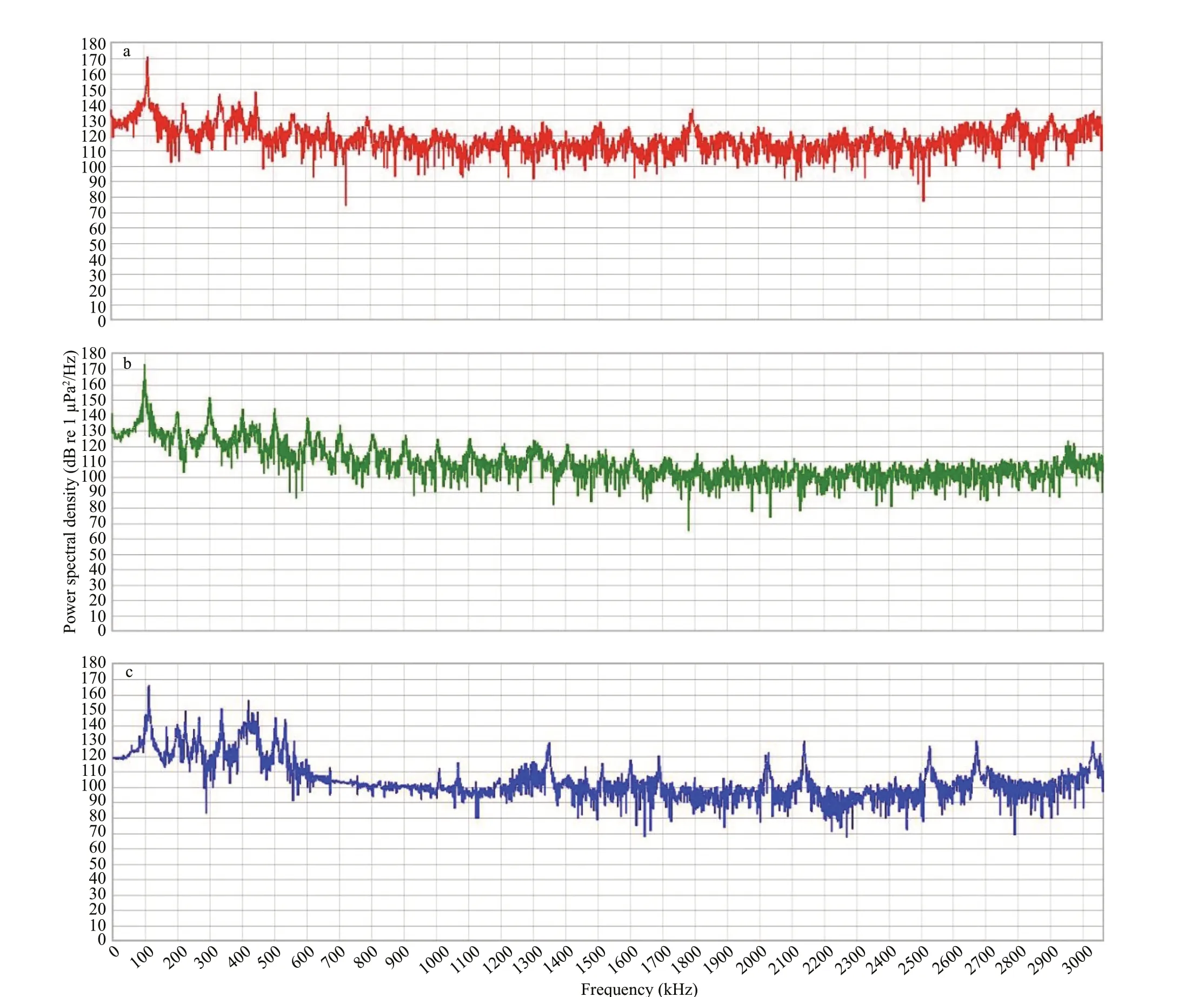
Fig.1 Variation in sound profiles for Eine Kleine Nachtmusik (a), Bach-Violin Concerto No.1 (b), and Anonymous Romanza (c)
Four treatments with quadruplicate in the present study, i. e., EK, BV, AR, and EN, were used. For each treatment, a rectangular Polyvinyl chloride (PVC)tank (50 cm×40 cm×40 cm) filled with seawater was used, and an underwater speaker was placed at the bottom of the tank. Different treatments were carried out in different compartments to avoid interference.At the beginning of the experiment, there were four cylindrical plastic crystal containers (diameter 10 cm,height 20 cm, volume 15 L) in one tank immersed into water with a mesh on the opening to ensure water exchange between the container and tank, and to prevent the larvae from escaping. There were 100 Zoea I larvae in each container as a replicate. The larvae were fed with newly hatchedArtemianauplii at 10∶00 and 17∶00 every day. In the meantime, the morality and molting were recorded. The motionless or white larvae were considered as dead, the molted larvae in each replication were transferred to a new identical container to decrease cannibalism.Temperature (22-25 °C), salinity (23-24), DO(>6.0 mg/L) was monitored daily by YSI ProDss multi-parameter water quality meter (Proplus, YSI,Yellow Springs, Ohio, USA). Ammonia nitrogen and nitrite (<0.5 mg/L) were monitored with HACH Portable multi-parameter water quality analyzer (US).The experiment lasted 20 d until all the larvae metamorphosed into crabs or died. Music transmission was carried out 3 h (8∶00-11∶00) every day during the whole experimental progress. An underwater speaker was placed at the bottom of the tank for the control group, but no sound was played.
