
Taxonomy and toxin profile of harmful benthic Prorocentrum(Dinophyceae) species from the Xisha Islands, South China Se a*
2022-06-16 02:49JianZOUQunLIHuiLIUYingLIULifenHUANGHaiyanWUJiangbingQIUHuaZHANGSonghui
Jian ZOU , Qun LI , Hui LIU , Ying LIU , Lifen HUANG , Haiyan WU ,Jiangbing QIU , Hua ZHANG ,5,**, Songhui LÜ ,6,**
1 Research Center of Harmful Algae and Marine Biology, College of Life Science and Technology, Jinan University, Guangzhou 510362, China
2 State Environmental Protection Key Laboratory of Drinking Water Source Management and Technology, Shenzhen Academy of Environmental Science, Shenzhen 510008, China
3 Yellow Sea Fisheries Research Institute, Qingdao 266071, China
4 College of Environmental Science and Engineering, Ocean University of China, Qingdao 266100, China
5 Shenzhen Academy of Environmental Science, Shenzhen 510008, China
6 Southern Marine Science and Engineering Guangdong Laboratory, Zhuhai 519000, China
Abstract Some benthic Prorocentrum can produce okadaic acid (OA) and dinophysistoxins (DTXs) that cause diarrheic shellfish poisoning (DSP) in humans. The diversity and toxin profiles (OA and DTXs) of benthic P rorocentrum were investigated in the Xisha Islands, South China Sea. The benthic Prorocentrum was identified by both morphological features and molecular phylogenies. Morphologies were examined by light, fluorescence, and scanning electron microscopy, and phylogenetic analyses were based on partial large subunit (LSU) rDNA and ITS1-5.8S-ITS2 (ITS) region. Seven Prorocentrum species including P. borbonicum, P. caipirignum, P. concavum, P. elegans, P. cf. emarginatum, P. lima complex, and P.rhathymum were identified in Xisha Islands. Among them, P. borbonicum and P. elegans were recorded in Chinese waters for the f rist time. OA and DTXs contents of seven benthic Prorocentrum species were evaluated based on liquid chromatography-tandem mass spectrometry (LC-MS/MS). All Xisha Islands strains of P. lima complex produced OA at contents ranging from 1 663 to 3 816 fg/cell. P. caipirignum also generated OA at 407 fg/cell, but other five species had no detectable toxins. Besides, interestingly, two strains of P. lima complex produced DTX-1 only (74 and 183 fg/cell) and another two strains generated an isomer of OA and DTX-2. Our findings provided insight into the biodiversity of benthic Prorocentrum in the Xisha Islands and pointed out the potential risk of DSP in this area.
Keyword: Prorocentrum; diversity; dinoflagellate; okadaic acid; dinophysistoxins; Xisha Islands
1 INTRODUCTION
Benthic dinoflagellates have drawn significant attention in recent decades (Berdalet et al., 2012).Some species could produce toxins and then cause harm to the health of aquaculture, benthic ecosystem,and humans (Berdalet et al., 2017). Even in the recent two decades, the blooms of benthic dinoflagellates occur more often and broadly, which further threaten public health (Accoroni and Totti, 2016; Zou et al.,2020). Among these dinoflagellates, species ofProrocentrumC.G. Ehrenberg is abundant and distributes globally, and these dinoflagellates could inhabit the planktonic and benthic environment(Boisnoir et al., 2019). So far, approximately 71 species of this genus were identified from the tropic to temperate regions (Murray et al., 2009; Luo et al.,2017; Chomérat et al., 2019). The planktonicProrocentrummicansC.G. Ehrenberg was first identified by Ehrenberg in 1834 and was considered as a type species ofProrocentrum(Ehrenberg, 1834).AmongProrocentrum, about 35 species ofProrocentrumepiphytically inhabit the benthic environment including macrophytes, dead corals,sediments, and so on (Hoppenrath et al., 2013;Rodríguez et al., 2018; Lim et al., 2019).
TheProrocentrummorphologically possesses two large thecal plates separated by a sagittal suture, a topmost periflagellar area composed of several tiny platelets and two pores (Fensome et al., 1993;Hoppenrath et al., 2013). Cell shape and size, the relative location of the nucleus, thecal surface micromorphology, thecal pore and intercalary band ornamentations and, most importantly, the architecture of periflagellar area are used for the morphological identifications of benthicProrocentrumspecies(Hoppenrath et al., 2013; Lim et al., 2019). However,the shape and size of cell and the numbers and patterns of pore vary among several species from different geographical regions, which cause difficulties for the taxonomy ofProrocentrum(Ten-Hage et al., 2000a;Hoppenrath et al., 2013). In general, the periflagellar area is located in the apical area ofProrocentrumand the ultrastructure analysis reveals that it is composed of a flagellar pore, an accessory pore, and several tiny platelets (Hoppenrath et al., 2013; Chomérat et al.,2019). The ultrastructure of periflagellar area is relatively conservative and then is used for the identification ofProrocentrum(Hoppenrath et al.,2013). However, to date, the details of the periflagellar area of certain species likeP.borbonicumL. Ten-Hage, J. Turquet, J.-P. Quod, S. Puiseux-Dao & Couté andP.elegansFaust are very significant but remain unclear (Faust, 1993; Hoppenrath et al., 2013; David et al., 2014).
The morphology ofProrocentrumis relatively homologous and has apparent plasticity (Hoppenrath et al., 2013; Zhang et al., 2015). Consequently, the misidentification ofProrocentrumcan be easily caused. Therefore, molecular phylogenetic analyses are necessary for the identifications of previously described species and descriptions of new species.For example, Mohammad-Noor et al. (2007)reexaminedP.arabianumMorton et Faust isolated from original stations and suggested it was synonymized withP.concavumFukuyo based on morphologies and phylogenies.P.maculosumM. A.Faust was synonymized with the toxic speciesP.hoffmannianumM. A. Faust (Rodríguez et al.,2018). The phylogenetic analysis is essential to well understand the genetic diversity inProrocentrum.Recently, a taxonomical study in Lesser Antilles,eastern Caribbean Sea revealed phylogenies of benthicProrocentrummainly divided into six clades based on large subunit (LSU) (D1-D2) sequences.Among these clades,P.limacomplex andP.fukuyoicomplex, which showed high genetic diversity, had six and five subclades, respectively (Chomérat et al.,2019). In addition, a recent study in Japan revealedP.limacomplex had high genetic diversity and existence of cryptic species (Nishimura et al., 2020a). Hence, it is necessary to research the phylogenies of benthicProrocentrumin a wide area.
