
Morphological analysis of Rhynchospio aff. asiatica(Annelida: Spionidae) and comments on the phylogeny and reproduction of the family Spionidae*
2022-06-16 02:55ZhiWANGTingXUJianWenQIUYingluJIZishanYUCaihuanKE
Zhi WANG , Ting XU , Jian-Wen QIU , Yinglu JI , Zishan YU , Caihuan KE
1 State Key Laboratory of Marine Environmental Science, College of Ocean and Earth Sciences, Xiamen University, Xiamen 361102, China
2 Department of Ocean Science and Hong Kong Branch of the Southern Marine Science and Engineering Guangdong Laboratory(Guangzhou), Hong Kong University of Science and Technology, Hong Kong 999077, China
3 Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou), Guangzhou 511458, China
4 Department of Biology and Hong Kong Branch of the Southern Marine Science and Engineering Guangdong Laboratory(Guangzhou), Hong Kong Baptist University, Hong Kong 999077, China
5 North China Sea Marine Forecasting Center of State Oceanic Administration, Qingdao 266061, China
6 College of Marine Life Sciences, Ocean University of China, Qingdao 266003, China
Abstract The genus Rhynchospio has fronto-lateral horns on prostomium, paired branchiae from chaetiger 2 to near the posterior end, capillary notochaetae only, and more than two pairs of pygidial cirri. Rhynchospio species are common in coastal soft bottom communities; nevertheless, many recorded Rhynchospio specimens around the world are currently undescribed. Here we described a Rhynchospio species based on specimens collected from Qingdao, China. Comparison with the reported DNA sequences of four gene markers (16S rRNA, 18S rRNA, 28S rRNA, and Histone H3) and brief morphological description of specimens collected from Jinhae Bay, South Korea, previously reported as Rhynchospio aff. asiatica, indicated that they are conspecific.Morphologically, specimens of R. aff. asiatica from Qingdao are characterized by having neuropodial hooded hooks from chaetigers 14-17 (vs. 10-23 in R. asiatica) to near pygidial chaetigers, sperm from chaetiger 11 to 14 (vs. from chaetiger 11 to 21-22 in R. asiatica), oocytes from chaetigers 16-17 to 26-39 (vs. from 22-24 in R. asiatica), and 4-6 (vs. up to 6 in R. asiatica) pygidial cirri. Genetically, Rhynchospio aff. asiatica is most closely related to R. arenincola Hartman, 1936 from California, USA with the interspecific distances of 20.02% (16S rRNA), 4.50% (18S rRNA), 8.44% (28S rRNA), 2.74% (Histone H3), and 6.10% (concatenated sequences). Water flow across the dorsum created by ciliary beating of the branchiae and nototrochs, observed on live specimens, may help transport gametes from reproductive segments in anterior and middle parts to the posterior brooding segments. Phylogenetic trees based on concatenated sequences of four gene markers of 54 spioniform species in 25 genera revealed two clades, covering the two subfamilies Spioninae and Nerininae respectively. Two families (i.e., Poecilochaetidae and Trochochaetidae) in the order Spionida were clustered within Spionidae, supporting a morphology-based proposal that these families bearing a pair of prehensile,grooved palps should be grouped within a more broadly defined family Spionidae. Mapping morphological and reproductive characteristics to the phylogenetic trees indicated that the ancestor of spionids might lack branchiae, broadcast spawn thick-envelop oocytes and ect-aquasperm, and produce planktotrophic larvae.
Keyword: Annelida; phylogeny; taxonomy; Yellow Sea
1 INTRODUCTION
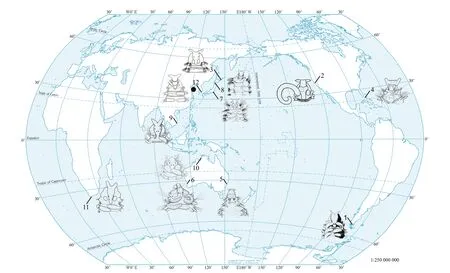
Fig.1 Type localities of Rhynchospio species (modified from Radashevsky et al. (2014))
Spionidae Grube, 1850 is a large family of annelids containing 39 valid genera and over 630 valid species(Read and Fauchald, 2021). There have been morphological (Mackie, 1996; Blake and Arnofsky,1999) and molecular (Abe et al., 2016) studies of the family, providing insights into the systematic relationships of the genera. However, the phylogenetic placement of some genera (e.g.,Aonides,Carazziella,Microspio,Dispio,Tripolydora, andPygospiopsis)has not been clarified, and in most genera the relationships among species have not been studied.This lack of phylogenetic studies is evident for spionids along the Chinese coasts: although there have been records of 17 genera and 69 species of spionids (Institute of Oceanology, Chinese Academy of Sciences and Liu, 2008; Zhou, 2008; Zhou and Li,2009; Zhou et al., 2010b), only 9 species in 4 genera(Scolelepis,Polydora,Boccardiella, andPseudopolydora) have been examined with molecular tools (Zhou et al., 2010a; Sato-Okoshi et al., 2013; Ye et al., 2015, 2017, 2019a, b).
RhynchospioHartman, 1936 is a genus of Spionidae characterized by a prostomium with fronto-lateral horns, paired branchiae from chaetiger 2 to near the posterior end, notochaetae all capillary and pygidium with more than two pairs of cirri (Foster, 1971;Radashevsky et al., 2014). A total of 13 species ofRhynchospiohas been described, but only 11 of species are considered valid (Fig.1);R.microcera(Dorsey,1977) andR.harrisae(Delgado-Blas and Díaz-Díaz,2010) might belong toMicrospio(Radashevsky et al.,2014). All adultRhynchospioare considered hermaphrodites, and currently 5 species, i.e.,R.glutaea(Ehlers, 1897),R.arenincola(Hartman, 1936),R.asiatica(Chlebovitsch, 1959),R.mzansi(Simon et al., 2019b), and an undescribed speciesR. aff.asiaticacollected from South Korea are classified as belonging to theRhynchospioglutaeacomplex characterized by having hooded hooks in neuropodia from chaetigers 9-18, tridentate both in juveniles and adults and pygidium with 2-5 pairs of cirri (Radashevsky, 2007a;Radashevsky et al., 2014). The other 7 species have quite distinct charateristics comparing to the species inRhynchospioglutaeacomplex (See Taxonomic key toRhynchospiospecies).
There have been several records ofRhynchospioalong the Chinese coast of the Yellow Sea, i.e.,“R.arenincola” and “R.glutaea” from Qingdao Bay(Wu et al., 1993; Ji, 2012), and “R.glutaea” from Shidao Bay (Zhou, 2008). TheseRhynchospiorecords are likely to be conspecific due to the short distance between Qingdao Bay and Shidao Bay (~210 km),but neither detailed morphological or DNA barcoding data were available to determine their taxonomic statuses. At about the same latitude across the Yellow Sea from Jinhae Bay (35°N) of South Korea, roughly 760 km from Qingdao, Radashevsky et al. (2014)found a species of theR.glutaeacomplex (temporally named “R. aff.asiatica”) based on 5.15% K2P genetic distance of the concatenated sequences of four genes(i.e., 16S rRNA, 18S rRNA, 28S rRNA, and Histone H3) fromR.arenincola. Further studies are needed to check whether theRhynchospiospecimens from Qingdao and Jinhae are conspecific or not.
Brooding activities on the body of female spionids is currently known only inStreblospioandRhynchospio, but whether the two genera use the same mechanism to transport their gametes to the dorsal brood region is unknown. InStreblospio, the gametes were suggested to be transported through coelom to the dorsal brood pouches formed by thinwalled, dorsolateral extensions of the coelom in middle segments (Collier and Jones, 1967). InRhynchospio, Radashevsky (2007a) speculated that the elongated capillaries forming a “hatchery” in posterior chaetigers could keep the elongated introsperm there, thus improving fertilization efficiency. However, it is unknown howRhynchospiotransport mature gametes from the anterior and middle fertile segments to the posterior “hatchery”.