2.4 Experiment 2: the effect of music on oxygen consumption rate
Before the experiment, the seawater used in the experiment was aerated to saturation. When the experiment started, the larvae of each stage were gently transferred to a container with 1-L aerated water in triplicates. The opening of the container was sealed with a plastic film to prevent the gaseous interchange. A control container without larvae was used to justify the aquatic respiration. For each treatment, four containers were water bathed in a rectangular tank with an underwater speaker placed at the bottom, and different treatments were carried out in different compartments to avoid interference, as in Experiment 1. The experiment lasted for 120 min.During the experiment, the music was played, and the DO at the beginning and the end of the experiment was measured with a YSI handheld Multiparameter water quality analyzer (Proplus, YSI, Yellow Springs,Ohio, USA). Groups of 100 Zoea I, 80 Zoea II, 60 Zoea III, 60 Zoea IV, and 40 Megalopa were used in the experiment, respectively. The larvae were dried at the end of the experiment to measure the net weight.The net weight of larvae was 0.085 mg (Zoea I),0.329 mg (Zoea Ⅱ), 0.5 mg (Zoea Ⅲ), 1.176 5 mg(Zoea Ⅳ), and 4.267 mg (Megalopa), respectively.
2.5 Date calculation and statistics analysis
The cumulative survival rate, staged survival rate,and oxygen consumption rate was calculated as follows:
cumulative survival rate (%)=(Of/Oi)×100,
staged survival rate (%)=(Of/Oix)×100,
oxygen consumption rate (mg/(g·h))=(A1-A2-A3)/(N×h×W),
Magalopa molting rate (%)=(Nm/Nt)×100,whereOiis the number of the larvae at the beginning of the experiment,Ofis the number of larvae at the end of each stage,Oixis the number of larvae at the beginning of each stage.A1is the DO (mg/L) at the beginning of the experiment,A2is the DO (mg/L) at the end of the experiment,A3is aquatic respiration,Nis the number of the larvae,his the experimental duration (h),Wis the weight of larvae of each stage(mg).Nmis the number of megalopae that molt into the juvenile crab, andNtis the total number of megalopa.
The cumulative survival rate, staged survival rate,oxygen consumption rate, and the development time were analyzed with SPSS for Windows, version 21.0.The normality and homogeneity of all data were tested by Levene’s test. One-way ANOVA and Tukey’s post hoc multiple comparisons were used to test significant differences among treatments. All the data are given as mean±SD.
3 RESULT
3.1 The effect of music on survival of larval P. trituberculatus
The staged survival rate is shown in Fig.2. The overall staged survival rate was more than 80% from Zoea I to Zoea Ⅲ, but for the Zoea Ⅳ and megalopa,it was less than 40%. There was no significant difference in survival rate among treatments until Zoea Ⅲ. For Zoea Ⅳ there were no significant differences in survival rate among musical treatments,but the survival rate of BV was significantly higher than EN (P<0.05). For the megalopa stage, the survival rate of BV and AR were significantly higher than EN (P<0.05).
The cumulative survival rate of different music treatments is given in Fig.3. There was no significant difference in the survival rate of larvae from Zoea I to Zoea Ⅲ. However, the survival rate of BV was significantly higher than EN in Zoea Ⅳ (P<0.05). For the megalopa, the survival rate BV was significantly higher than other treatments (P<0.05).