Interest in marine epibenthicProrocentrumhas grown in recent years because most of them are potential toxins producers (Zhou and Fritz, 1993; Lim et al., 2019) or harmful to marine invertebrates (Zou et al., 2020). Among the benthicProrocentrum, nine species of them produce toxic polyether compounds like okadaic acid (OA) and/ or its related derivatives dinophysistoxins (DTXs), borbotoxins, and prorocentrolides (Ten-Hage et al., 2000a; Amar et al.,2018; Lim et al., 2019; www.marinespecies.org/).These toxic compounds can contaminate shellfish and subsequently cause shellfish poisoning in humans(Morton, 1998; Ten-Hage et al., 2000b). In addition,some species (e.g.,P.concavum) have no detectable diarrheic shellfish poisoning (DSP) toxins but harm marine invertebrates (Zou et al., 2020). DSP toxins are lipophilic and frequently cause severe poisoning events. OA and its DTX derivatives can induce diarrhea, nausea, vomiting, and abdominal pain in humans (Toyofuku, 2006). OA was first extracted from an oceanic sponge. It is a polyether derived by a 38-carbon fatty acid and causes strong inhabitation of protein phosphatases-1 and -2A activities (Goris et al., 1989). So far, OA producers ofProrocentrumincludeP.limacomplex (Ehrenberg) F. Stein,P.caipirignumSilvia M. Nascimento,P.hoffmannianum,P.rhathymumA. R. Loeblich Ⅲ, Sherley & Schmidt,P.concavum,P.faustiaeS. L. Morton,P.leveM. A.Faust, Kibler, Vandersea, Tester & Litaker, andP.fukuyoi(Yasumoto et al., 1987; Dickey et al., 1990;Morton, 1998; Ten-Hage et al., 2000b; Marasigan et al., 2001; An et al., 2010; Luo et al., 2017; Nishimura et al., 2020b). Among them, the toxicP.limacomplex are cosmopolitan species and extensively distributed from tropical to temperate areas (Yasumoto et al.,1987; Bouaı¨cha et al., 2001; Vila et al., 2001; Luo et al., 2017).
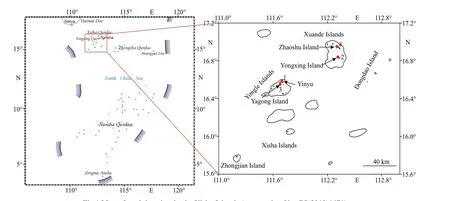
Fig.1 Map of studying sites in the Xisha Islands (map review No. GS(2019)1671)
Considering the potential threat of benthicProrocentrumto public health and fishery of Xisha Islands in China, the investigation on the biodiversity and toxicity of these species is urgently required in this area where relatively little information related to benthic dinoflagellates is available now. Here, we identified benthicProrocentrumspecies based on light, epifluorescence and scanning electron microscopy coupled with molecular phylogeny.Besides, the toxin contents were detected using liquid chromatography-tandem mass spectrometry (LC-MS/MS). The present study aims to provide an insight into the diversity of benthicProrocentrumand to identify DSP toxins produced by these species in the Xisha Islands.
2 MATERIAL AND METHOD
2.1 Study region and sampling
The survey region was the Xisha Islands(15°46′N-17°08′N, 111°11′E-112°54′E), the largest archipelago in the South China Sea, and the islands are internationally called as Paracel Islands. This region has a tropical climate. The islands consist of Xuande Islands and Yongle Islands, with a total land area of 9.22 km2and a total marine area of 500 000 km2.Sampling was carried out at four sites on the Xisha Islands between 2015 and 2017 (Fig.1).
Samples of macroalgae, seagrasses, dead coral reefs, and sediments were collected by the diver in shallow water (~2-5-m depth) and transferred into bottles underwater. They were filtered using a 120-μm screen to remove sands and substrates, and the microalgae were collected on 20-μm mesh. The microalgae retained on the latter filters were resuspended for cell isolation. Single-cell ofProrocentrumwas isolated using a light microscope(CX31, Olympus, Japan) and cultivated in a 96-well cell culture cluster containing L1 medium without silicate (Guillard and Hargraves, 1993). Successfully established strains were transferred into glass tubes with L1 medium without silicate and used to research the biodiversity of benthicProrocentrumin the Xisha Islands. All cultures were maintained at temperature of 25 °C (±0.5 °C) and 30-31 in salinity, with a photoperiod of 12-h light∶12-h dark and irradiation of 120 μmol photons/(m2·s).
2.2 Morphological observation
To describe the morphologies of benthicProrocentrumfrom Xisha Islands, light, fluorescence,and scanning electron microscopy were carried out by the descriptions of Zou et al. (2020). In general, cells fixed were used to take pictures using a light microscope (BX61, Olympus, Japan) equipped with a digital camera (QImaging Retiga 4000R, British Columbia, Canada). The cell sizes of eachProrocentrumspecies were calculated based on the length and width of at least 30 cells. Live cells andcells stained with Fluorescent Brightener 28 (Sigma,USA) and SYBR Safe DNA gel stain (Invitrogen,Thermo Fisher Scientific, USA) were taken micrograph at ×1 000 magnification for the observations of shape and location of nucleus and chloroplasts.
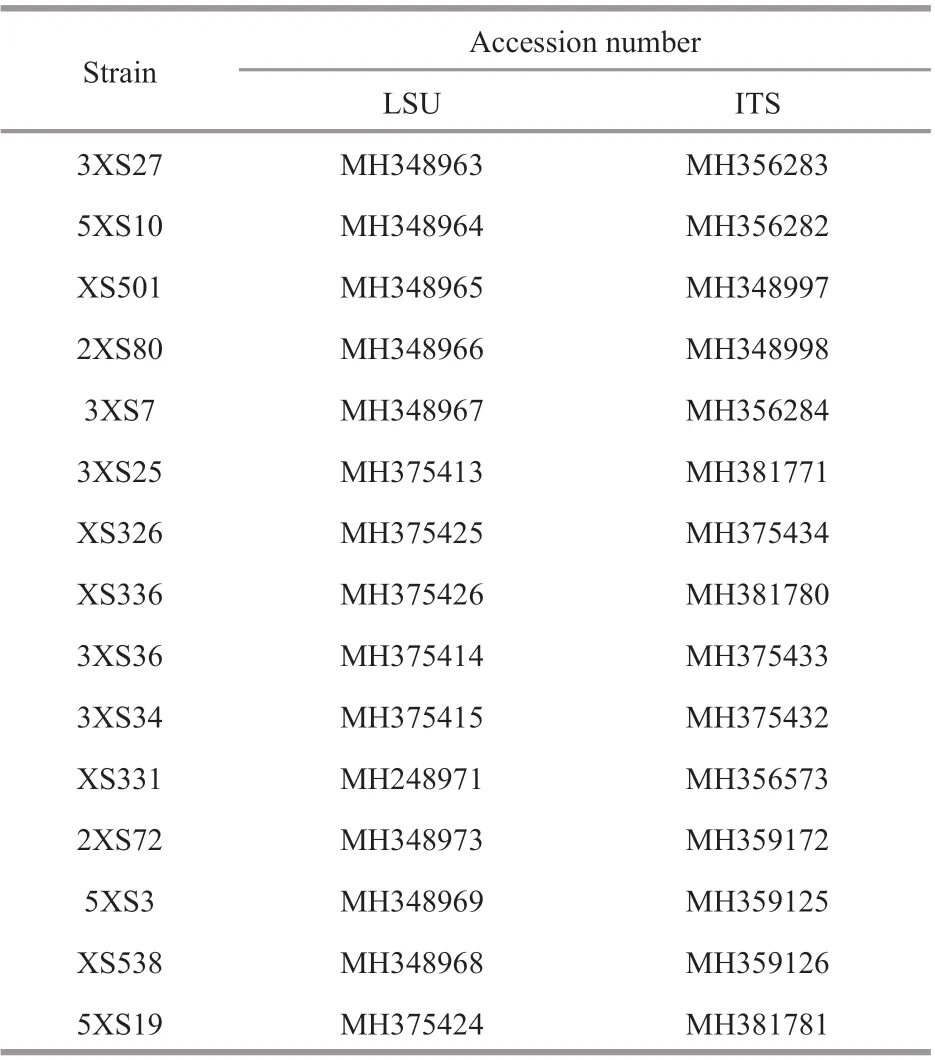
Table 1 The accession number of benthic Prorocentrum from Xisha Islands
A 5-mL well-grown algal culture was collected and rinsed thrice with filtered seawater and ultrasonically cleaned for 40 min. The cells were fixed in a 2.5% v/v solution of glutaraldehyde dissolved in filtered seawater and maintained at 4 °C overnight. The fixed microalgae were rinsed with Milli-Q water and dehydrated using a series of graded ethanol (10%,30%, 50%, 60%, 80%, and 95% v/v) for 5 min at each step and followed by 99.9% v/v twice for 15 min every time. Then, cells were dried to a critical point by supercritical drying (CPD 300, Leica, Wetzler,Germany) and coated with gold (Leica EM SCD 500,Leica, Wetzler, Germany). The prepared samples ofProrocentrumwere identified and photographed using a ZEISS ULTRATM55 scanning electron microscope. The platelet arrangements of the periflagellar areas ofProrocentrumwere identified using the descriptions of Hoppenrath et al. (2013).