Morphological and reproductive variations among spioniform genera had been used for uncovering their phylogenetic relationships (Sigvaldadóttir et al.,1997; Blake and Arnofsky, 1999). According to similar reproductive characteristics such as the occurrence of thick-enveloped oocyte, ect-aquasperm,and absence of egg capsule/mass in tube, twenty-one genera, including four spioniform genera (i.e.,Poecilochaetus,Trochochaeta,Heterospio, andUncispio) previously treated as independent families(i.e., Poecilochaetidae, Trochochaetidae,Longosomatidae, and Uncispionidae), had been grouped within the clade Nerininae in a cladistic analysis, which suggested that these four spioniform families should be considered as junior synonyms of Spionidae (Blake and Arnofsky, 1999). However,such analyses did not rule out the possibility of homoplasy in morphological and reproductive characteristics among genera of spionids, which may influence their clustering. A recent phylogenetic analysis conducted based on 16S rRNA and 18S rRNA gene sequences of spioniform genera, showed thatPoecilochaetusandTrochochaetawere nested within the clade of subfamily Nerininae, which confirmed previous suggestions to place them into Spionidae (Abe and Sato-Okoshi, 2021). However,phylogenetic analysis based on more gene sequences,together with morphological characteristics, are still needed to make better understanding of their phylogenetic placements within Spionidae.
Previous phylogenetic studies had provided useful information on the selection of ingroups and outgroups in constructing credible phylogeny among spionids using nucleotide data. Besides Abe and Sato-Okoshi(2021), Rousset et al. (2007) and Struck et al. (2007,2008) had also indicated close relationships between Spionidae and the two spioniform families Poecilochaetidae and Trochochaetidae, although these relationships were not well resolved. Also in these studies, several families (i.e., Sabellariidae,Sabellidae, and Serpulidae) of the order Sabellida,showed sister group relationships with Spionidae comparing to all other annelid families.
Another two families, i.e., Apistobranchidae and Longosomatidae, once believed to be a part of Spioniformia, however, had been found to be distantly related to Spionidae (Struck et al., 2008; Zrzavý et al.,2009; Blake and Maciolek, 2019; Blake and Petti,2019). For Longosomatidae, morphological similarities had been found between its single genusHeterospioand cirratulids (Blake and Maciolek,2019). Therefore, we followed these results to include Poecilochaetidae and Trochochaetidae as ingroups,and Sabellariidae, Sabellidae, and Serpulidae as outgroups in our phylogenetic analysis.
To explain the evolution of the morphological types of oocytes and sperm in spionids, two opposing hypotheses had been proposed. Söderström (1920)considered thin and smooth enveloped oocytes as a plesiomorphic characteristic and thick and sculptured envelope as a derived characteristic evolved in Spionidae. Hannerz (1956), on the contrary, suggested that thick-envelope oocytes might have evolved in a primary spionid and thin-envelope oocytes evolved in Spioninae due to reduction of the original thick envelope in connection with the evolution of brooding.Although the latter hypothesis had been supported by a comprehensive cladistic analysis of morphological and reproductive characteristics among genera of Spionidae (Radashevsky, 2007b), such analysis could not avoid the possible influences of homoplasy, and phylogenetic analysis using molecular data are needed to assess these hypotheses.
In this study, we presented the morphological features forRhynchospiospecimens collected from Qingdao Bay, China. Through observation of the ciliary beating in live specimens, we proposed a mechanism for transporting gametes from anterior and middle fertile segments to the brooding hatchery in posterior segments. By including DNA barcoding sequences for all available genera of spioniform annelids in our phylogenetic analyses, we aimed to determine the taxonomic statuses and phylogenetic placement of theRhynchospiospecimens from Qingdao Bay, and to study the evolution of reproduction of the family Spionidae. We also aimed to use nucleotide data, morphological and reproductive characteristics to make better estimates of the phylogenetic placements of two spioniform families(i.e., Poecilochaetidae and Trochochaetidae) recently included within Spionidae.
2 MATERIAL AND METHOD
2.1 Sample collection and preservation
Specimens ofRhynchospiowere collected from the high intertidal zone of Qingdao Bay (36°3′39.6″N,120°19′15.6″E), China, on November 2, 2014, June 18, 2015 and August 31, 2018. The sampling site was close to one of the two outfalls along the coast, located at the eastern side of Zhanqiao (the Pier, Supplementary Fig.S1). Sediments were washed through a 0.5-mm sieve and the retained specimens were fixed either with 10% formaldehyde in seawater and later transferred into 75% ethanol for morphological analysis or directly in 95% ethanol for DNA extraction.
2.2 Morphological analysis
Parapodia and chaetae of chaetigers 1, 2, 4, 10, 20,30, and a posterior one were dissected with iris scissors and mounted on slides. Photographs of parapodia and chaetae, taken using a Sony DSCWX350 digital camera mounted on an Olympus CX31RTSF microscope, were used for line-drawings.Scanning electron microscopy (SEM) was used for detailed observation of cilia on palps and branchiae.The anterior part of specimen #17 (Catalog No.:XMU-Pol-2021-050) was dehydrated in pure ethanol,dried with the critical point drying method, mounted on conductive carbon adhesives, sputter coated with gold and observed with a LEO 1530 FESEM scanning electron microscope.
2.3 Molecular-based phylogenetic analyses
2.3.1 DNA extraction, PCR amplification, and sequencing
Two specimens (XMU-Pol-2021-049, XMUPol-2021-050) preserved in 95% ethanol were used for DNA extraction. The posterior segments were dissected, and the genomic DNA was extracted using a DNeasy blood & tissue kit (QIAGEN, Hilden,Germany). Four primer pairs were used to amplify corresponding genes, i.e., 16SAR-L and 16SBR-H for the mitochondrial 16S rRNA gene (Palumbi et al.,1991); 1F and 9R for the nuclear 18S rRNA gene(Giribet et al., 1996); NLF184/21 and D3aR for the nuclear 28S rRNA gene (Lenaers et al., 1989; Van der Auwera et al., 1994) and H3af and H3ar for the Histone H3 gene (Colgan et al., 1998). The PCR protocol followed Zhang et al. (2018). PCR products were purified using a Zymoclean™ Gel DNA Recovery Kit and sequenced using Sanger sequencing at BGI Hong Kong.
2.3.2 Phylogenetic analyses
The sequences of these four genes of 24 spioniform genera, includingTrochochaetaandPoecilochaetus,as well as outgroups (i.e., Serpulidae, Sabellidae, and Sabellariidae) deposited in GenBank (https://www.ncbi.nlm.nih.gov/) were downloaded for phylogenetic analyses (Table 1). Taxa with no DNA sequences available, such as the generaAmphipolydora,Carazziella,Laubieriellus,Lindaspio,Pygospiopsis,Scolecolepides, andTripolydorain Spionidae and the family Uncispionidae, were not included in the analyses. Two families previously considered as members of Spioniformia, Apistobranchidae and Longosomatidae were excluded from the phylogenetic analyses due to their distant relationships from the spionid clade (Struck et al., 2008; Zrzavý et al., 2009;Blake and Maciolek, 2019; Blake and Petti, 2019).The four gene sequences were aligned using the MUSCLE algorithm implemented in the software Mesquite (Edgar, 2004), and poorly aligned positions were removed with the Gblocks Server (http://molevol.cmima.csic.es/castresana/Gblocks_server.html).The four trimmed genes were concatenated using
SequenceMatrix v.1.7.8 (Vaidya et al., 2011). The K2P genetic distances (Kimura, 1980) betweenRhynchospiospecies was estimated based on each gene sequence (i.e., 246-bp 16S rRNA, 1 493-bp 18S rRNA, 292-bp 28S rRNA, 289-bp Histone H3) and their concatenated sequence (2 320 bp) using MEGA X (Kumar et al., 2018). Two phylogenetic analyses were conducted using the Maximum Likelihood (ML)and Bayesian Inference (BI) approaches. One analysis focused on the phylogeny amongRhynchospiospecies, which were carried out based on the concatenated sequences (2 444 bp) of partial 16S rRNA (294 bp), 18S rRNA (1 502 bp), 28S rRNA(323 bp), Histone H3 (325 bp) genes ofRhynchospio,Boccardia, andPygospiorooted withMarenzelleriasequences. The other analysis focused on the phylogeny among spioniform genera, which were constructed based on the concatenated sequences(2 483 bp) of partial 16S rRNA (302 bp), 18S rRNA(1 535 bp), 28S rRNA (321 bp) and Histone H3(325 bp) genes of Spionidae, Sabellidae, and Sabellariidae rooted with Serpulidae sequences. The best fitting nucleotide-substitution model was evaluated using jModelTest v.2.1.1 (Darriba et al.,2012). Specifically, the ML analysis was conducted using the “thorough bootstrap” option with 1 000 bootstrap replicates via raxmlGUI v.2.0.0 (Edler et al., 2021). The BI analysis was conducted using MrBayes v.3.2.0 with corresponding models used in the ML analyses and Markov Chains were run for 10 000 000 generations with topologies being sampled every 1 000 generations (Ronquist and Huelsenbeck, 2003). The first 25% trees were discarded as “burn-in” and software Tracer v.1.7.1 was used to check for the convergence of the trees(Rambaut et al., 2018).