3.2 The effect of music on the development of larval P. trituberculatus
There was no significant difference in cumulative development time among treatments (P>0.05)(Table 1). The metamorphosis of megalopa is given in Fig.4. The megalopa molted mainly between 17-19 days post-hatch. A higher molting rate of megalopa was observed in musical treatment (Fig.4).
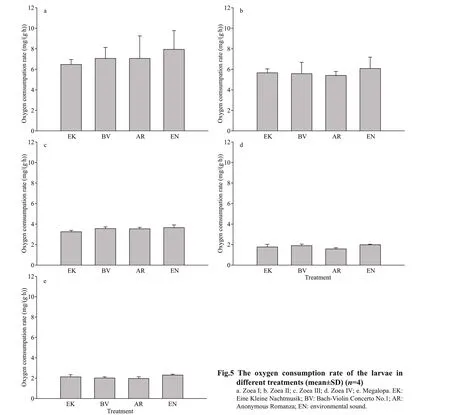
3.3 The effect of music on oxygen consumption rate
The oxygen consumption rate is given in Fig.5.With the development of larvae, the oxygen consumption rate decreased. There was no significant difference among treatments in oxygen consumption rate (P>0.05).
4 DISCUSSION
The objective of this study was to determine the effect of music on larval crustacean development. The result showed that the larvae treated with music BV and AR had a significantly higher survival rate than EN. Though the cumulative development time of the larvae was not affected, a significant increase in molting rate was found in the megalopa stage. Thus,the experiment indicates the potential effects of musical exposure during larval development. To the best of our knowledge, this is the first evidence that music could influence crustacean larvae.
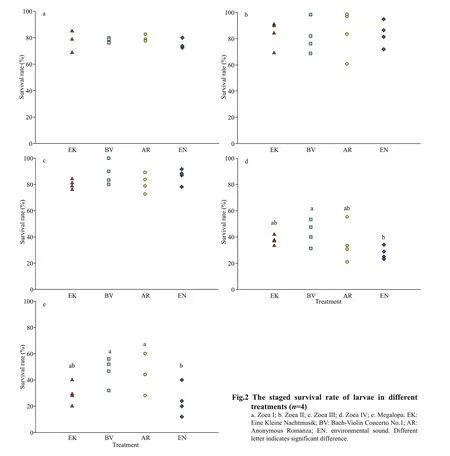
For crustaceans, the sensory structure of sound is statocyst (Lovell et al., 2005). Previous studies investigating the auditory capabilities of adult crustaceans have focused on electrophysiological methods. For example, the statocyst of adult prawn,Palaemonserratus, was shown to be sensitive to the motion of water particles induced by low-frequency sounds ranging from 100-3 000 Hz (Lovell et al.,2005). The hearing threshold of crab is not widely studied. Some research based on electrophysiological methods showed it could be as high as 140 dB(Hughes et al., 2014). Therefore, the sound pressure of the music was set atca. 160 dB in this study so that it could be more likely to reach the hearing threshold ofP.trituberculatus. However, because the crab larvae are tiny and lack strong tissue density gradients in early development, their acoustic receiving structure is not reported as far as we know. There is also the possibility that the observed effect of music is related to particle motion rather than to the pressure component. The pressure (P) and velocity components(v) are related asP=v×ztheoretically, wherezis the parameter that indicates the impedance of the medium(Urick, 1983). As a result, the larvae might detect particle motion by the body surface directly. A previous study showed that the larval development of the scallopP.novaezelandiae, which has no reported acoustic receiving structure, was significantly obstructed by the noise exposure (De Soto et al.,2013).
