
Chinia gen. nov.—the second diatom genus simonsenioid raphe from mangroves in Fujian, China*
2022-06-16 02:49HuinaLINWeiweiWULinSUNAndrzejWITKOWSKIXiaoyeLIVishalPATILJunrongLIANGXuesongLIYahuiGAOChangpingCHEN
Huina LIN , Weiwei WU , Lin SUN , Andrzej WITKOWSKI , Xiaoye LI , Vishal PATIL ,Junrong LIANG , Xuesong LI , Yahui GAO ,,**, Changping CHEN ,**
1 School of Life Sciences and Key Laboratory of Ministry of Education for Coastal and Wetland Ecosystems, Xiamen University,Xiamen 361102, China
2 State Key Laboratory of Marine Environmental Science, Xiamen University, Xiamen 361102, China
3 Institute of Marine and Environmental Sciences, University of Szczecin, Mickiewicza 16a, PL-70-383 Szczecin, Poland
Abstract A new monotypic diatom genus Chinia gen. nov. from estuarine mangrove habitats in Fujian,China is described. Light, scanning and transmission electron microscopy observations ofits morphology show that this diatom differs from any established raphid diatom genus. The new genus is distinguished by a strongly undulate valve face, eccentric raphe canal, and perforated copulae. Its distinctly elevated and perforated raphe canal, classified as simonsenioid type, is supported by sparse infundibulum-like portulae, and the portulae are separated by large fenestrae. The valves of Chinia also have hymenate areolar occlusions and the raphe lacks a central nodule. However, unlike most Bacillariaceae with two plastids,including Simonsenia, Chinia has a single plastid similar to those in Entomoneis and Surirella. Despite the similarities to Simonsenia, Chinia features porous raphe canal and loose striation, which is different from those of Simonsenia that has a solid raphe canal and distinct rows of areolae. These distinctive cytological and morphological characters suggest an affinity of the new genus to the Bacillariaceae.
Keyword: marine diatoms; East China Sea; taxonomy; Bacillariaceae; new genus; simonsenioid raphe
1 INTRODUCTION
Mangrove forests are plant communities occurring in the intertidal zone of tropical and subtropical estuaries and coastal areas. Due to their transitional location between the ocean, estuary, and land, they support high levels of primary productivity and a rich biodiversity (de Albuquerque Ribeiro et al., 2019). As primary producers, diatoms play a vital part in the food web of the mangrove forests (Siqueiros-Beltrones and Sánchez-Castrejón, 1999; Gao, 2001).Several unique and rare species/genera of diatoms have been found (Siqueiros-Beltrones and Sánchez-Castrejón, 1999; Siqueiros-Beltrones and López-Fuerte, 2006; Seddon et al., 2014; Bąk et al., 2019; Li et al., 2020c) among the habitats associated with mangrove forests, in the plankton or the benthos on the sediment or the mangroves themselves (Du and Jin, 1983; Maples, 1983; Chen et al., 2010; Sahoo and Ansari, 2018; Sahoo et al., 2018). However, only very few publications reported new diatom taxa from mangrove forests along the coast of southeastern China (Chen et al., 2010; Li et al., 2020c).
In the last decade, a wider effort by diatomologists characterized the diatoms of the Chinese Pacific coast, adding to a number of published papers on local and regional scales, using morphological and molecular approaches (Liu et al., 2012; Li et al., 2015,2017, 2018a; Witkowski et al., 2016; Krzywda et al.,2019). This led to the description of several genera and dozens of species (new to science) and expanded the records of species occurring in various habitats of the Yellow Sea, the East China Sea, and the South China Sea, including epibiotic diatoms (Li et al.,2014, 2018b, 2020a, 2020b).
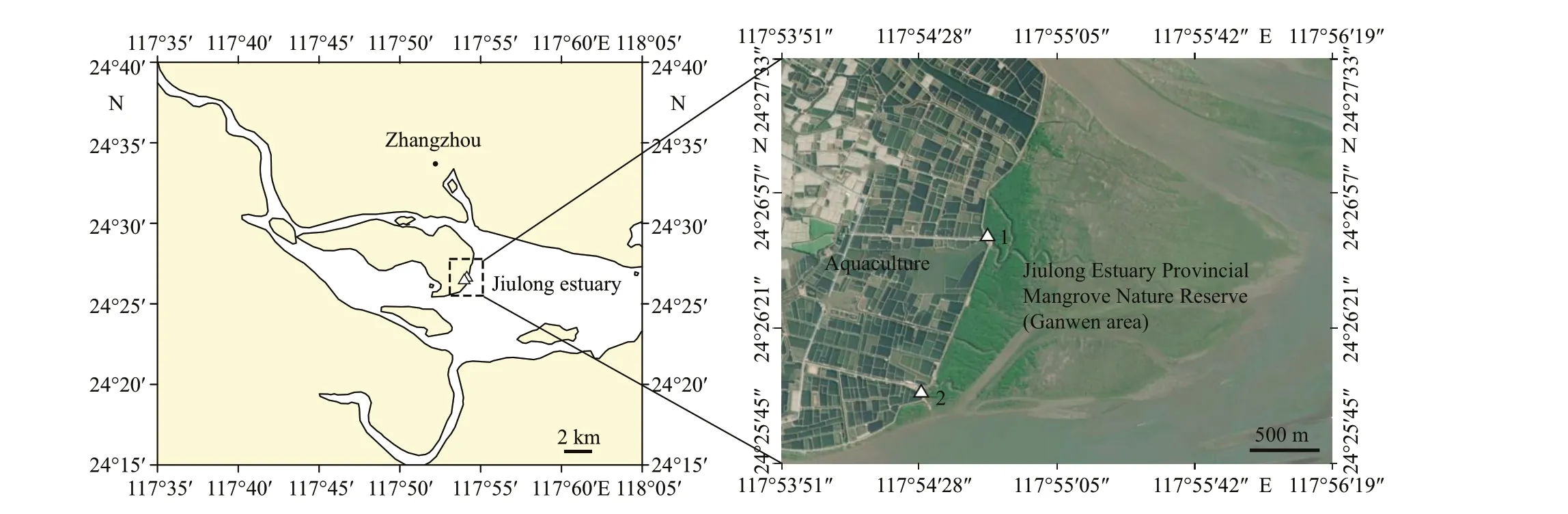
Fig.1 Location of the Jiulong Estuary Provincial Mangrove Nature Reserve (Ganwen area), Zhangzhou, Fujian Province,SE China
In this paper, a new diatom genus featuring the simonsenioid canal raphe type is described from mangroves in the Jiulong estuary on the Fujian coast in the South China Sea. The paper presents some cytological characteristics (plastid) and a detailed ultrastructure revealed by light microscopy (LM),scanning electron microscopy (SEM) and transmission electron microscopy (TEM) observations for this diatom genus, along with some basic information on ecology. At present, our attempts to obtain molecular data for this taxon were not successful, and its possible phylogenetic position is discussed based on its morphology.