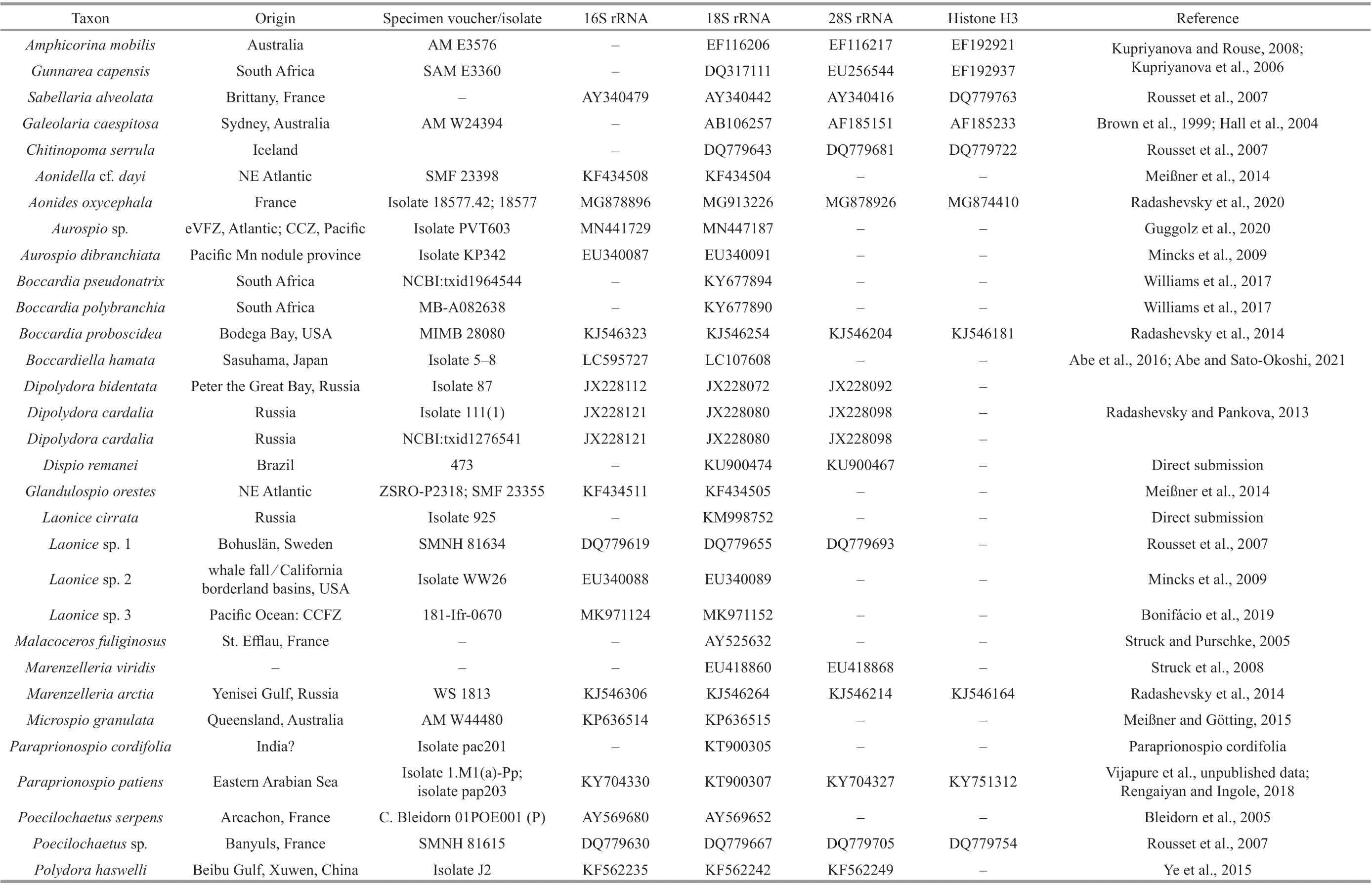
Table 1 GenBank accession numbers and specimen voucher/isolate information of used spionids in this study
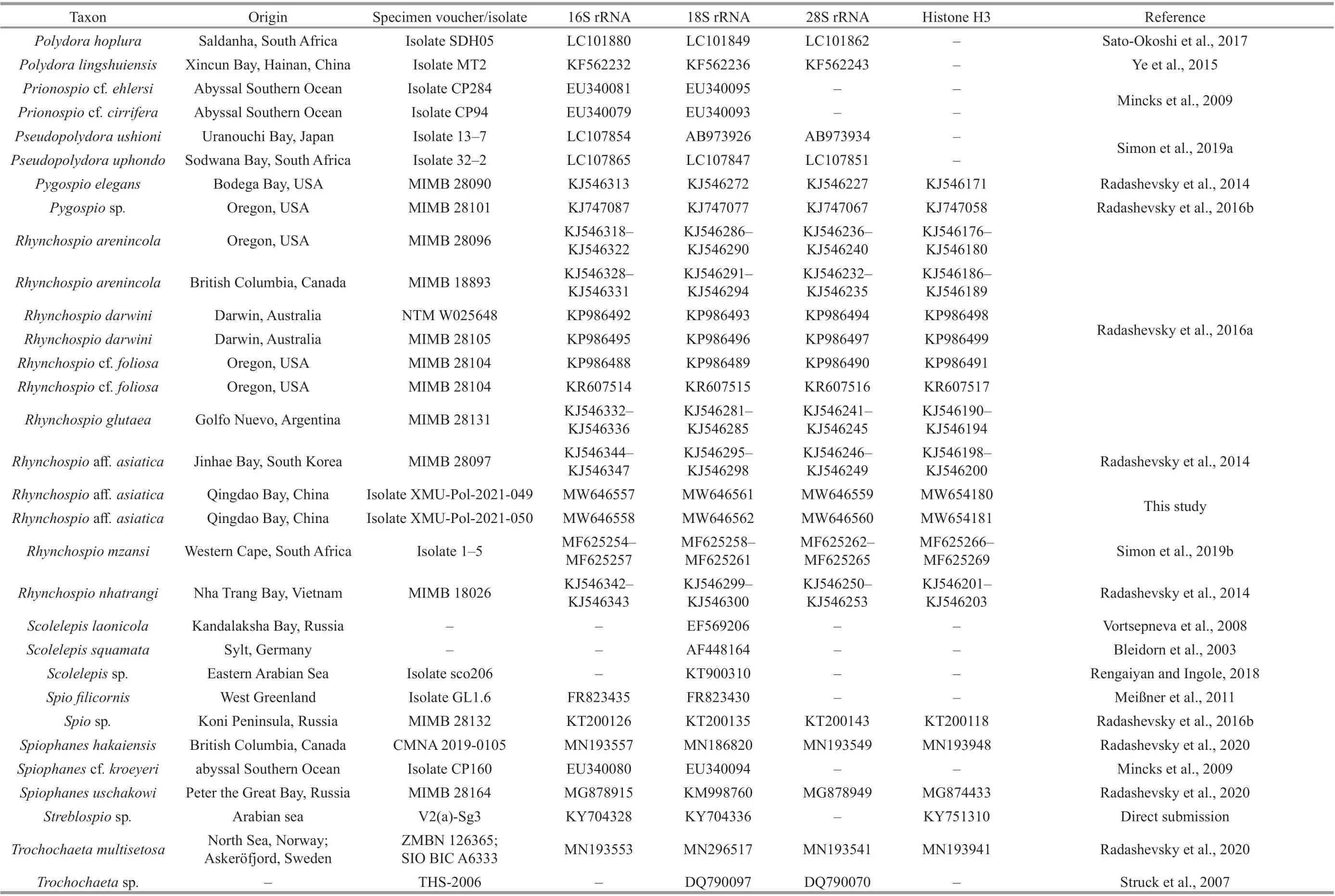
Table 1 Continued
2.4 Morphological and reproductive characteristics of spionids
To study the evolution of morphological and reproductive characteristics with reference to the molecular-based phylogenetic analyses, the following characteristics were compiled for all the 54 analyzed spioniform species. There are morphological groups:Group A: heavy spines present in chaetiger 5 and fullsized branchiae starting from chaetiger 7, with exceptions in four genera: branchiae inBoccardiaandBoccardiellastarting from chaetiger 7 in juveniles but chaetiger 2 in adults; branchiae inDipolydorastarting from chaetiger 7-10; though not included in the phylogenetic analysis, adultTripolydorahas fullsized branchiae from chaetiger 7, but short ones in chaetigers 2-6. Group B: without heavy spines and branchiae starting from chaetiger 1 or 2 to near the end. Group C: branchiae starting from chaetiger 1, 2,or 3, on a certain number of anterior segments. Group D: without branchiae in adults. Types of sperm (i.e.,Ect: ect-aquasperm; and Int: introsperm). Types of oocyte (i.e., type I: oocytes with thick envelopes bearing numerous vesicles; type II: oocytes with thick envelopes without