2.3 DNA extraction, PCR amplification and sequencing
A 5-mL well-grown cells ofProrocentrumwere centrifuged at 13 000×gat 4 °C for 2 min and extracted using a MiniBEST Universal DNA Extraction Kit(TaKaRa, Tokyo, Japan) based on manufacturer’s instructions. The extracted total DNA was amplified using the primers D1R/D3B (Scholin et al., 1994) and ITS1F/ITS1R (Pin et al., 2001) for large ribosomal subunit (LSU) and ITS1-5.8S-ITS2 region,respectively. The 50-L reactions consisted of 22-μL sterile Milli-Q water, 1 μL of each primer with a concentration of 1 μmol/L, 1-μL total DNA (final concentration, 10-50 ng/μL), and 25-μL 2X Accurate Taq Master Mix (dye plus) (Accurate Biotechnology Co., Ltd.). The reactions were amplified based on previous methods of Zou et al. (2020). The products were sequenced using an ABI PRISM 3730XL(Applied Biosystems, CA, USA).
2.4 Sequence alignment and phylogenetic analyses
The sequenced products were jointed using pairwise alignment (ContigExpress V. 3) and the obtained new sequences were uploaded to the GenBank database (Table 1). The new sequences of partial LSU and ITS were aligned with those ofProrocentrumdownloaded from the GenBank database using multiple sequence alignment(MUSCLE, www.ebi.ac.uk/Tools/msa/muscle/).Adenoideseludens(Herdman) Balech andKarenabrevis(C. C. Davis) Gert Hansen & Ø.Moestrup were used as outgroups of LSU and ITS phylogeny,respectively. The tool RAxML-HPC2 on XSEDE v.8.2.12 (https://www.phylo.org/) was used to build the maximum-likelihood trees with default algorithm parameters and node support was run with 1 000 bootstrap repetitions. The best models GTR and SYM+G were selected using PAUP v 4.0 to analyze Bayesian Inference (BI) in LSU and ITS region,respectively. The BI trees were constructed with MrBayes V. 3.1.2 and four Markov chain Monte Claro chains calculated no less 10 000 000 replicates with sampling every 100 generations. The pairwise genetic distances were aligned using MUSCLE and calculated using MEGA-X (Kimura 2-parameter model).
2.5 Identification of toxin profile
A 0.5-L exponential algal culture (three triplicates)was harvested at 12 000×gat 4 °C for 10 min. Pellets were added with 3-mL methanol and ultrasonicated(3s∶3s) for 40 min at an ice bath. The extractions were centrifuged at 3 000×gat 4 °C for 15 min. The pellets were extracted with 3-mL methanol twice and all supernatants were put together. The 0.5-mL supernatants were filtered with 0.22-μm spin-filters(Pall Corporation, USA). Three toxins (OA, DTX-1,and DTX-2) were determined and quantified as follows. Chromatographic separation was carried out using an HPLC system (Shimadzu prominence LC-20ADXR) coupled with a tandem mass spectrometer(5500 QTRAP LC-MS/MS system, AB Sciex Instruments, Foster City, CA). Toxins were separated using Phenomenex Kinetex XB-C18 (150 mm×2.1 mm, 2.6 μm) column. The mobile phase consisted of acetonitrile (solvent A) and 0.15% formic acid in water (solvent B). The initial proportion of solvent A was 20%, increased to 90% within 7 min, kept constant for 3 min, and reduced to 20% in 0.1 min. The total run time for the analysis was 12 min. The injection volume was 5 μL and the flow rate was kept constant at 0.35 mL/min. The turbo ions-pray source settings were: Ions-pray voltage (negative polarity), -4 500 V;Curtain gas, 20 psi; GS1 and GS2, 50 psi; Ion Source Temperature, 550 °C; CAD, medium. Nitrogen served as nebulizer gas and collision gas in both modes.Multiple reaction monitoring with two products were selected: OA and DTX-2, 803.5>563.4 (55 eV) /255.2(61 eV); DTX-1, 817.6>563.4 (60 eV) /255.2 (65 eV).
3 RESULT
3.1 Morphological feature
3.1.1ProrocentrumborbonicumTen-Hage
Prorocentrumborbonicumcells are oval to ovoid and almost symmetric. The posterior area of cell is broadly rounded (Fig.2a & d). The cell length is 24.40-31.92 μm (average=26.18±3.43 μm,n=68)and its width is 18.45-26.69 μm (average=22.84±1.99 μm,n=68). Their length/width (L/W) ratios are 1.05-1.25. The pyrenoid is located in the center of the cells and is encompassed by numerous golden-brown chloroplasts (Fig.2b). The nucleus is small, ovoid and located in the posterior (Fig.2c). The thecal plates are ornamented by numerous depressions and dozens of scattered pores. The thecal surfaces of the plate have two types of large, sparse pores. The diameters vary between 0.12 and 0.2 μm in large pores(average=0.15±0.02 μm,n=30) and between 0.08 and 0.09 μm in small pores (average=0.08±0.03 μm,n=30). The center of cell is devoid of large pores but it is ornamented by scattered small pores and depressions (Fig.2e-f). The wide, V-shaped periflagellar area is located in the anterior areas of cells and consists of eight or nine platelets with unequal size and shape (1, 2, 3, 4, 5, 6a, 6b, 7, and 8).The periflagellar area has one relatively large flagellar pore and a small accessory pore (Fig.2h).
3.1.2Prorocentrumcaipirignum
Morphologies ofP.caipirignumcells are symmetrically oval to ovoid (Fig.3a & d). The length and width are 35.1-45.0 μm (average=40.57±2.44 μm,n=31) and 28.0-33.4 μm (average=31.59±1.98 μm,n=31), respectively, with L/W ratios of 1.13-1.41.The pyrenoid has starch ring and is located in the center of cell. Many chloroplasts radially arrange from the center to the margin (Fig.3b). The heartshaped nucleus is located in the posterior of cell(Fig.3c). The large lateral plates are ornamented with many kidney-shaped pores, but the center of cells is devoid of these structures (Fig.3e-f). The pores are 0.69-0.98-μm long (mean 0.84±0.09 μm,n=30) and 0.27-0.49-μm wide (mean 0.38±0.06 μm,n=30)(Fig.3g). The periflagellar area is V-shaped and platelet pattern is 1, 2, 3, 4a, 4b, 5 and 6 (Fig.3h-i).