2 MATERIAL AND METHOD
The study site is located in the mangrove forest in the Jiulong estuary, the Jiulong Estuary Provincial Mangrove Nature Reserve (Ganwen area), in Longhai district of Zhangzhou city in Fujian Province,Southeastern (SE) China (24°26′4″N-24°26′45″N,117°54′29″E-117°54′46″E; Fig.1). The mangrove ecoregion features a semidiurnal tide ranging from 1.0 to 5.5 m (Chen et al., 2017). The ecoregion was listed as one of the key protected wetlands in China,with an area covering about 420.2 hm2and large aquaculture facilities located in the vicinity.
Samples of mangrove leaves (KandeliaobovataSheue, H. Y. Liu & J. Yong), fiddler crabs (Ucaarcuatade Haan), seaweed (IshigeokamuraiYendo),mangrove prop roots (K.obovata), stones, shells, and sediment were collected with forceps in high tide zone during low tide in May 2018, May 2019, August 2019, and October 2019. Water salinity, pH, and temperature were measured in situ using a Horiba U-52 portable probe (Japan). The samples were kept in zip lock bags, cryopreserved and taken to the laboratory.
In laboratory, samples were rinsed twice with filtered seawater through 0.45-μm filters to wash offplanktonic species. Then, diatoms were removed from different biotic substrates in ultrasound bath(300 W for 30 s) and washed four times with the filtered seawater. Diatom samples were then boiled at 100 ℃ for 20 min in concentrated HCl to remove organic matter of the cells, and washed with distilled water 10 times until neutral pH was reached.Permanent microscope slides of the processed samples were mounted with Naphrax for LM observations under the Olympus BX51 microscope with a DP71 digital camera system (Olympus, Japan)and a 100× oil immersion objective. Cleaned diatom frustules for SEM observations were mounted onto small pieces of coverslip or 0.22-μm pore size filters,air-dried and fixed on aluminum stubs, then coated for 60 s with 4-5 nm of gold palladium (working distance:60 mm, current: 30 mA) in the Leica EM CPD300(Germany). SEM observations were conducted on the JEOL JSM-6390A (Japan) and FEI Quanta 650 FEG Scanning Electron Microscopy (USA) at 20 kV.Frustules were mounted onto a copper grid and airdried for TEM observations using the Hitachi HT-7800 (Japan) at 80 kV. Three-dimensional (3D) model images ofChinialonghaiensisgen. et sp. nov. were illustrated with the SolidWorks. Samples and permanent slides were deposited at the School of Life Sciences, Xiamen University, China. Diatom terminology followed Round et al. (1990) and Witkowski et al. (2015).