vesicles; and type III: oocytes with thin envelopes without vesicles). Reproductive patterns (i.e., Bro: brooding; and Nbro: non-brooding).Egg protection modes (i.e., cap I: hollow cylinder attached to tube; cap II: beadlike string capsules attached to the wall of tube; cap III: series of single capsules; intub: egg cocoon/egg mass in tube; onfem:on the body of female; eem: external egg masses/external jellylike cocoon; and Brsp: broadcast spawning into water column). Types of larval development (i.e., Pla: planktotrophic; Lec:lecithotrophic; and Dir: direct development). The morphological groups were created by the authors in this study and the other groups mentioned here were present on different studies (Jamieson and Rouse,1989; Blake and Arnofsky, 1999; Radashevsky et al.,2016a, 2018; Blake et al., 2020).
3 RESULT
3.1 Systematics
Family Spionidae Grube, 1850
GenusRhynchospioHartman, 1936
Type species:RhynchospioarenincolaHartman,1936, by original designation. Type locality: San Mateo County, central California, USA.
Diagnosis (emended from Blake et al., 2020):Prostomium with fronto-lateral horns, caruncle variously developed; eyespots two pairs; occipital antenna absent. Branchiae from chaetiger 2 to near the end, free from dorsal lamellae or only fused basally. Notochaetae all capillary. Neurochaetae capillaries, hooded hooks, and sabre chaetae.Pygidium with cirri or lobes.