3.1.3Prorocentrumconcavum
Broad oval cells ofP.concavumare symmetrical,and the cell length and width are 40.6-52.6 μm(average=48.1±2.7 μm,n=47) and 35.1-43.0 μm(average=39.9±2.0 μm,n=47), respectively (Fig.4a &d). Pyrenoid is located in the center of cells and chloroplasts position peripherally and radiate toward the cell center (Fig.4b). The cells have L/W ratios of 1.07-1.33 and have oval nucleus at the posterior(Fig.4c). The thecal cell surface possesses numerous round to oval reticulate-foveate depressions and scatters pores encircled by ring-like structures(Fig.4e-g). Thecal pores are 0.14-0.29 μm (mean 0.18±0.14 μm,n=26) and are loosely scattered on the cell surface except in central area (Fig.4e-g). Figure 4h-i shows the wide V-shaped periflagellar area and the arrangement of periflagellar platelets (1a, 1b, 2, 3,4, 5, 6, 7, and 8).
3.1.4Prorocentrumelegans
Prorocentrumeleganscells are oval and slightly asymmetric (Fig.5a & d). They are 18.76-22.04-μm long (average=20.31±0.9 μm,n=65) and 14.61-17.82-μm wide (average=15.84±1.14 μm,n=65),with L/W ratios of 1.17-1.49. There is no pyrenoid but numerous chloroplasts are observed (Fig.5b).The teardrop-shaped nucleus is located between the posterior and the center of cell (Fig.5c). The thecal view of each cell is flattened. Except the center, the theca is ornamented with pores in two different sizes. The diameters of larger pores are 0.25-0.37 μm (average=0.30±0.03 μm,n=33) and those of smaller pores are 0.07-0.12 μm (average=0.1±0.01 μm,n=41). The larger pores near the periflagellar area generally form radial rows. There are no pores in the center of each cell (Fig.5e-f). The wide,V-shaped periflagellar area is located in the cell apex and consist of seven or eight platelets (1, 2, 3, 4, 5,6, 7, and 8). Platelet 1 possesses a distinct wing and the flagellar pore larger than the accessory pore(Fig.5h-i).
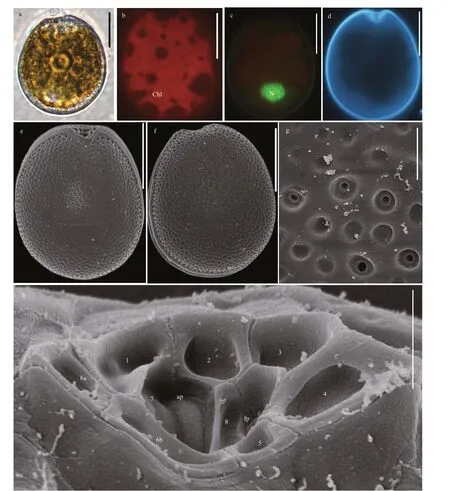
Fig.2 The morphological characters of Prorocentrum borbonicum based on light microscopy (LM) and scanning electron microscopy (SEM)
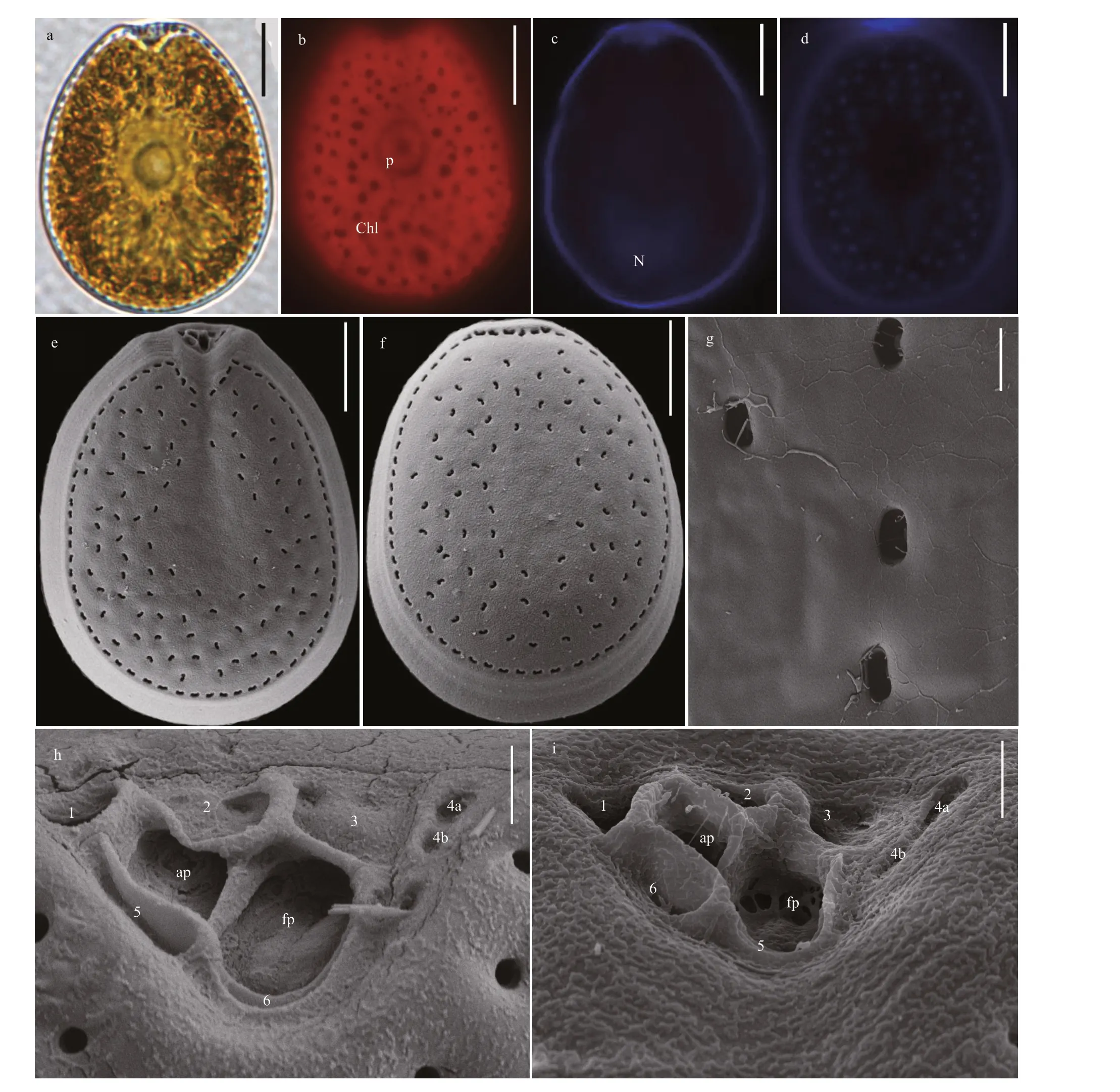
Fig.3 The morphological characters of Prorocentrum caipirignum based on light microscopy (LM) and scanning electron microscopy (SEM)
3.1.5Prorocentrumcf.emarginatum
Round to ovalP. cf.emarginatumcells are asymmetric and 29.95-37.65-μm long (average=33.98±2.04,n=32) and 23.22-33.76-μm wide(average=28.89±3.11 μm,n=32), with L/W ratios of 1.06-1.50 (Fig.6a & d). The elongated nucleus and pyrenoid encircled by numerous chloroplasts from center to periphery are located in the posterior and in the center area of the cell, respectively (Fig.6b-c). The thecal surfaces are foveate and ornamented with pores of two different sizes. The mean diameter of the larger pores is 0.18 μm and that of the smaller pores is 0.1 μm. The centre of each cell lacks pores (Fig.6e-f).Pores of both sizes are located in deep depressions and arrange in stripes (Fig.6g). The anterior narrow V-shaped periflagellar area is composed of nine platelets (1, 2, 3, 4, 5, 6a, 6b, 7, and 8). A wing-shaped spine is observed between platelet 1 and platelet 2 in each cell (Fig.6h & i).