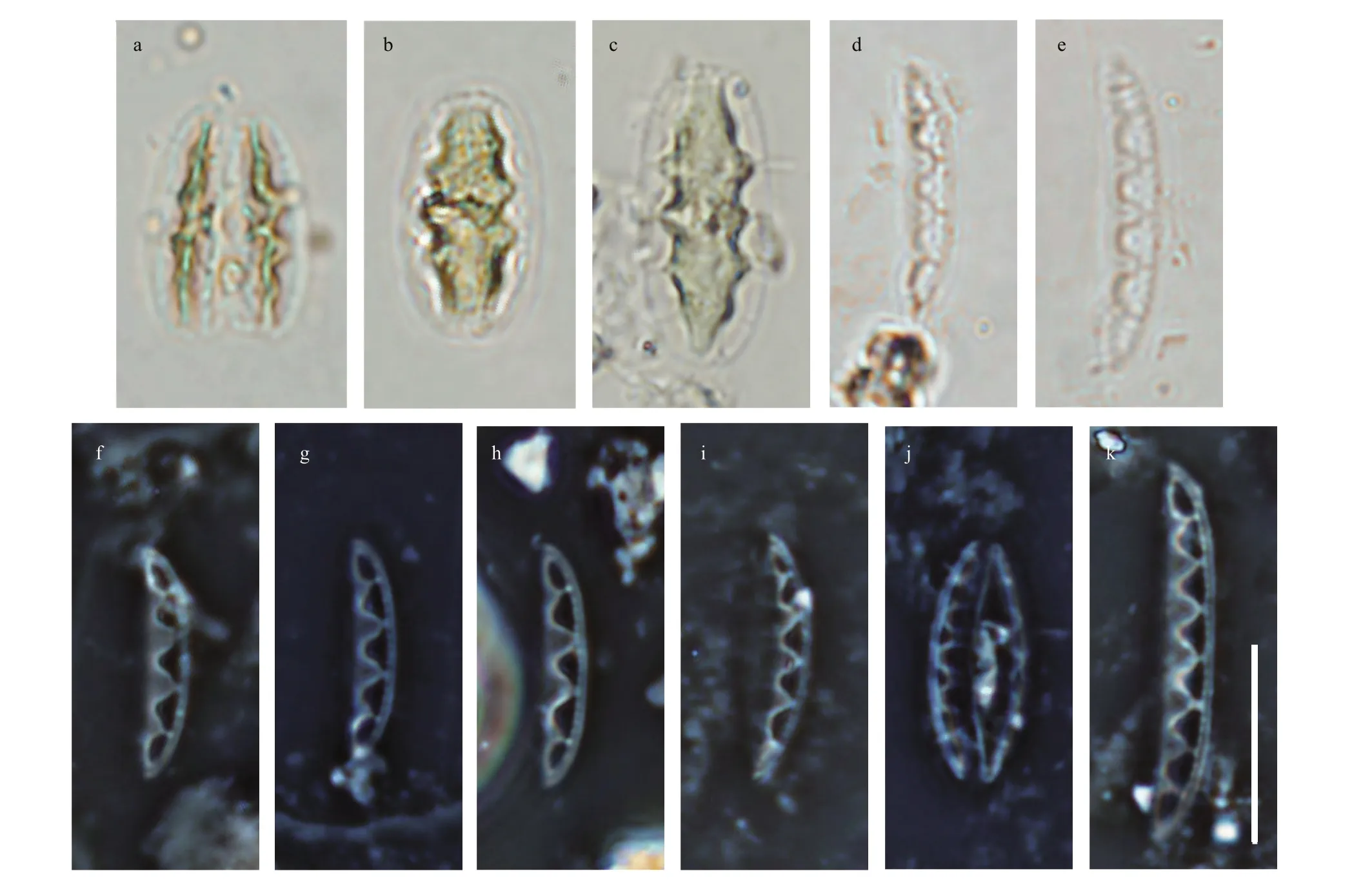
Fig.2 Light micrographs of Chinia longhaiensis gen. et sp. nov. showing the cell and valve size diminution series with the distinct fenestrae features
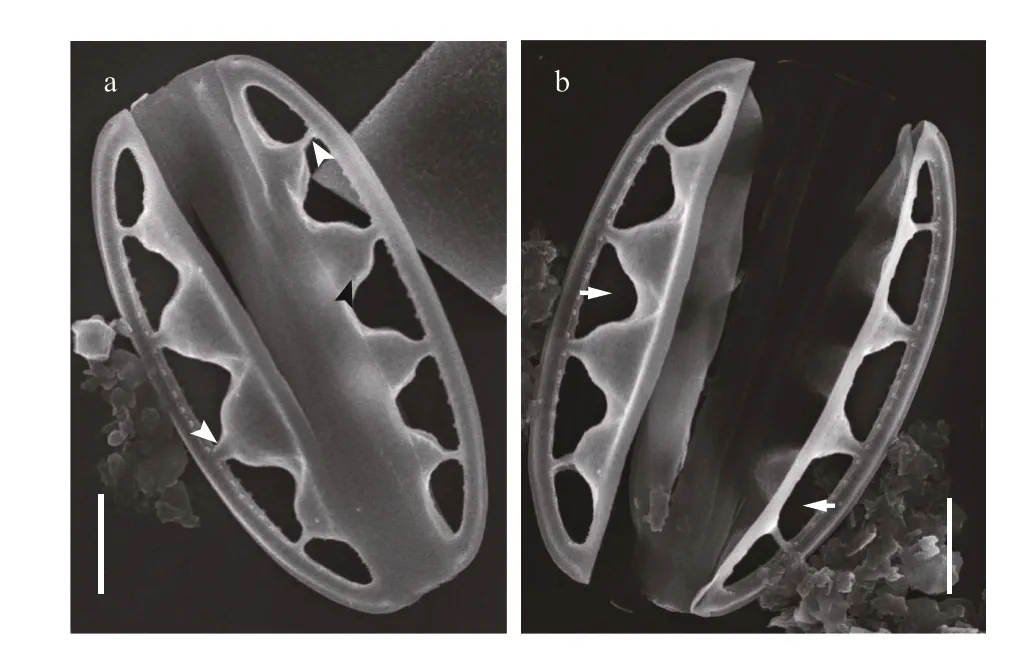
Fig.3 SEM images of Chinia longhaiensis gen. et sp. nov.,showing the outline of whole frustule exterior with large fenestrae and fenestral bars
3 RESULT
3.1 Taxonomy
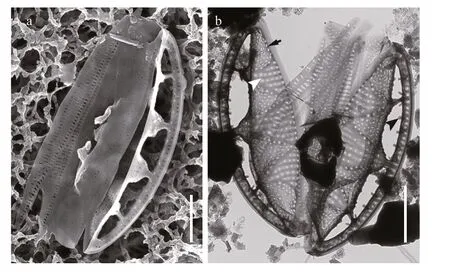
Fig.4 SEM and TEM images of Chinia longhaiensis gen. et sp. nov. showing the girdle bands of cell
The new genus was observed from each type of sampled substrates. The taxon was conspicuous due to its characteristic elliptical frustules and large fenestrae (cf. Figs.2-7).