Remark: Blake et al. (2020) introduced the eyespots ofRhynchospioas present or absent, but we found that 10 of the 11Rhynchospiospecies have 4 eyespots, whileR.glutaeahas only 2 after the original description.
Rhynchospioaff.asiatica
(Figs.1-5, Supplementary Fig.S1, Tables 1-3,Supplementary Table S1; Supplementary Videos S1-S4)
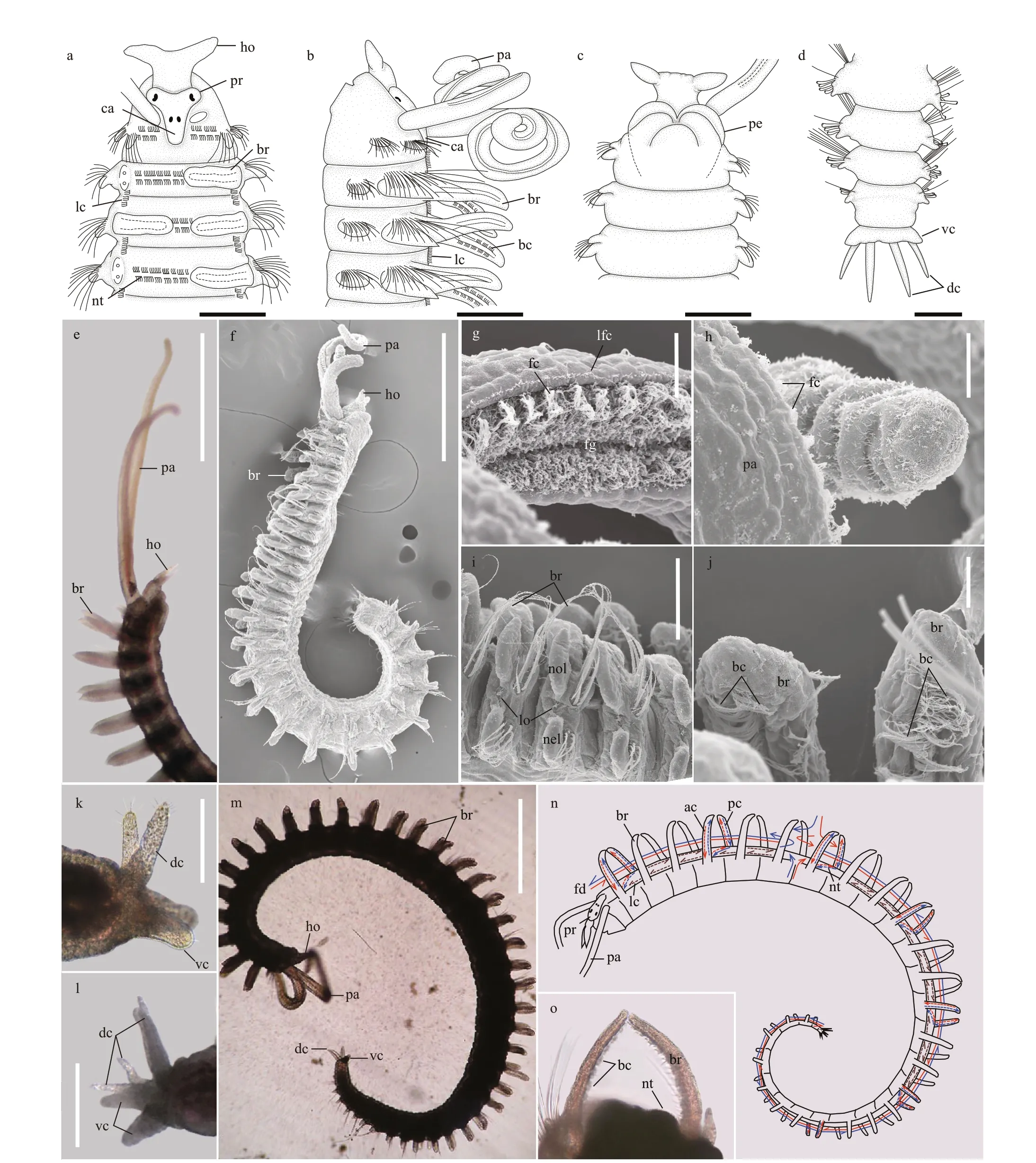
Fig.2 Morphology of Rhynchospio aff. asiatica
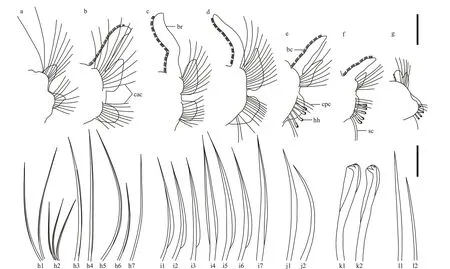
Fig.3 Parapodia and chaetae of Rhynchospio aff. asiatica
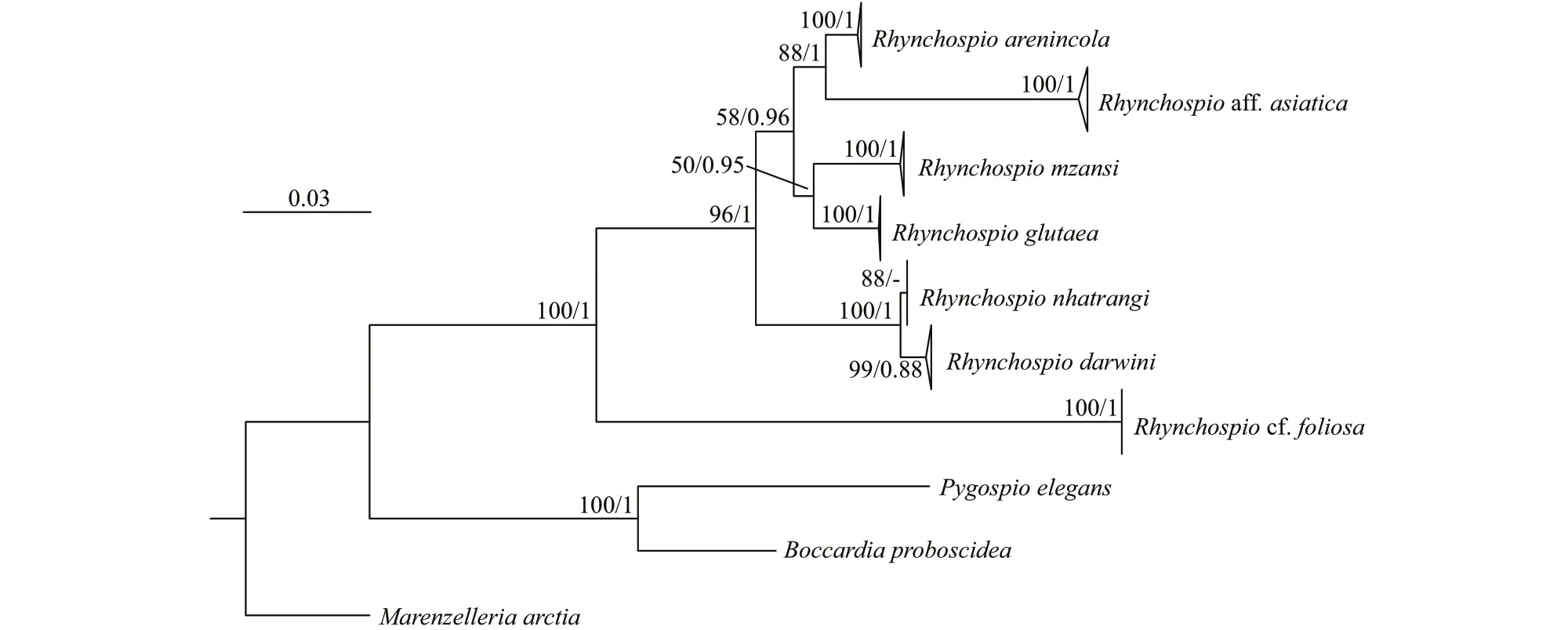
Fig.4 Phylogenetic tree reconstructed with Maximum Likelihood (ML) and Bayesian Inference (BI) methods
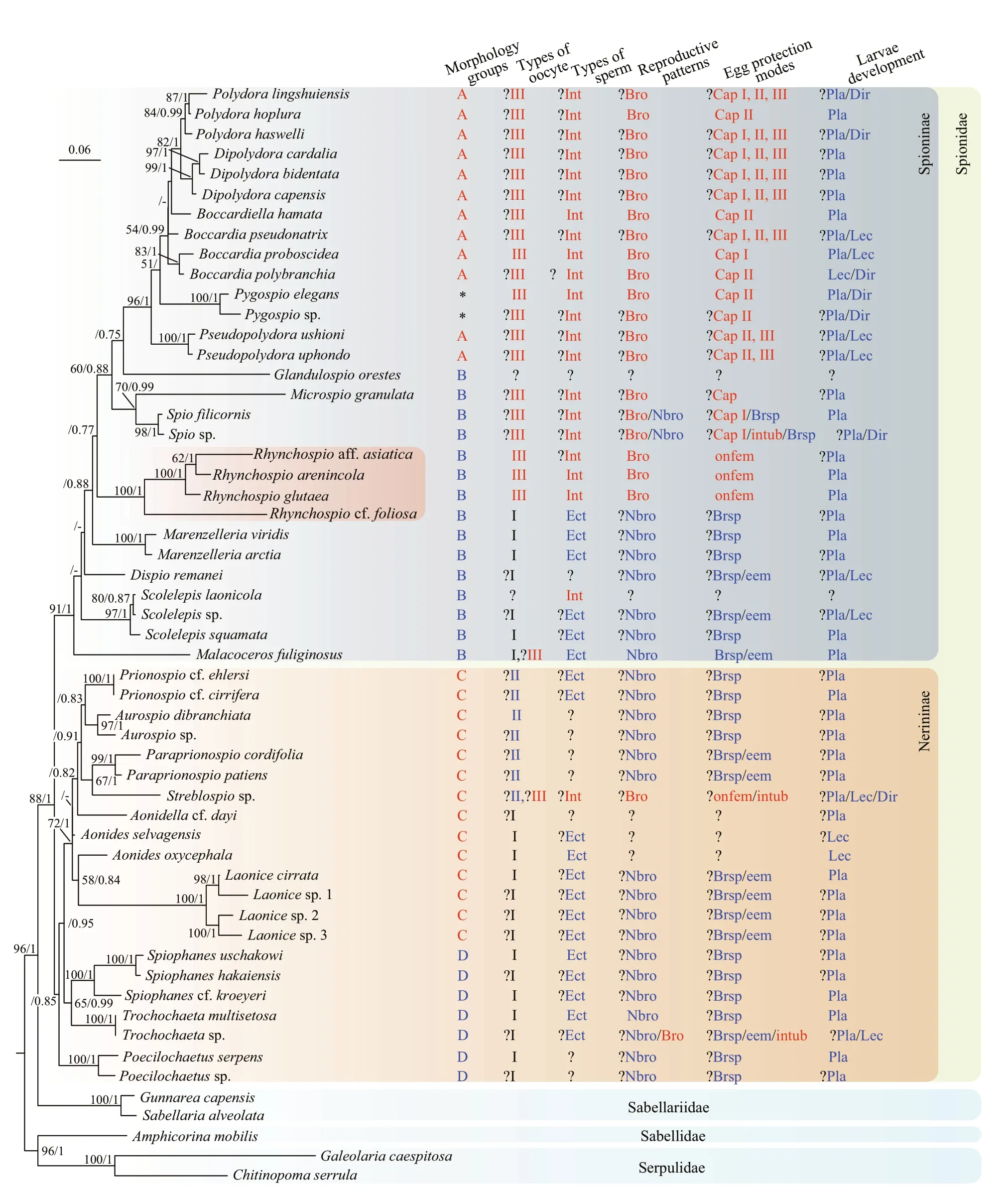
Fig.5 Phylogenetic tree reconstructed with Maximum Likelihood (ML) and Bayesian Inference (BI) methods
Materials examined: One specimen (XMUPol-2021-050) collected on 2 November 2014, three specimens (XMU-Pol-2021-034, XMU-Pol-2021-035,and XMU-Pol-2021-036) collected on 18 June 2015,and thirteen specimens (XMU-Pol-2021-037, XMUPol-2021-038, XMU-Pol-2021-039, XMU-Pol-2021-040, XMU-Pol-2021-041, XMU-Pol-2021-042, XMUPol-2021-043, XMU-Pol-2021-044, XMU-Pol-2021-045, XMU-Pol-2021-046, XMU-Pol-2021-047, XMUPol-2021-048, and XMU-Pol-2021-049) collected on 31 August 2018 from an intertidal muddy sand beach in Qingdao Bay, Shandong, China (Table 2).