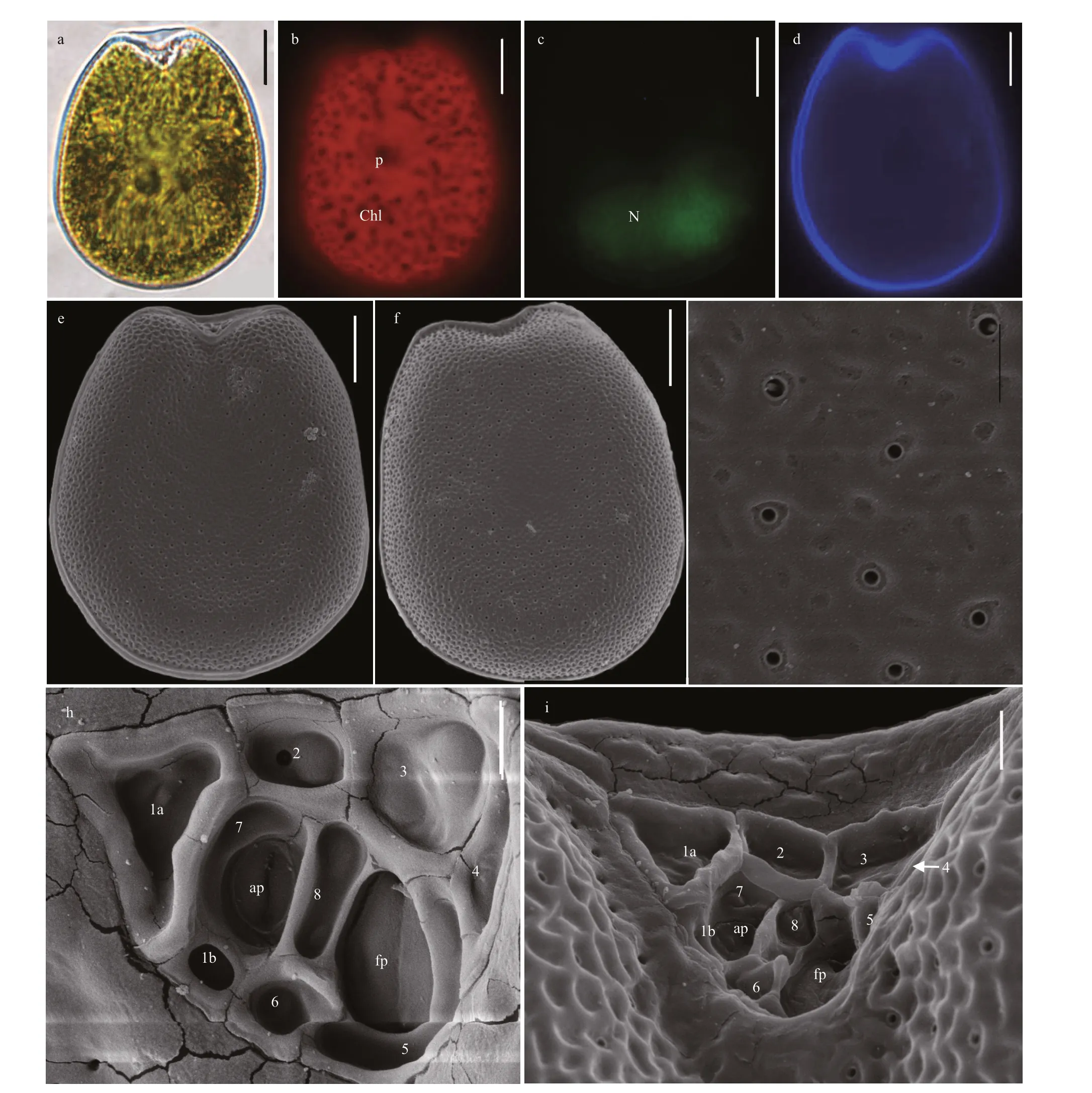
Fig.4 The morphological characters of Prorocentrum concavum based on light microscopy (LM) and scanning electron microscopy (SEM)
3.1.6Prorocentrumlimacomplex
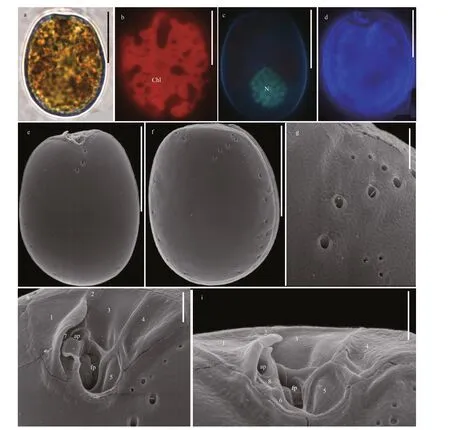
Fig.5 The morphological characters of Prorocentrum elegans based on light microscopy (LM) and scanning electron microscopy (SEM)
Prorocentrumlimacomplex cells are oval to oblong-oval and symmetric (Fig.7a & d). The length,width, and ratios of L/W are 34.8-40.7 μm (average=38.09±1.59 μm,n=33), 22.9-27.4 μm (average 25.47±1.26,n=33), and 1.36-1.61, respectively.Pyrenoid is located in the center of cell and is surrounded by numerous chloroplasts (Fig.7b). The oval nucleus is located in the posterior area of cell(Fig.7c). The thecal plates are smooth and ornamented by many scattered and oblong to kidney-shaped pores with 0.25-0.50 μm in length and 0.17-0.39 μm in width. The central area of cell is devoid of pores. The marginal pores arrange in ring (Fig.7e-g). Seven platelets present in a wide, V-shaped periflagellar area(1, 2, 3, 4, 5, 6, and 7). A flagellar pore is much larger than an accessory pore (Fig.7h-i).
3.1.7Prorocentrumrhathymum
The shapes ofP.rhathymumcell are asymmetrically oval, and sizes are 27.5-34.7-μm long(average=32.8±1.5 μm,n=32) and 20.7-25.5-μm wide (average=23.6±1.14 μm,n=32), with L/W ratios of 1.2-1.56 (Fig.8a & d). The cell lacks pyrenoid but has numerous chloroplasts (Fig.8b).The posterior of cell possesses kidney-shaped nucleus (Fig.8c). The large lateral plates are ornamented with pores in two size. The diameters of larger pores are 0.41-0.71 μm (average=0.57±0.09 μm,n=33) in the depressions, and the smaller pores are only 0.07-0.24 μm (average=0.15±0.07,n=33). The pores radiate from the smooth center to the margin (Fig.8e & f). The wide V-shaped periflagellar area is composed of nine platelets (1, 2,3, 4, 5, 6a, 6b, 7, and 8) and has an ear-shaped wing supported by platelet 1 (Fig.8h-i).
3.2 Molecular phylogeny
The maximum likelihood and Bayesian inference differed only in a few topologies based on LSU region, and they also had a few topological differences in ITS analysis. Phylogenies inferred from LSU rDNA and ITS region are illustrated by ML trees(Fig.9 & Supplementary Fig.S1).