Systematics:
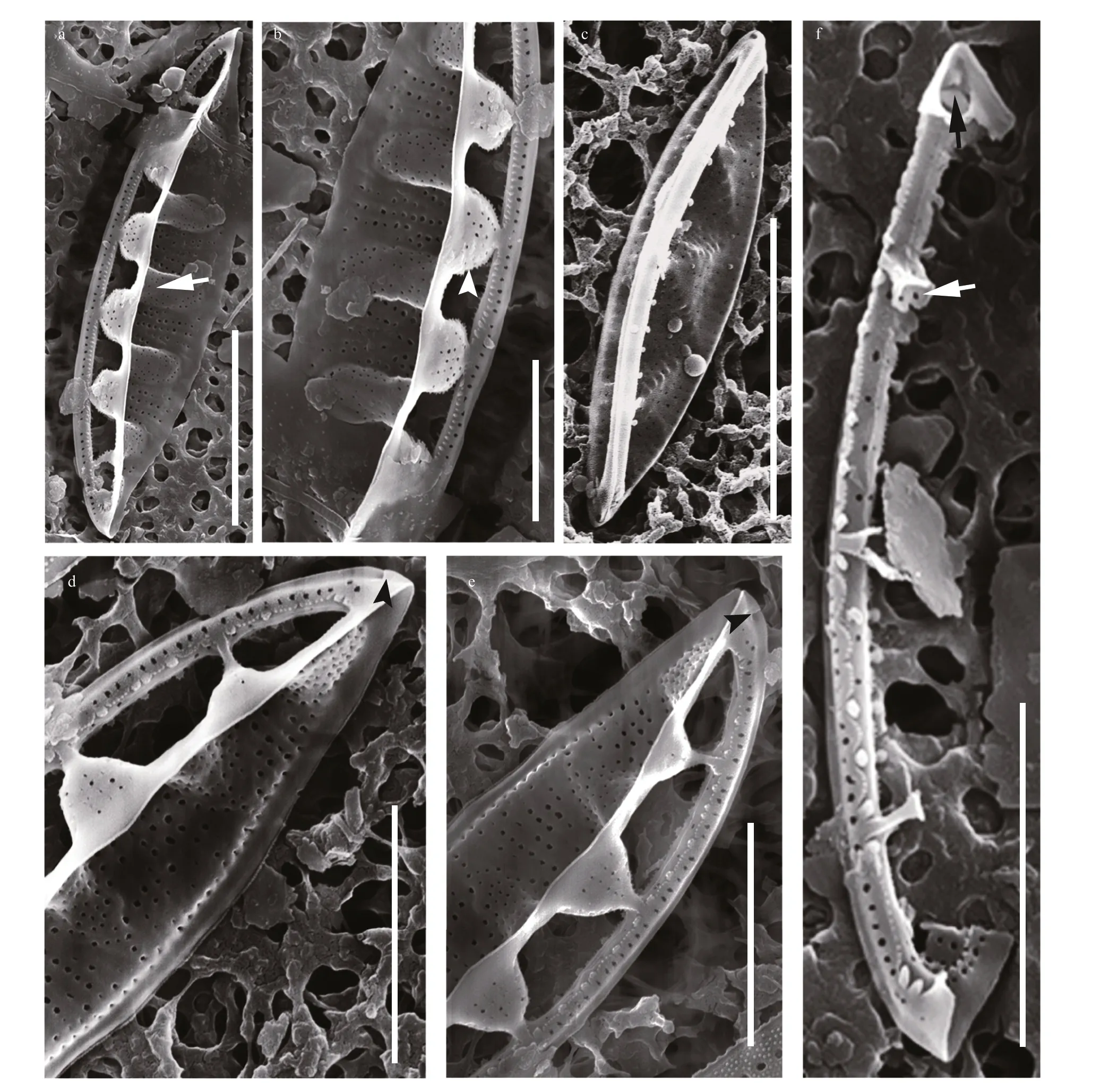
Fig.5 SEM images of Chinia longhaiensis gen. et sp. nov.
Class: Bacillariophyceae Haeckel
Subclass: Bacillariophycideae D. G. Mann
Order: Bacillariales Hendey
Family: Bacillariaceae Ehrenberg
ChiniaY. Gao, H. Lin, C. Chen et A. Witkowski gen. nov. (Figs.2-3)
Description: Frustules elliptical in girdle view with a single plate-like plastid (Fig.2). Valves lanceolate with acutely rounded apices, valve face distinctly undulate (Fig.3). Girdle relatively broad,composed of densely perforated copulae with apically oriented rows of pores (Fig.4). Raphe-bearing canal perforated, positioned on the elevated keel, moderately eccentric (Figs.5a & c, 6b & c). Raphe slit continuous,without central nodule (Fig.6a). External apical raphe endings strongly bent in one direction (Fig.5d-e),internally ending in somewhat elevated helictoglossae(Fig.5f). Raphe canal connected to cell interior through 3-8 infundibulum-like portulae (Figs.2-7).Parallel transapical striae composed of irregularly distributed small, circular areolae, observable by TEM and SEM (Figs.4-6). When observed by LM,cells with a single plate-like plastid.