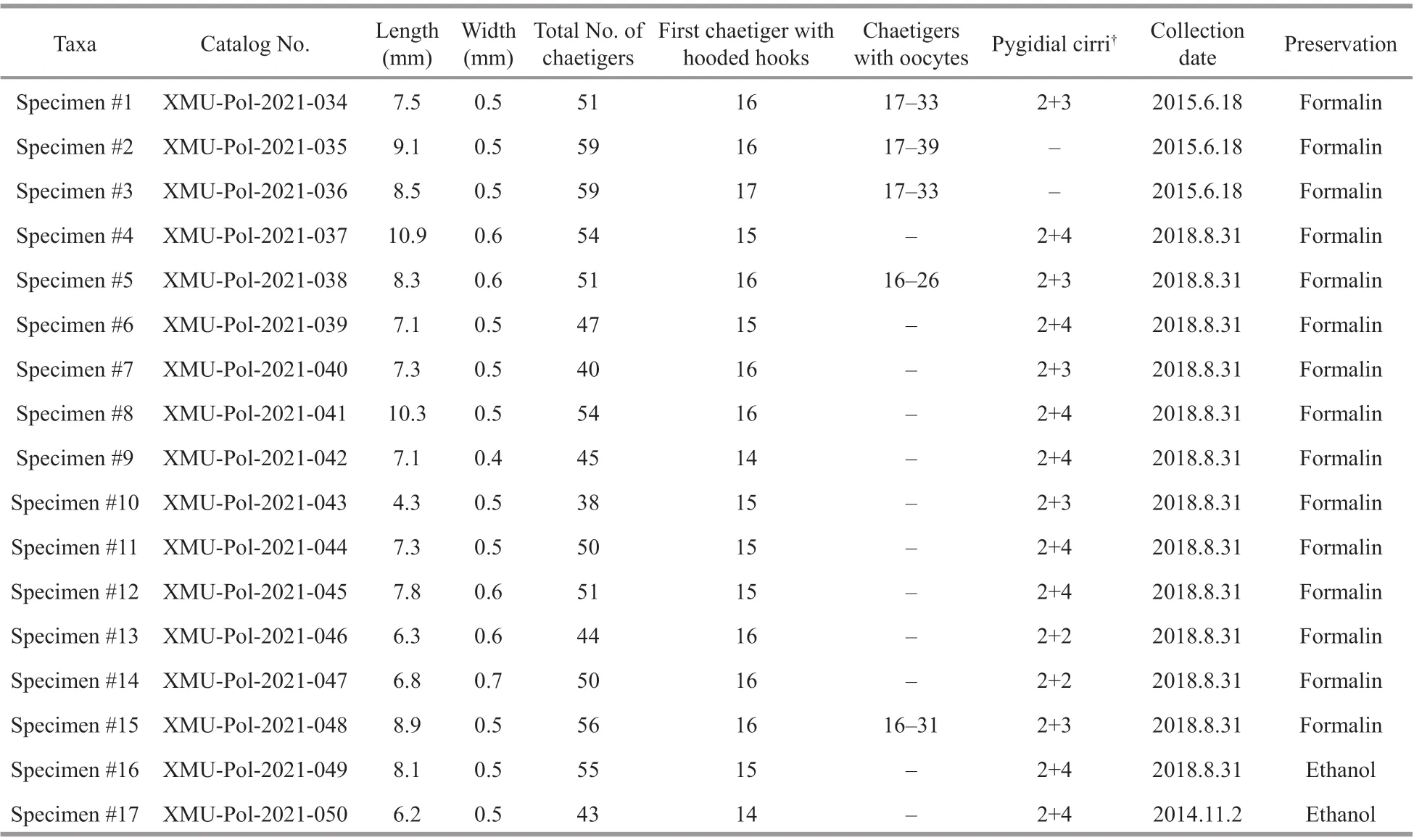
Table 2 Major morphological characteristics for 17 specimens of Rhynchospio aff. asiatica (XMU-Pol-2021-034 to XMUPol-2021-050) used in this study
Description: Body slender, anterior part slightly wider and taper offto the end (Fig.2a-d). Prostomium T-shaped, bearing two conical latero-frontal horns(Fig.2a); caruncle not elevated above prostomium,reaching chaetiger 1 (Fig.2a & b); nuchal organs not observed; eyespots two pairs arranged in inverted trapezoid, small, brownish in color, anterior eyes crescent, twice as large as posterior ones (Fig.2a).Occipital tentacle absent (Fig.2a). Palps extending backward to chaetiger 9, with ventral longitudinal food groove along whole length (Fig.2b, e-h).
Branchiae flattened, present from chaetiger 2 to near the posterior end (Fig.2e & m), free from notopodial postchaetal lamellae (Fig.3b-g); branchiae longer, wider in anterior and middle chaetigers,shorter, narrower in posterior chaetigers (Fig.2a, b, e,m). Paired branchiae bending towards each other in each segment, with branchial surfaces oriented parallel to longitudinal body axis (Fig.2a, i, j). Two parallel rows of branchial cilia present along inner surface of each branchia (Fig.2b, j, n), together with double-row nototrochs, forming a ciliary ring on dorsum of each segment (Fig.2a, b, o). Metameric nuchal organs not observed. Short bands of cilia present between segments on dorsolateral edges(Fig.2a & b). Notopodial and neuropodial postchaetal lamellae of chaetiger 1 triangular, small (Fig.3a);notopodial postchaetal lamellae larger from chaetiger 2, subtriangular with terminal tips, becoming smaller from middle to posterior chaetigers (Fig.3b-g).Neuropodial postchaetal lamellae oval shaped from chaetiger 2, becoming wider and shorter in middle chaetigers, and obtuse triangular in posterior chaetigers (Fig.3b-g).
Anterior chaetigers with two rows of capillary chaetae in both notopodia and neuropodia (Fig.2i).Chaetae in anterior row with wide unilateral or bilateral limbation and fine granulation on distal part of shaft (Fig.3i). Chaetae in posterior row longer and thinner, without granulation (Fig.3h). Hooded hooks present only in neuropodia, 3-7 in posterior row(Fig.3e-g), from chaetiger 14-17 to posterior end(Fig.3e-g), with one main fang and two smaller upper teeth situated one above the other (Fig.3k). Sabre chaetae without limbation, 2-3 per rami, present below hooded hooks from chaetigers 14-17 (Fig.3e-g,j). Companion chaetae without limbation, 2-4 in number, present in anterior row of hooded hooks(Fig.3e-g, l).