In the ML tree inferred from LSU rDNA (Fig.9),Adenoideseludenswas an outgroup. Considering the whole tree, the phylogenies ofProrocentrumdivided into two large clades. The first clade, including clade A, clade B, clade C,P. cf.norrisianum, andP.glenanicum, mainly consisted of benthicProrocentrum. The second clade, including clade D and clade E, was composed of planktonic and epibenthic species (Fig.9). In the first clade, clade A consisted ofP.hoffmannianum,P.caipirignum,P.limacomplex,P.consutumChomérat & Nézan,andP.bimaculatumChomérat & Saburova with a high support value (95/1.00). The new obtained strains ofP.limacomplex from Xisha Islands belonged toP.limacomplex, with a support value of 84/1.00. Moreover,P.limacomplex divided into two subclade and four strains from Xisha Islands were located in a same clade (100/1.00).P.caipirignum,which consisted of XS331 and other strains, andP.hoffmannianumwere the sister clade ofP.limacomplex, with the highest support value (100/1.00).Besides, the newly characterized strain 2XS72 and other sequences ofP.concavumdownloaded from GenBank formed a clade with a full support value(100/1.00). The clade A and clade B were a sister clade with clade C, which consisted ofP.playfairi,P.sipadanenseMohammad-Noor, Daugbjerg et Moestrup, andP.borbonicum.P.borbonicumconsisted of Xisha strains (XS538, 5XS3, 5XS19)and other strains showed the highest support(100/1.00). In the clade mainly composed of planktonic species, clade D consisted ofP.fukuyoicomplex,P. cf.sculptile, andP. cf.emarginatum,with a high support value (98/1.00). The new characterized strains and two previous strains were composed ofP. cf.emarginatumclade (100/1.00). In addition, a clade E, comprising Xisha strains of two benthic species (P.elegansandP.rhathymum) and other strains of planktonic species, includingP.dentatumStein,P.minimum(Pavillard) J. Schiller,P.triestinumJ. Schiller,P.gracileF. Schütt,P.micans, andP.koreanumM.-S. Han, S. Y. Cho &P. Wang in M.-S. Hana, was sister of clade D. Among them,P.eleganswas basal toP.dentatumStein and divided into three subclades. In addition, the new obtained strains (X501 and 2XS80) belonged toP.rhathymum, which consisted of two subclades,with a support value of 93/1.00. In the analysis using ITS sequences,Karenabreviswas an outgroup. In general, the tree topologies were similar to the phylogenies based on LSU sequences and exhibited analogous results (Supplementary Fig.S1).
3.3 Detection of toxins
The contents of OA, DTX-1, and DTX-2 in all fifteen strains (oneP.concavumstrain, oneP.caipirignumstrain, twoP.rhathymumstrains,threeP.borbonicumstrains, twoP.elegansstrains,twoP. cf.emarginatumstrain, and fourP.limacomplex strains; Table 2) from Xisha Islands were analyzed using LC-MS/MS. All strains ofP.limaproduced OA, with 1 663- 3 816 fg/cell (Table 2).TwoP.limacomplex strains (XS336 and XS326)from Yongxing Island generated DTX-1 ranging from 74 to 183 fg/cell (Table 2). In addition,P.caipirignumstrain (XS331) from Yongxing Island produced OA(407 fg/cell) but no DTXs. Other five species,includingP.rhathymum,P.concavum,P.borbonicum,P.elegans, andP. cf.emarginatum, from Xisha Islands had no detectable toxins (Table 2).
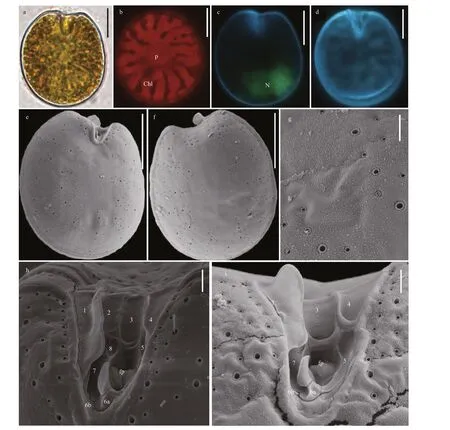