Fig.6 TEM (a-b, d-e) and SEM (c) of Chinia longhaiensis gen. et sp. nov.
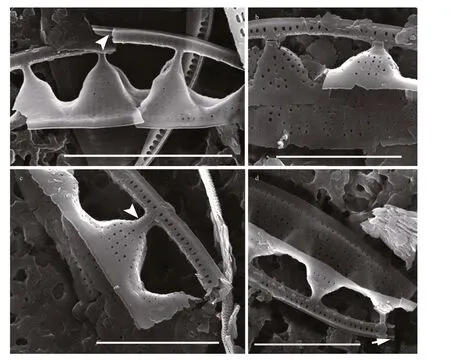
Fig.7 SEM images of broken Chinia longhaiensis gen. et sp. nov. valves
Etymology: The genus is dedicated to the esteemed biologist and diatomologist Professor Dexiang JIN (CHIN T. G., 1910-1997) from Xiamen University, China, for his contributions to the establishment of the Diatom Laboratory at Xiamen University in the 1930s and in recognition of his achievements in both the marine diatom and biological sciences.
Generitype:ChinialonghaiensisY. Gao, H. Lin,C. Chen et A. Witkowski sp. nov.
Taxonomic note: The new genus resemblesSimonseniawith the simonsenioid type of canal raphe elevated and supported on portulae; the raphe slit without a central brake (central nodule) and perforated fibulae. However, they differ in terms of the perforated raphe canal, reduced fenestral braces and densely perforated copulae inChiniagen. nov. The two genera also differ in terms of plastids, with two apically positioned plastids inSimonsenia(Witkowski et al.,2015; Kim et al., 2019) and one plate-like plastid inChinia(this study).
ChinialonghaiensisY. Gao, H. Lin, C. Chen et A.Witkowski sp. nov.
Description: Frustules solitary, small, elliptical in girdle view with distinct fenestrae (Fig.2), and one plate-like axial plastid (Fig.2b-c). Valves (n=25)lanceolate with acutely rounded apices 8.0-17.0-μm long, 2.1-3.6-μm wide. Valve surface undulate with distinct portulae connecting the raphe-bearing canal to the inner cell. Transapical striae not resolvable by LM.
3.2 Morphology observed by EM
When observed in a girdle view, the valves show a series of elevated conical parts (infundibulum-like)supporting the portulae alternating with depressed ones (Figs.2-7) and with a distinct hyaline mantle(Fig.4). The elevated valve parts constitute the striated valve surface, whereas the depressed, finely perforated parts correspond to fibulae (Fig.5a). The raphe canal is elevated above the valve surface, supported by portulae and moderately eccentric. Careful examination of SEM images reveals reduced fenestral bars (braces) that adhere tightly to the external surfaces of portula tubes (Fig.3a). The raphe canal external surface is finely perforated on each side (40-62 in 10 μm;n=15) and somewhat displaced toward the proximal mantle, with a raphe slit continuous from pole to pole (Figs.5-6) and external apical ends strongly bent on the same side, ending in small and distinct grooves at the poles (Figs.5d-e, 6d).Internally, the raphe terminates in small but distinct helictoglossae (Fig.5f). The raphe canal is separated from the valve surface by distinct fenestrae (4-5 in 10 μm;n=15) (Fig.3). The raphe canal is ornamented with spinules and lace-curtain-like siliceous deposits,and equipped with two or three spinules at the poles(Figs.5f & 6c). Tiny spinules positioned over the outer wall of the canal point toward the valve face(Figs.4b, 5c & f, 6c). Transapical striae are composed of small hymenate, irregularly distributed areolae(Fig.6e). External areolae are positioned in shallow parallel groves (Figs.4b, 5, 6) (42-56 in 10 μm;n=10).Striae forming areolae are distinctly larger than those of perforated plate-like fibulae (Fig.4b).
We were unable to observe the portulae and fibulae from the valve interior in a position perpendicular to the electron beam due to the geometry of the valve forming a relatively high cone with an elevated raphe canal that is unstable in SEM/TEM preparations. For this reason, we have illustrated these structures using 3D images with the SolidWorks (Fig.8) compiled from numerous SEM and TEM images obtained during this study. The peculiar fenestral bars (braces)adhere to the external surface of the portulae (Fig.8a;see also Fig.3a). Internally, plate-like fibulae are perforated near the portula tube-passage (Fig.8c; see also Fig.5a-e). Portulae turn into passages connecting the valve interior to the canal raphe (Fig.8d-f; see also Fig.5f).
Holotype: Slide No. GWH18050204 (holotype designated here on Fig.2h) housed at the School of Life Sciences, Xiamen University, Xiamen, China(see Figs.2-7).