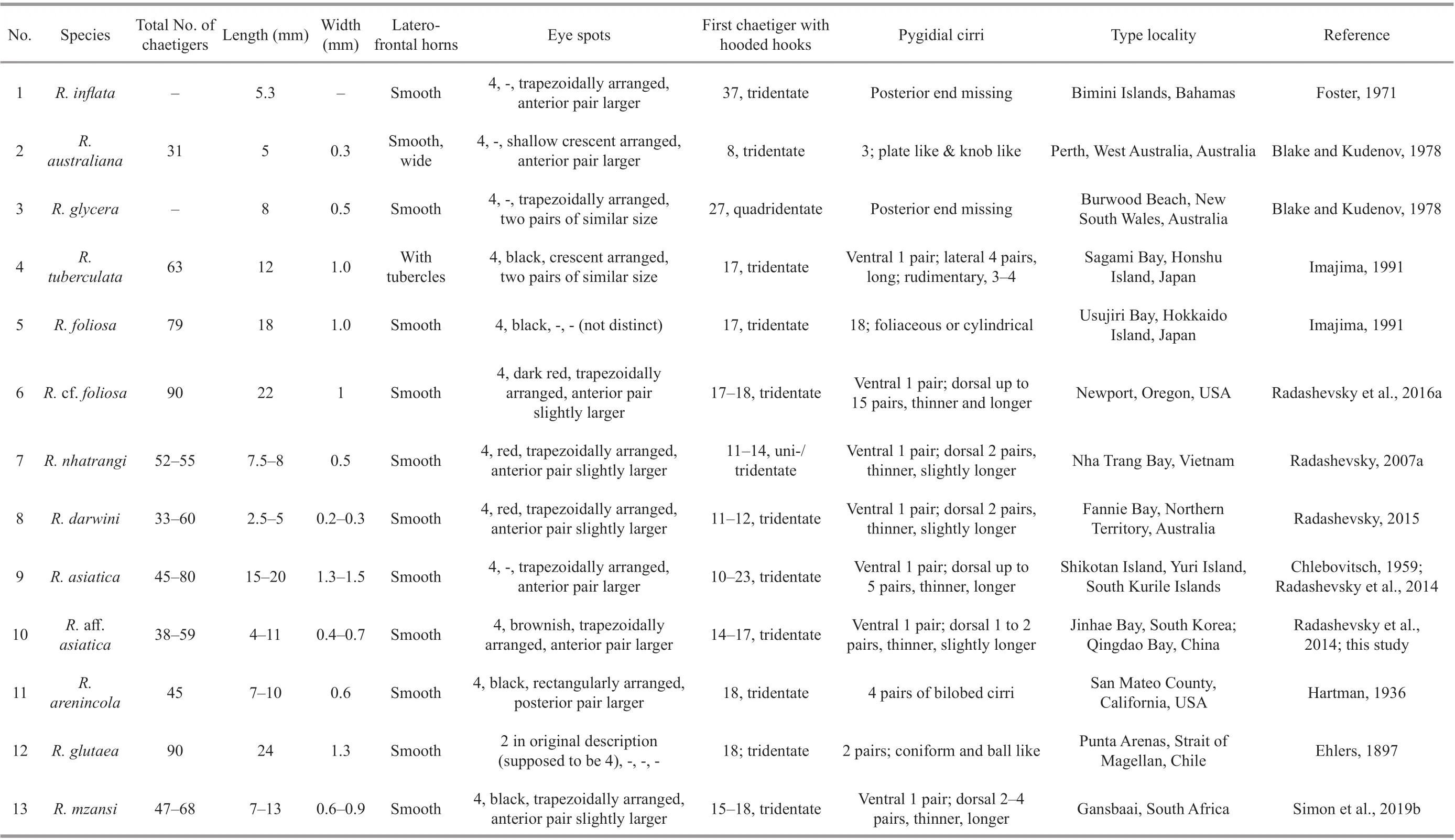
Table 3 Key morphological data of Rhynchospio species around the world
Pygidium with one pair of stumpy ventral cirri and 1 to 2 pairs of thinner and longer dorsal cirri in most specimens (Fig.2d, k, l).
Dorsal tunnel: Observations of live specimens ofRhynchospiocollected from Qingdao revealed a dorsal tunnel for the first time (Fig.2m-o). On most segments, a somewhat closed circle is formed by paired branchiae and the dorsum (Fig.2o;Supplementary Video S1). All these circles together form a dorsal tunnel. Two rows of parallel cilia are present along the inner side of branchiae and the dorsum, forming two parallel ciliary rings on each segment (Fig.2j & n). The anterior ciliary band beats backward and the posterior ciliary band beats forward,creating a dexioplectic metachronal wave with the effective beating of cilia being to the right of the wave(Supplementary Videos S2 & S3). The same pattern of cilia beating has been widely observed in annelids(Knight-Jones, 1954). When the anterior row of cilia in all segments beats in the same direction from anterior to posterior, it forms a backward water flow;when the posterior row of cilia in each segment beats from posterior to anterior direction, it forms a forward water flow (Supplementary Video S3). By coordinating the beating of the anterior and posterior ciliary bands,the worm can transport small particles along the dorsal tunnel from anterior to posterior, or vice versa(Supplementary Videos S3 & S4).
Ecology: The specimens ofRhynchospioaff.asiaticaexamined in this study live in muddy sand sediment in the high intertidal zone of Qingdao Bay, a habitat encountering fluctuations in environmental conditions created by inundation of tides and discharge of domestic sewage (Supplementary Fig.S1). Du et al. (2011) indicated higher sedimentary organic content (1.81% vs. 0.44%) in the studied intertidal zone than that in offshore zone. In addition,as a tourist spot, the study site encounters frequent human disturbances including trampling and clam digging especially in the summer. The abundance ofRhynchospioin the study site varied seasonally from high at 21 522 inds./m2in August to low at 308 inds./m2in May (Wang, 2015).
K2P genetic distances: Among theRhynchospiospecies compared, the Qingdao Bay sequences are most closely related to those ofR. aff.asiaticafrom Jinhae Bay, South Korea. The K2P genetic distances between the two populations was only 0.41% for 16S rRNA, 0.12% for 18S rRNA, 0 for 28S rRNA, 0.88%for Histone H3, and 0.19% for the concatenated sequences. These small genetic distances clearly indicated that the Qingdao population is conspecific to the South Korea population, which will be described as a new species (Vasily I. Radashevsky, personal communication). Among the describedRhynchospiospecies,R.arenincolahas the smallest average K2P distances withR. aff.asiatica, i.e., 6.10% for the concatenated sequences of four genes, 20.02% for 16S rRNA, 4.50% for 18S rRNA, 8.44% for 28S rRNA, and 2.74% for Histone H3. Other species are more distantly related (Supplementary Table S1).
3.2 Phylogeny and morphology in spionids
Phylogenetic analysis (Fig.4) shows that allRhynchospioaff.asiaticaspecimens collected from Qingdao and Jinhae form a well-supported clade(BS=100; BPP=1) that is sister toR.arenincolafrom California (BS=88; BPP=1); and all sevenRhynchospiospecies included in the analysis form a monophyletic clade (BS=100; BPP=1). All spionids form a single clade (BS=88; BPP=1) that is sister to Sabellariidae (BS=96; BPP=1). Two clades are detected within spionids: clade 1 (here referred to as Spioninae) includesPolydora,Dipolydora,Boccardiella,Boccardia,Pygospio,Pseudopolydora,Glandulospio,Microspio,Spio,Rhynchospio,Marenzelleria,Dispio,Scolelepis, andMalacoceros(BS=91; BPP=1); clade 2 (here referred to as Nerininae) includesPrionospio,Aurospio,Paraprionospio,Streblospio,Aonidella,Aonides,Laonice,Spiophanes,Trochochaeta, andPoecilochaetus, although with low support values(BS=46; BPP=0.85). Morphologically, all taxa within morphology group A, together withPygospio, form a single clade (BS=96; BPP=1), while all taxa within morphology group B showed paraphyletic relationships (excludingSpioandMicrospio). All taxa within morphology group C form a single clade(BS=72; BPP=1), whilePoecilochaetuswithin morphology group D showed paraphyletic relationship with the group formed bySpiophanesandTrochochaeta(BS=65; BPP=0.99).
3.3 Reproductive and larval characteristics in spionids
Despite there are ambiguous or missing information in reproductive and larval characteristics, distinct clusters corresponding to the phylogenetic tree could be noted within each characteristic (Fig.5). Four clusters of spionids according to types of oocytes include a cluster corresponding to type III, a cluster corresponding to type II, and two separate clusters corresponding to type I at the basal positions of Spioninae and Nerininae. Two exceptions inMalacocerosandStreblospioare noted with both thick- and thin-enveloped oocytes. Three characteristics (i.e., types of sperm, reproductive patterns and egg protection modes) are basically divided into two clusters, i.e., one cluster includes taxa corresponding to those with oocyte type III,while the other cluster including other spioniform taxa excluding some species ofTrochochaeta,Streblospio, and an ectoparasitic spionidScolelepislaonicola. Almost all spioniform taxa have planktotrophic larval development, while lecithotrophic and direct development modes are only found in about 1/4 to 1/3 of the analyzed genera in this study.