Fig.6 The morphological characters of Prorocentrum cf. emarginatum based on light microscopy (LM) and scanning electron microscopy (SEM)
Interestingly, we found a suspected DTX-2 in twoP.limastrains (3XS34 and 3XS36). The peak area ratios between different transitions of OA and suspected DTX-2 in samples were compared with the standard OA and DTX-2 (Supplementary Table S1),which demonstrated that the qualitative of OA could be satisfactorily supported by these results, but the suspected DTX-2 peak could not be testified completely because there was about 10% variation between samples and standard (Fig.10). The mass spectrums of standard DTX-2 and the suspected DTX-2 in samples at collision energy 80 EV showed that the fragment ions of pseudo-molecular ions of both standard DTX-2 and the suspected compounds were basically same and their relative abundance were also similar (Fig.10). However, the retention time of OA did not differ more than 0.04 min betweenstandard and samples, and this difference in retention time reached up to 0.08-0.1 min between the DTX-2 standard and the corresponding peak in both samples.Therefore, we considered this suspected DTX-2 was an isomer of OA and DTX-2.
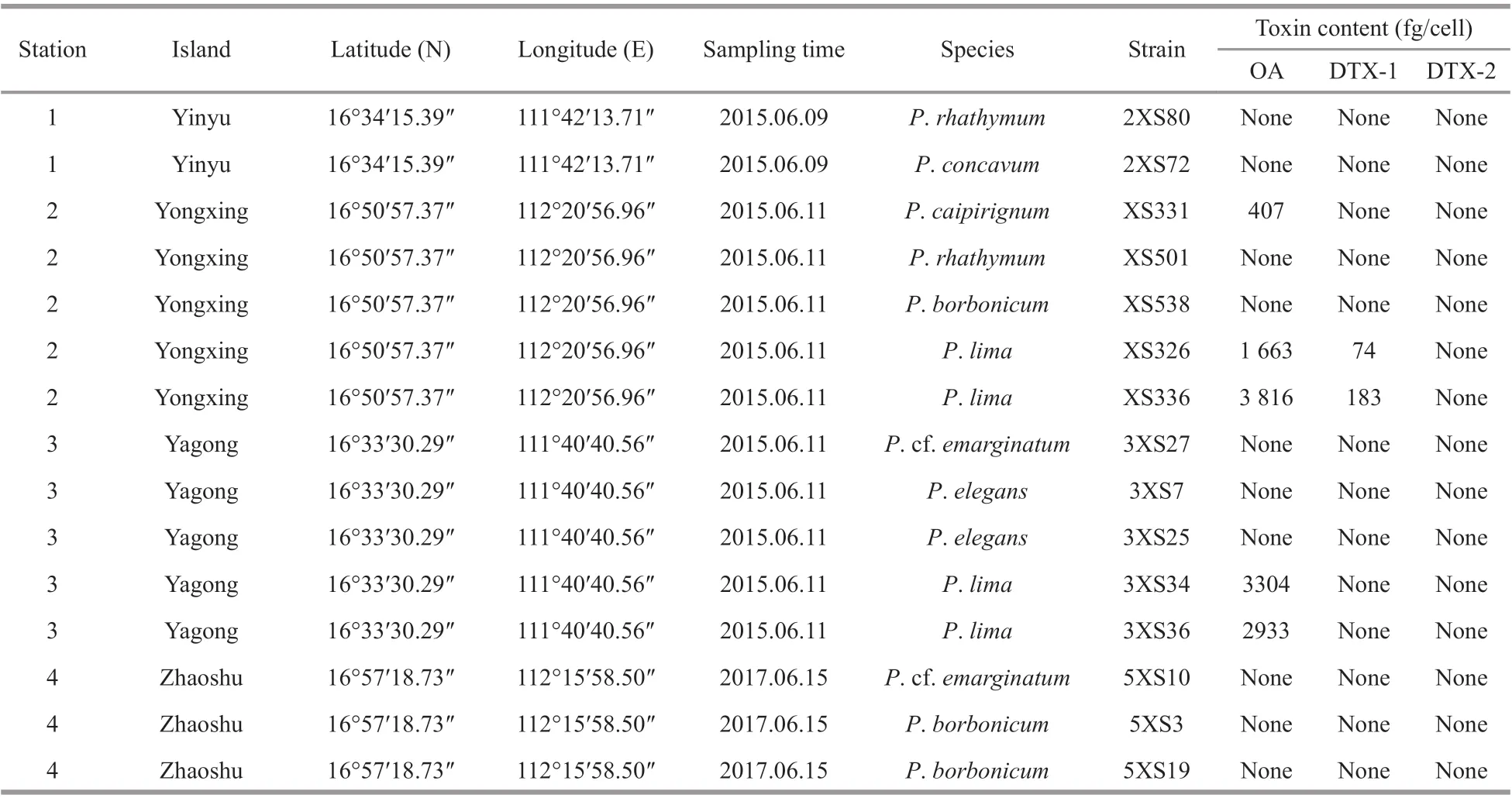
Table 2 Benthic Prorocentrum observed in this study
4 DISCUSSION
4.1 Taxonomic characteristics of benthic Prorocentrum from Xisha Islands
4.1.1ProrocentrumborbonicumandProrocentrumelegans
ProrocentrumborbonicumandP.eleganswere first reported in Chinese water, allowing us to compare the morphological and phylogenetic characteristics of Xisha Islands strains with other previous strains.Except the periflagellar area, the morphologies ofP.borbonicumfrom Xisha Islands are consistent with the original descriptions (Ten-Hage et al., 2000a).They described the species with only eight platelets,but the periflagellar area of Xisha Islands strains consisted of nine platelets as platelet 6 divided into 6a and 6b (Fig.2h). Our findings are in agreement with the descriptions by Hoppenrath et al. (2013), who showed the periflagellar area ofP.borbonicumconsisted of eight or nine platelets. Besides,P.borbonicumcould morphologically differ from the similar speciesP.sipadanense. The latter is smaller(17.9-23.9-μm long and 15.0-19.8-μm wide) and devoid of pores in the cell center, butP.borbonicumfrom Xisha Islands was larger (24.40-31.92-μm long and 18.45-26.69-μm wide) and scattered small pores in the center of cell (Fig.2). Also, they could differ from the characteristics of phylogeny (Fig.9).P.borbonicum, comprising Xisha Islands strains and other sequences from GenBank with highest support value, was basal toP.sipadanensefrom France andP.playfairiCroome from Australian freshwater in ML tree inferred from LSU rDNA, with a high support value (99/1.00, Fig.9). In addition,P.borbonicumwas less reported to date and was found in France, the southeast Indian Ocean (Ten-Hage et al., 2000a),Greece, Mediterranean (Aligizaki et al., 2009), and Xisha Island, China (present study).