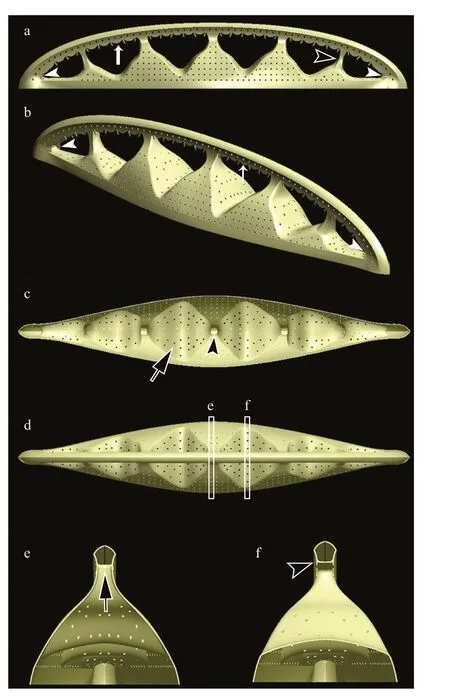
Fig.8 Three-dimensional models of Chinia longhaiensis gen. et sp. nov. illustrated by the SolidWorks
Isotype: Slide No. SZCZ27378 housed in the Diatomological Collection of Andrzej Witkowski at the Institute of Marine and Environmental Sciences,University of Szczecin, Szczecin, Poland.
Type habitat: Jiulong estuary mangrove area in Longhai district, the city of Zhangzhou, Fujian Province,China (24°26′4″N-24°26′45″N; 117°54′29″E-117°54′46″E), collector: Huina LIN, May 2, 2018.
Etymology: The specific namelonghaiensisrefers to the sampling site located in Longhai district, the city of Zhangzhou, Fujian Province, Southeastern China.
Ecology: Currently,Chinialonghaiensiswas only observed at this particular sampling site. The new taxon was found in four benthic habitats (mangrove leaves, fiddler crabs, seaweeds, and mangrove prop roots) collected from the study area. The taxon inhabits a brackish-water environment with salinities ranging from 8 to 20, in the neutral pH range (7.12 to 7.84), with dissolved oxygen (DO) ranging from 4.59 to 7.68 mg/L and water temperature ranging from 26 to 30 °C. The associated diatom flora included:SimonseniaaveniformisA. Witkowski, A. Gomes &E. Gusev,Psammodictyonpanduriformevar.continuum(Grunow) P. Snoeijs,Tryblionellagranulata(Grunow) D. G. Mann,TryblionellapunctataW. Smith,SurirellaatomusF. Hustedt,DiploneisbombusC. G. Ehrenberg,Pseudofallaciatenera(Hustedt) Y. Liu, J. P. Kociolek & Q. Wang,Luticolasp.,Naviculasp.,Amphorasp., andPleurosigmasp.; they are dominant diatoms in the examined samples.
Taxonomic note: At a first glance,Chinialonghaiensissp. nov. slightly resemblesSimonseniacf.paucistriataandS.paucistriata(Kim et al., 2019),marine species from the Korean part of the Yellow Sea coast and the Texas coast of the Gulf of Mexico.They have a similar shape, canal raphe position, and portulae.SimonseniapaucistriataandS. cf.paucistriatain fact show morphological features of the valve face that resembleC.longhaiensisas they have an undulate valve surface and solitary rows of areolae in the transapical striae (Kim et al., 2019).However, the main differences between these species are the size of the fenestrae and the width of the girdle;the size and the length are much larger inC.longhaiensisthan in anySimonseniaspecies(Lange-Bertalot, 1979; Witkowski et al., 2014, 2015;You et al., 2016; Kim et al., 2019), includingS. cf.paucistriataandS.paucistriata, whereasC.longhaiensislacks the free-standing, rib-like,supporting braces for the raphe canal which are known only inSimonseniaspp. (Kim et al., 2019; this paper).
4 DISCUSSION
No species resemblingC.longhaiensishave been recorded so far. In this context,Chiniagen. nov. is a surprising discovery for having its novel combination of cytological and morphological characteristics.
4.1 Comparison with other canal raphe-bearing genera
Compared to other canal raphe-bearing diatoms,Chiniahas similar simonsenioid raphe ofSimonsenia,but differs from the genusNitzschiain the raphe system, which applies in particular to the marginal(diagonal) position of the raphe canal within the frustule. The girdle that in both genera is composed of several copulae each bearing one to two rows of pores. To a certain extent, the fibulae ofChiniaandSimonseniaare slightly similar. The fibulae in the former genus are perforated and spacious with a slightly conical surface but are well separated from the valve surface, perforated and horizontally positioned in the latter genus (Lange-Bertalot, 1979;Witkowski et al., 2014). Although both genera have an elevated raphe canal, the raphe canal inChiniais perforated with rows of solitary pores, whereas that inSimonseniais closed and forms an unperforated tube.These two genera differ in the development of fenestral bars (braces) that are clearly visible inSimonsenia. The fenestral bars play apparently an important role in supporting the raphe-bearing tube,and the reduced fenestral bars adhering to the external portulae surface inChiniaare merely visible by careful examination (Witkowski et al., 2015; this study). They also differ in terms of plastid morphology:two apically positioned plastids inSimonsenia(Witkowski et al., 2015; Kim et al., 2019) and one plate-like plastid inChinia(this paper).