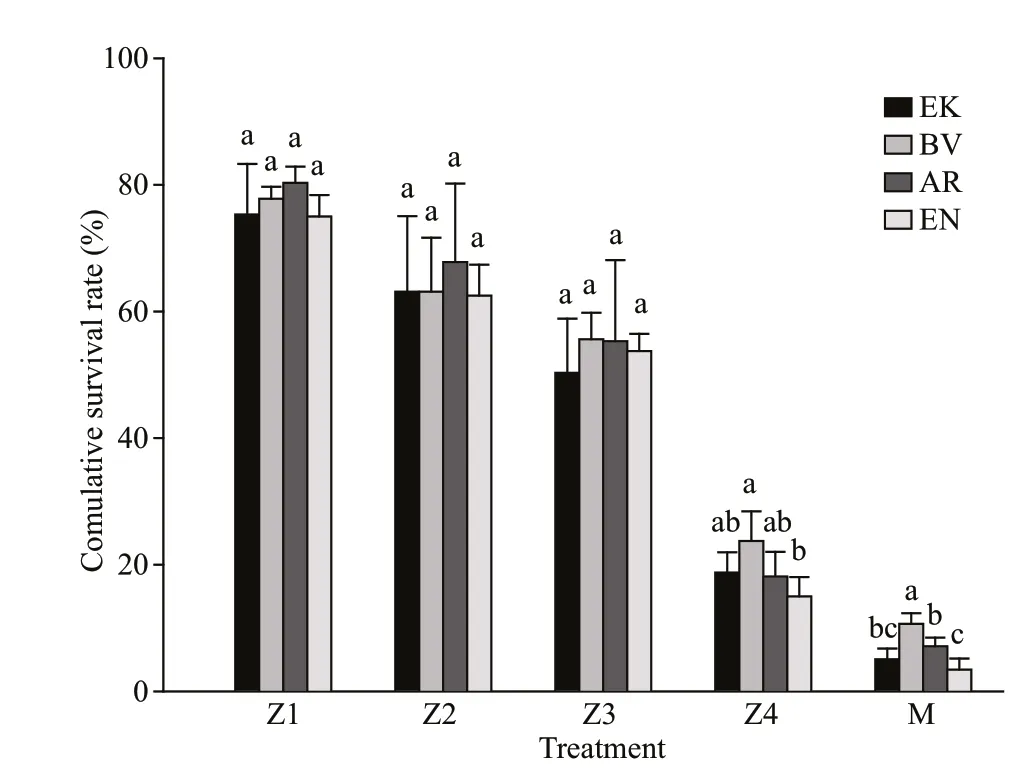
Fig.3 The cumulative survival rate of the larvae in different treatments (mean±SD) ( n=4)
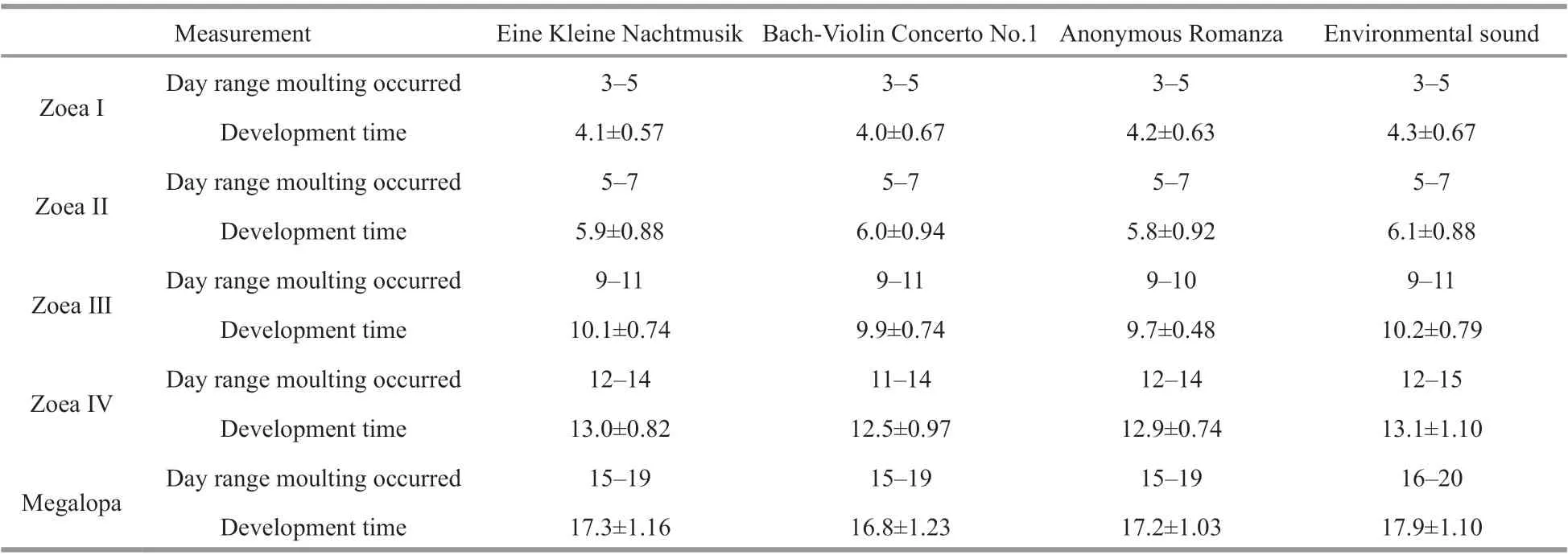
Table 1 Cumulative development time (days; mean±SD), and molting time range (days) of larvae in different treatments
In this experiment, all the music elevated the survival rate at the end of the metamorphosis states,and the larvae had a similar oxygen consumption rate among the four treatments. Nevertheless, shrimp exposed to noise had a higher oxygen consumption rate (Lagardère and Régnault, 1980), indicating higher energy loss. There is a direct conversion relationship between sound pressure and particle vibration (Hughes et al., 2014). As an unpredictable arrhythmic vibration, noise may elevate the stress response, increase predation risks (Wale et al., 2013;Sabet et al., 2016), and inhibit growth (Papoutsoglou et al., 2013, 2015). On the contrary, the function of communicating neurons and neurotransmitter levels could be activated by specific rhythms (Large and Crawford, 2002; Papoutsoglou et al., 2008a).Transmission of “classical” music into rearing tanks has been shown to appreciably enhance the fish homeostasis status and growth rate through a highly positive response in their brain auditory system(Papoutsoglou et al., 2007, 2008a, 2010). In this study, music may also help to reduce stress responses.It was observed that the music-treated larvae lay around the wall of the container, while the majority of EN larvae swam at the bottom of the tank with a higher risk of cannibalism. Fish populations subjected to music for 2 h daily exhibited the lowest levels of dopamine metabolite 3,4 dihydroxyphenylacetic acid(DOPAC), while homovanillic acid (HVA), and serotonin metabolite 5-hydroxyindoleacetic acid (5-HIAA) also showed a tendency towards lower values(Papoutsoglou et al., 2010). These neurotransmitters were proved to be closely related to aggressive behavior (De Almeida et al., 2005). Therefore, the classical music may act as a specific vibration resulted in more homogeneous populations and changed the social interactions (Papoutsoglou et al., 2007, 2010,2008b, 2013) by decreasing stress-induced brain serotonergic and dopaminergic activity (Papoutsoglou et al., 2008b), and ameliorating mass variation(Jobling, 1995; Papoutsoglou and Lyndon, 2006;Karakatsouli et al., 2007).