4 DISCUSSION
Rhynchospioaff.asiatica, originally recorded from Jinhae, South Korea, is characterized by having sperm in chaetigers 11-14, and oocytes starting from chaetiger 16 (Radashevsky et al., 2014). This species described based on individuals from Jinhae will be formally named in a manuscript submitted for publication (Vasily I. Radashevsky, personal communication). OurRhynchopsioindividuals from Qingdao are characterized by having smooth laterofrontal horns, four trapezoidally arranged eyespots,neuropodial hooded hooks starting from chaetigers 14-17 to posterior end, sperm in chaetigers 11-14,oocytes from chaetigers 16-17 to chaetigers 26-39,and 2-3 pairs of pygidial cirri. Morphologically, the specimens from Qingdao could be classified as belonging to theRhynchospioglutaeacomplex, and further identified asR. aff.asiaticadue to their similarity in the distribution of sperm and oocytes along segments (Radashevsky et al., 2014). Molecular results support the two populations ofRhynchospioto be conspecific by showing quite small inter-population K2P distances.
Although thirteenRhynchospiospecies including an unnamed speciesR. cf.foliosahave been recorded,molecular data are only available for seven species(Radashevsky et al., 2014, 2016a; Simon et al., 2019b;this study), and the phylogenetic placements of the other six species (i.e.,R.asiatica,R.foliosa,R.tuberculata,R.australiana,R.glycera, andR.inflata) are still undetermined. Based on morphological characteristics, threeRhynchospiospecies, i.e.,R.australiana,R.glycera, andR.inflata,have apparent divergences withR.foliosa/R. cf.foliosa, the two tropical species (i.e.,R.darwiniandR.nhatrangi), and those in theRhynchospioglutaeacomplex, Specifically,R.australianabears a pair of broad horns (rather than digitiform in theR.glutaeacomplex), lacking (rather than possessing) notopodial capillaries on chaetiger 1, plate-like (rather than cirriform) pygidium, and perpendicularly (rather than parallelly) oriented branchial surface, inner and outer ciliary bands (rather than two parallel inner bands),and lacking (rather than possessing) transverse ciliary dorsal ridges on each segment (Blake and Kudenov,1978). The branchial orientation and ciliary pattern ofR.australianaare more similar to those ofMalacoceros(Hourdez et al., 2006) andAonides(Radashevsky, 2015).R.glycerahas two posterior pairs of digitiform nuchal lobes (rather than lacking nuchal lobes in theR.glutaeacomplex) on caruncle,and bears quadridentate (rather than tridentate)hooded hooks (Blake and Kudenov, 1978).R.inflatawas named after its inflated anterior body region and its neuropodial hooded hooks starting from chaetiger 37, far posterior than those in the otherRhynchpsiospecies (Table 3). Not described but showed in figures(Figs. 113, 114 in Foster, 1971),R.inflataseems to resembleR.australianain that they both have ciliated branchiae on inner and outer margins. Another species,R.tuberculata, however, is similar in morphology to species of theR.glutaeacomplex,except that it has three small tubercles on the underside ofits lateral-frontal horns. Further study forR.australiana,R.glycera,R.inflata, andR.tuberculata, might uncover their divergences in phylogeny with the remainingRhynchospiospecies.
Water flow caused by ciliary action of nototrochs may help to transfer the oxygen dissolved in seawater into tubes of some genera of Spioninae (Simon,1967). InRhynchospioaff.asiatica, however, the water flow is stimulated by ciliary beating on the inner side of paired branchiae and nototrochs of each segment, which leads to directional movement of water along the dorsum of the worm. The water flow on the dorsum ofR. aff.asiaticacould be observed from directional movement of small particles suspended in the water tunnel, indicating that it may have a function of transporting the gametes from anterior and middle fertile segments to the posterior hatchery formed by elongated capillaries. Given the similarity in the arrangement of ciliary bands, it may be reasonable to infer that perhaps this water flow system exists in allRhynchospiospecies (perhaps excludingR.inflataandR.australianawhich have perpendicularly oriented branchial surfaces), which can be confirmed by observing live specimens in the future. This feature may cause adaptive changes,given that high fertilization efficiency might be desirable for these spionids that produce only a small amount of sperm and oocytes (Radashevsky, 2007a).
Our study has contributed to a better understanding of the phylogeny of Spionidae. The genera within Spionidae could be basically divided into two clades/subfamilies, i.e., Spioninae and Nerininae (Fig.5).Five genera (i.e.,Dispio,Malacoceros,Marenzelleria,Rhynchospio, andScolelepis) previously grouped within Nerininae due to their similarities in reproductive characteristics, such as thick-enveloped oocytes, ect-aquasperm, and broadcast spawning(Blake and Arnofsky, 1999), form a single clade with the other genera belonging to Spioninae with relatively high support values (BS=91; BPP=1) in this study. This phylogenetic result is supported by the morphological evidence that all the five genera have branchiae on most of the segments (referring to morphology Group B), similar to that ofSpioandMicrospio, which belong to Spioninae (Blake and Arnofsky, 1999). Similar close relationships between these genera (i.e.,Marenzelleria,Rhynchospio,Malacoceros,Dispio, andScolelepis) and those belonging to Spioninae were also showed in phylogenetic results of Abe and Sato-Okoshi (2021),while they were grouped within Nerininae according to morphological characteristics (i.e., number of eyes,pigmentation, and the position of gastrotrochs). In addition, our phylogenetic results agree with that of Abe and Sato-Okoshi (2021) by showing that the two spioniform genera (i.e.,TrochochaetaandPoecilochaetus) are clustered within the clade of Spionidae. The two phylogenetic analyses thus provided molecular evidences to support the proposal by Blake and Arnofsky (1999) that Trochochaetidae and Poecilochaetidae should be included within a broadly defined Spionidae.
The phylogenetic results (Fig.5) support the suggestion proposed by Hannerz (1956) and Radashevsky (2007b) that the ancestor of spionids might have thick-envelope oocytes, and thin-envelope oocytes is a derived trait in Spioninae. Given that thin-enveloped oocytes and introsperms have widespread occurrence in other annelid families(Jamieson and Rouse, 1989; Blake and Arnofsky,1999), they might have occurred more than once in the evolutionary history of annelids, and their evolution within Spioninae could be considered as adaptive changes to various brooding activities, such as having capsules/egg mass with brooding in tube or brooding on the dorsum of female. With three spioniform genera, i.e.,Spiophanes,Trochochaeta,andPoecilochaetus, at the base of the phylogenetic tree, our results further indicate that the common spionid ancestor might have no branchiae, and various arrangement patterns of branchiae (Groups A, B, C)together with various brooding/egg protection modes might be derived characteristics. This conclusion differs from the scenario proposed by Radashevsky(2007b) that the first spionid might have branchiae on most segments, which might be due to the basal position of the genera such asMarenzelleria,Dispio,Scolelepis, andMalacocerosin his cladistic analysis.Overall, our phylogenetic analysis and mapping of reproductive characteristics to the phylogenetic tree revealed a potential evolutionary history of reproductive characteristics in spionids.
Taxonomic key toRhynchospiospecies (modified from Radashevsky et al., 2014)


5 DATA AVAILABILITY STATEMENT
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
6 ACKNOWLEDGMENT
We thank Mengsheng ZHANG, Shaoyue LI,Shuhui WANG, Liangli TU, Fanqi ZHAO, Haixia ZHONG, Wenyao CUI, Xiangjun YANG, Chen CHEN, Congcong SHAO, Lin ZHU, and Ankang TENG from College of Marine Life Sciences, Ocean University of China for assistance with collectingRhynchospiospecimens. We also thank Dr. Vasily I.RADASHEVSKY for helpful information during the course of our study.
 Journal of Oceanology and Limnology2022年3期
Journal of Oceanology and Limnology2022年3期
- Journal of Oceanology and Limnology的其它文章
- Typhoon-induced wind waves in the northern East China Sea during two typhoon events: the impact of wind field and wave-current interaction*
- Effect of subsea dispersant application on deepwater oil spill in the South China Sea*
- Geochemical characteristics of cold-seep carbonates in Shenhu area, South China Sea*
- Examination of seasonal variation of the equatorial undercurrent termination in the Eastern Pacific diagnosed by ECCO2*
- Deviation of the Lagrangian particle tracing method in the evaluation of the Southern Hemisphere annual subduction rate*
- Immunostimulatory effect of quaternary degree and acetyl group of quaternized chitosan on macrophages RAW 264.7*