As shown by Witkowski et al. (2015) and You et al.(2016), the simonsenioid raphe system inSimonseniaandChinia(this study) bears some similarity to the surirelloid type in the apical part of the valve; however,this similarity only applies to the external view, as the surirelloid raphe-bearing tube is connected to the valve surface, while simonsenioid raphe-bearing tubes are suspended. The surirelloid raphe system also differs from that ofChiniain the wing of the raphe, which is supported by portulae (=alar canals) at distinct locations inChiniabut connected to the rest of the valve in the surirelloid system (Krammer and Lange-Bertalot,1987; Round et al., 1990; Ruck and Kociolek, 2004;Ruck and Theriot, 2011). In contrast to the nitzschioid morphology, where the raphe canal is more or less open toward the valve interior, the separation of the raphe canal by large fenestrae makesChiniaeasy to distinguish from mostNitzschiaspecies (Krammer and Lange-Bertalot, 1987; Round et al., 1990; Ruck and Theriot, 2011; Trobajo et al., 2013; Mann and Trobajo,2014). Table 1 presents a summary of the morphological features of Chinia gen. nov. and a comparison of the new genus with the established genera including,among others, Simonsenia, Nitzschia, and Surirella.
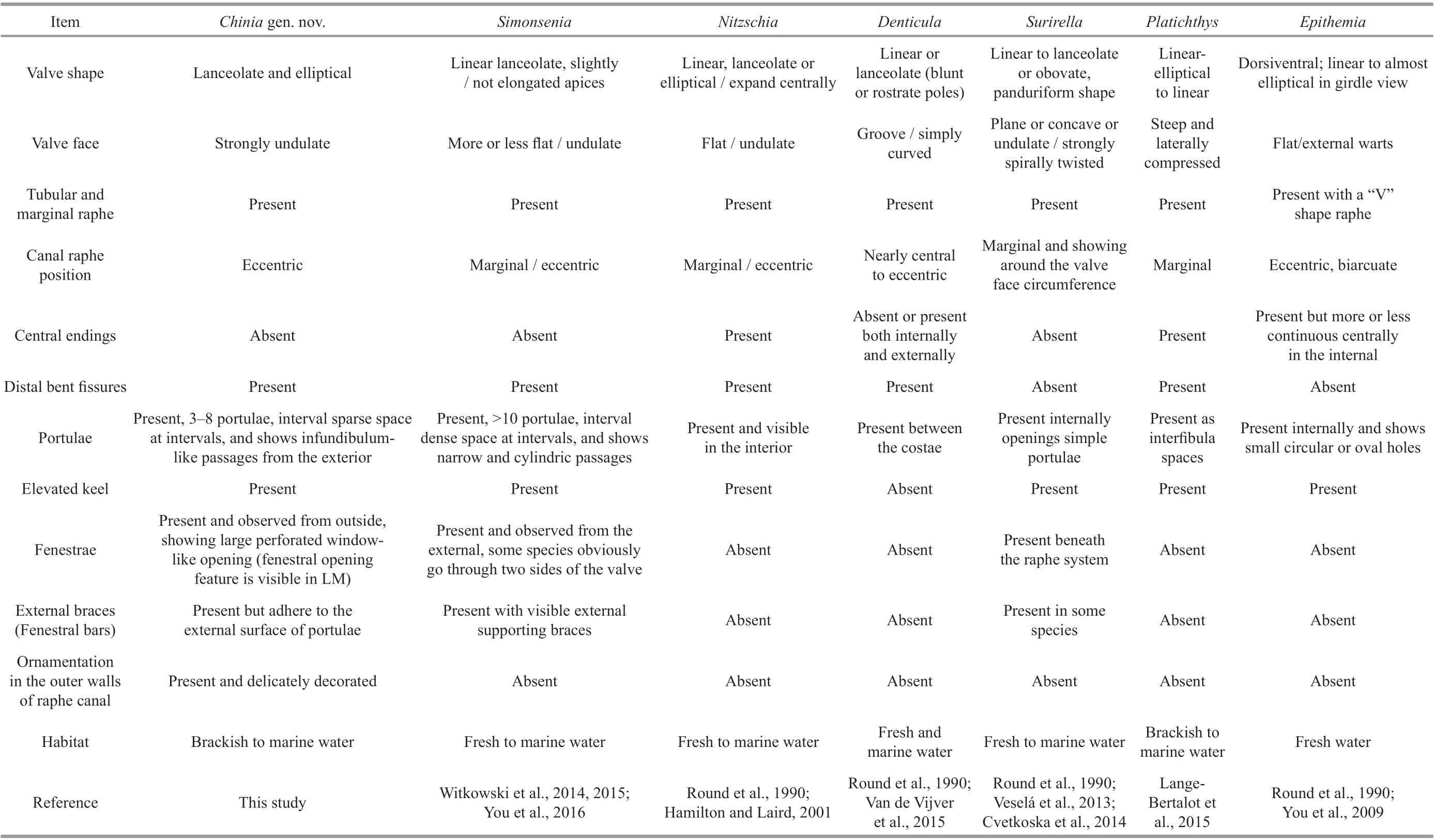
Table 1 Comparison in morphology of Chinia gen. nov. and established genera Simonsenia, Nitzschia, Denticula, Surirella, Platichthys, and Epithemia
4.2 Classification of Chinia
The longitudinal slit called a “raphe” across the valve component of the siliceous exoskeleton of diatoms serves as a diagnostic character for a monophyletic group of diatoms referred to as “raphid pennates”. It is this raphe that allows raphid pennate diatoms to actively glide over various substrates. This raphe system takes several morphological forms across the raphid diatoms, including the “canal”raphe, where the raphe is associated with a canal-like structure supported by transverse siliceous bridges(fibulae). The canal raphe was considered to be the result of a single evolutionary event for decades(Simonsen, 1979; Sims and Paddock, 1982; Round et al., 1990), though more recently, molecular data derived from nuclear and chloroplast genes (Ruck and Theriot, 2011) suggested that the canal raphe system developed independently in Bacillariales,Rhopalodiales, and Surirellales.