The other possibility is that some specific component of the music may act as a settlement cue.The late-stage larval crab could detect and respond to specific sound cues. For example, a previous study showed that megalopae of the rocky-reef crab(Leptograpsusvariegatus) exhibited a significant reduction in time to metamorphosis when exposed to underwater reef sound (Stanley et al., 2011),suggesting the sound may act as an acoustic settlement cue. The swimming crabP.trituberculatusis widely distributed on the west pacific coast. They often lay eggs in shallow seas with a depth of 3-5 m, especially near harbors or estuaries. The larvae were observed to settle in the open sandy beach. In this study, the musical treatments had a peak in the spectra around 100-500 Hz and higher frequency pulses from 500-3 000 Hz. Thus, the acoustic component may have a similar characteristic with the sound cue of the specific habitat of larvalP.trituberculatus. The speculation was also supported by the fact that the planktonic larvae from Zoea I to Zoea Ⅳ did not respond to music, but megalopa treated with music showed a significantly higher molting rate.
5 CONCLUSION
In conclusion, the present results indicate that sound transmitted positively affects larval survival and development, especially for the late-stage larvae.The music may have a rhythmic vibration that showed a soothing effect and/or acted as a settlement cue. The sound exposure during critical growth intervals may contribute to the development of larval crustaceans.Environmental auditory enrichment with classical music could be a possible way to elevate the performance of larval development for swimming crab. However, the threshold of music and the underlying physiological mechanism on crustacean larva still needs further exploration.
6 DATA AVAILABILITY STATEMENT
The data that support the findings of this study are available from the corresponding author upon reasonable request.
 Journal of Oceanology and Limnology2022年3期
Journal of Oceanology and Limnology2022年3期
- Journal of Oceanology and Limnology的其它文章
- Typhoon-induced wind waves in the northern East China Sea during two typhoon events: the impact of wind field and wave-current interaction*
- Effect of subsea dispersant application on deepwater oil spill in the South China Sea*
- Geochemical characteristics of cold-seep carbonates in Shenhu area, South China Sea*
- Examination of seasonal variation of the equatorial undercurrent termination in the Eastern Pacific diagnosed by ECCO2*
- Deviation of the Lagrangian particle tracing method in the evaluation of the Southern Hemisphere annual subduction rate*
- Immunostimulatory effect of quaternary degree and acetyl group of quaternized chitosan on macrophages RAW 264.7*