Three distinct morphological categories of canal raphe were defined by Ruck and Kociolek (2004) and Witkowski et al. (2015). The nitzschioid type shows a simple canal and fibulae on a relatively simple keel,positioned on the valve at least somewhat eccentrically and open to the valve interior. This type of canal raphe can be found in the generaBacillaria,Hantzschia,Nitzschia,Pseudonitzschia,Fragilariopsis, andDenticula. The surirelloid type presents a wing elevated on a shallow or deep keel, with the raphe canal connected to the valve interior by narrow tubes(portulae) and is found in the generaSurirella,Stenopterobia,Petrodictyon, andPlagiodiscus. The simonsenioid type displays an elevated raphe canal,which is either strongly or moderately eccentric and runs along the apical axis of the frustule on a wing(ala), supported by portulae (alar canals) and two rows of fenestral bars (braces). This canal raphe type has so far been found inSimonseniaonly (Lange-Bertalot, 1979; Witkowski et al., 2015; Kim et al.,2019).
While the surirelloid canal raphe type appears to be homogenous among the taxa belonging to Surirellales(Ruck and Kociolek, 2004), Bacillariales show a wide variation in the raphe canal position and ultrastructure,representing two types of raphe systems: nitzschioid and simonsenioid (Witkowski et al., 2015; Kim et al.,2019; Mann et al., 2021). Despite the differences in the structure of raphe canal systems outlined above,Simonseniais well positioned within Bacillariceae. In one of the largest published DNA sequence datasets of the family (Mann et al., 2021),Simonseniaclade is derived within Bacillariaceae. This relationship is congruent with other molecular phylogenies of the family constructed by the use of plastid-encoded markers (Witkowski et al., 2015; Kim et al., 2019).
While we were unable to obtain any molecular data fromChinia, we could propose a classification forChiniabased on the simonsenioid canal raphe morphology it exhibits, which leads us to tentatively include the new genusChiniain the Bacillariaceae,withSimonsenia. It does not escape our notice thatChiniais unique in the Bacillariaceae in having a single plate-like plastid, which is typical toSurirellaandEntomoneis(Round et al., 1990).Simonseniaitself, like most of the Bacillariaceae, is characterized by anterior and posterior plastids (Witkowski et al.,2015; Kim et al., 2019).
4.3 Distribution and ecology
At present,Chinialonghaiensiswas observed in our mangrove zone collections only, though it was present in all sampled habitat types. The taxon did not reach a high relative abundance (approximately 1%)in any of the samples. It is not clear how water salinity affects its distribution asC.longhaiensiswas sampled in the salinity ranging from 8 to 20; however, the taxon developed well under our laboratory culture conditions at salinity up to ca. 35. This can be interpreted as an indication of euryhaline autecology.Chinialonghaiensiswas only present in sonification samples from biological substrates e.g. mangrove prop roots, mangrove leaves, fiddler crabs, and seaweeds, but not present in other samples such as stones, shells, or sediments. Although no images ofChiniafrustules attached to the substrates studied were made, we consider that, in terms of functional ecology, it may represent a marine epibiontic diatom.To the best of our knowledge, a similar diatom has never been published even as an unidentified taxon.The discovery ofC.longhaiensisis a contribution to the variation in the canal-raphe structure and adds a new genus with simonsenioid canal raphe.
5 CONCLUSION
A novel epibiontic diatom genus,Chiniagen. nov.,is described with a new species,Chinia longhaiensissp. nov., from estuarine mangrove habitats in Fujian,China. The new genus is distinguished by its single plate-like plastid, and disctinct morphological characters showing simonsenioid type raphe canal with sparse infundibulum-like portulae and large fenestrae. These distinctive features ofChiniasuggest its affinity to the Bacillariaceae.
6 DATA AVAILABILITY STATEMENT
All data generated and/or analyzed in this study are available from the corresponding author on reasonable request.
7 ACKNOWLEDGMENT
We greatly appreciate the useful advice on this new genus provided by Prof. Tamotsu NAGUMO at the Nippon Dental University in Japan; Dr. Xueliang HOU at the School of Life Sciences, Xiamen University for his helpful suggestions on the nomenclature of this new genus and species, and Dr.Caiming WU, Dr. Luming YAO, and Ms. Jie LIU from Xiamen University for their assistance with EM observations. We are grateful to anonymous reviewers for their comments and suggestions which greatly improved this manuscript. The authors greatly appreciate English language edits and critical comments by Dr. Matt ASHWORTH, Texas State University at Austin.
 Journal of Oceanology and Limnology2022年3期
Journal of Oceanology and Limnology2022年3期
- Journal of Oceanology and Limnology的其它文章
- Typhoon-induced wind waves in the northern East China Sea during two typhoon events: the impact of wind field and wave-current interaction*
- Effect of subsea dispersant application on deepwater oil spill in the South China Sea*
- Geochemical characteristics of cold-seep carbonates in Shenhu area, South China Sea*
- Examination of seasonal variation of the equatorial undercurrent termination in the Eastern Pacific diagnosed by ECCO2*
- Deviation of the Lagrangian particle tracing method in the evaluation of the Southern Hemisphere annual subduction rate*
- Immunostimulatory effect of quaternary degree and acetyl group of quaternized chitosan on macrophages RAW 264.7*