The morphologies ofP.elegansfrom Xisha Islands were consistent with the original descriptions from Twins Cay, Belize (Faust, 1993). This species was characterized by a large, wide V-shaped periflagellar area, a protuberant spine at platelet 1 and 2 different sizes of pores (David et al., 2014). They isolatedP.elegansstrain Dn208EHU and firstly obtained molecular sequences with high support values, which revealed that this species obviously differed from otherProrocentrumspecies (David et al., 2014). In the present study, the phylogenies showed thatP.elegans,comprising two Xisha Islands stains and two strains from Canary Islands and France, divided into three subclades, which indicates this species has high genetic diversity. To date,P.eleganswas only reported in Biscay, French (David et al., 2014), Twin Cays, Belize(Faust, 1993), and Chinese waters (present study).
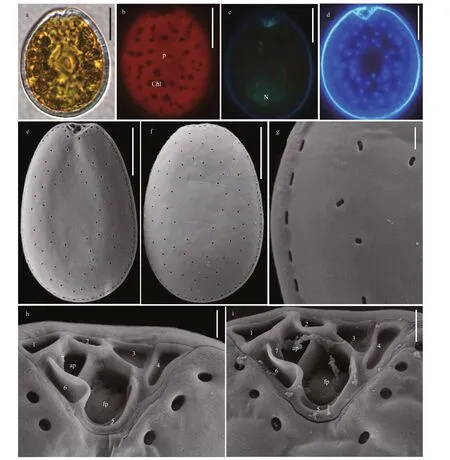
Fig.7 The morphological characters of Prorocentrum lima complex based on light microscopy (LM) and scanning electron microscopy (SEM)
4.1.2Prorocentrumlimacomplex andProrocentrumcaipirignum
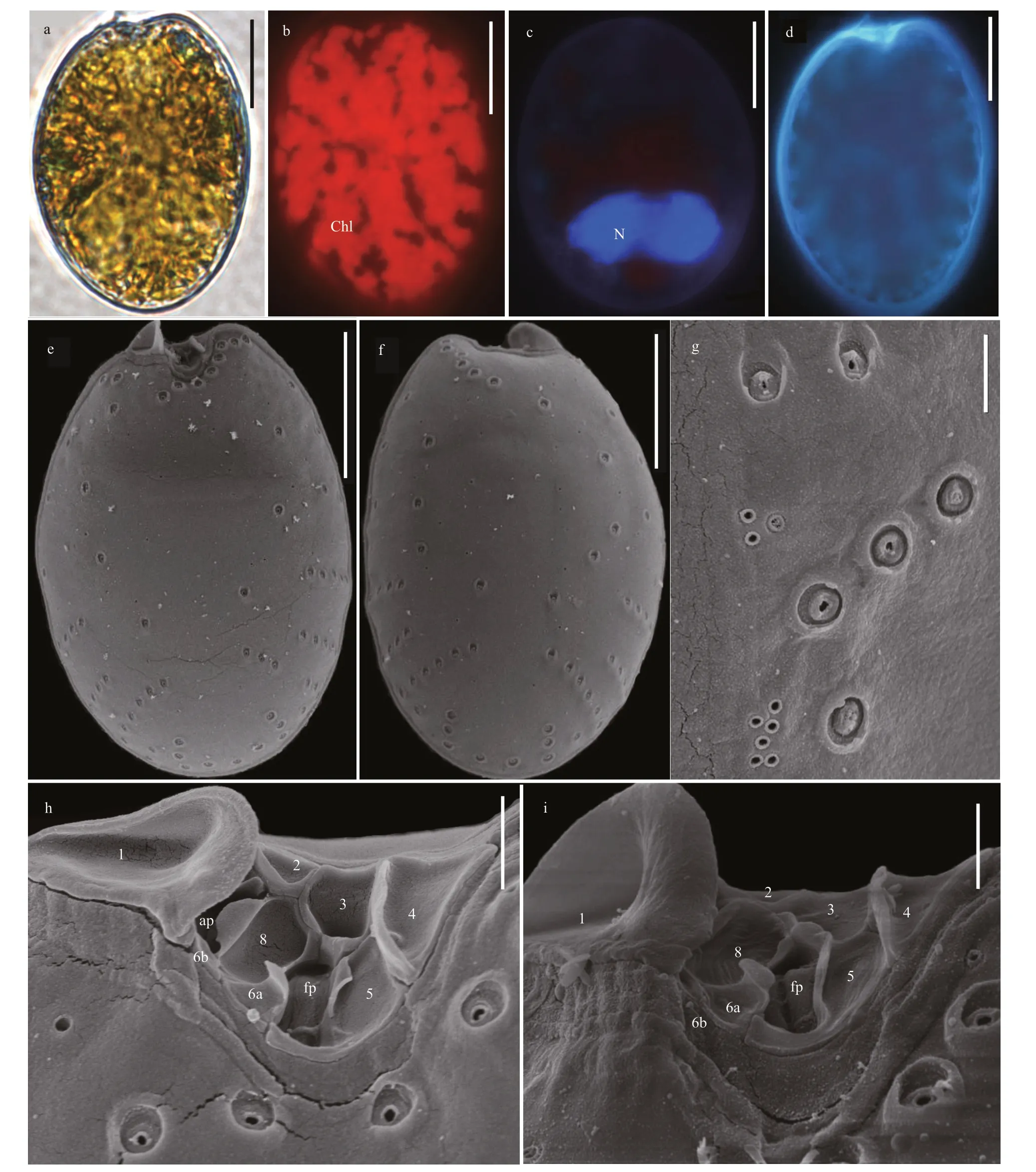
Fig.8 The morphological characters of Prorocentrum rhathymum based on light microscopy (LM) and scanning electron microscopy (SEM)
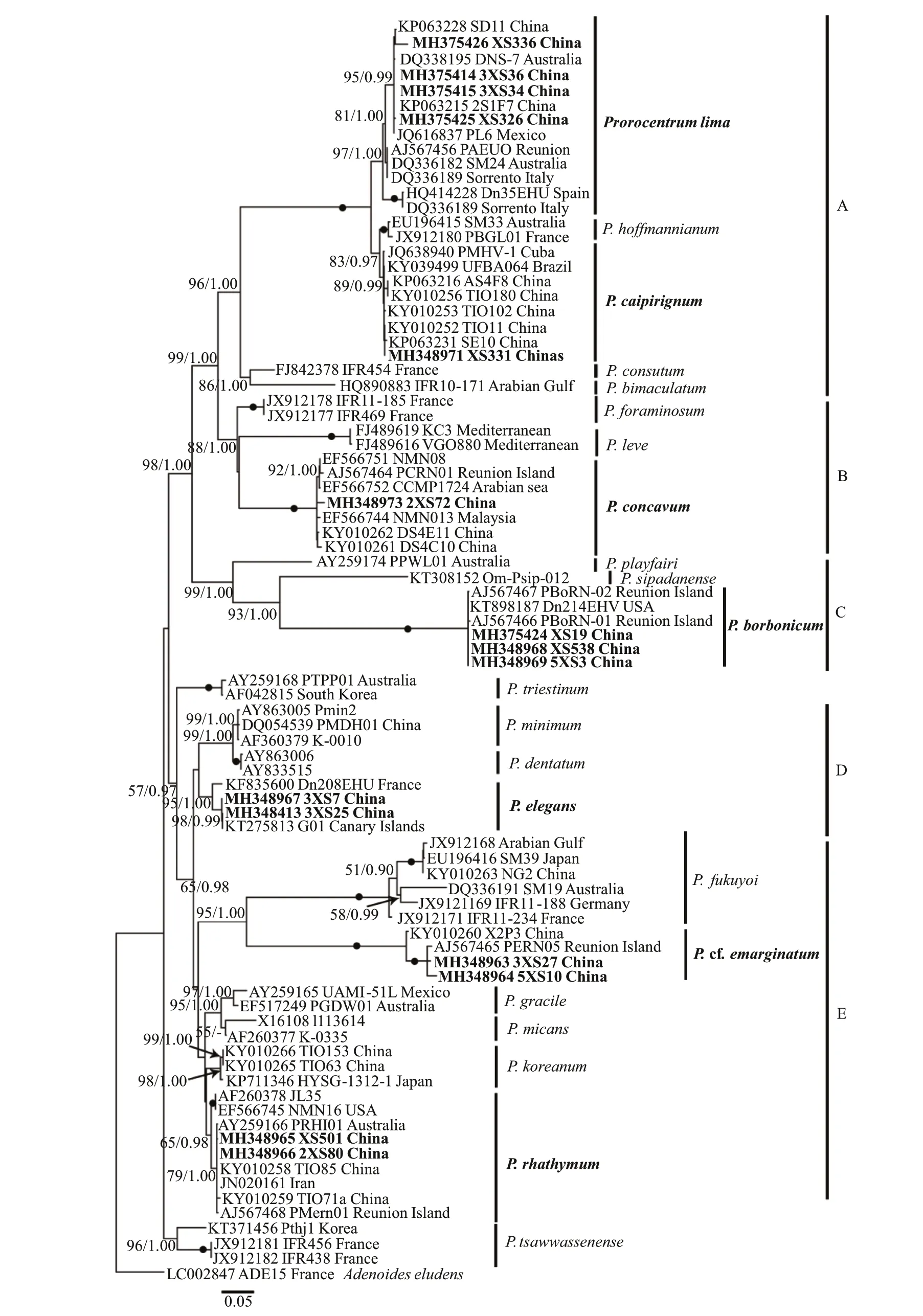
Fig.9 Molecular phylogenetic tree (maximum likelihood) of Prorocentrum inferred from D1-D3 LSU rDNA
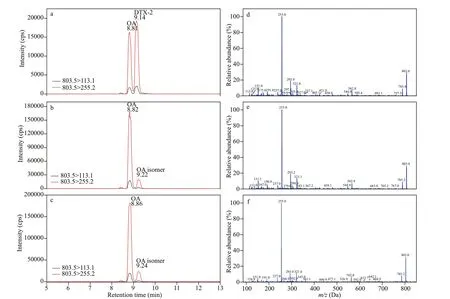
Fig.10 Multiple reaction monitoring (MRM) chromatograms of: OA and DTX-2 reference standards (a), and OA and putative DTX-2 in strain 3XS34 and 3XS36 extracts (b and c, respectively); mass spectrum of DTX-2 in standard (d),and putative OA isomer in 3XS34 (e) and 3XS36 (f) with a collision energy of 80 eV
We found DSP toxins were detected only inP.limacomplex andP.caipirignum. A study suggested thatP.limacomplex could divide into five morphotypes collected from Hainan Island closed to our study area(Zhang et al., 2015), whileP.caipirignumwas erected by morphotypes 4 ofP.limacomplex based on the descriptions of Nascimento et al. (2017). The Xisha Islands strains ofP.limawere basically in accord with original descriptions of specimens collected in French Polynesia, New Caledonia and Ryukyu Islands(Fukuyo, 1981). The exception was that Platelets 8 of periflagellar area in Xisha Islands strains was absent and platelet 7 was not obvious (Fig.7h-i). Besides,P.limacomplex was morphologically analogous withP.caipirignum. They could only be morphologically distinguished by their cell shape and thecal plate ornamentations (pore shape and size; absence or presence of a flange) (Nascimento et al., 2017). The distinct differences betweenP.caipirignumandP.limacomplex in morphology presented in the present study. For instance,P.caipirignumcells are oval (L/W ratios: 1.13-1.41) whileP.limacomplex is ovoid (L/W ratios: 1.36-1.61). The marginal pores ofP.limacomplex were elongated, butP.caipirignumhas kidney-shaped marginal pores, which twice to thrice larger than the former one (Nascimento et al.,2017). Moreover, the periflagellar area (1, 2, 3, 4a, 4b,5, and 6) ofP.caipirignumwere consistent with the original descriptions that platelet 2 extended the right lateral plate and separated the flagellar pore and accessory pore (Nascimento et al., 2017). Interestingly,the platelet 4 potentially divided into 4a and 4b in Xisha Islands stains (Fig.3h-i), which differed from the original descriptions (Nascimento et al., 2017). It is necessary to further clarify the morphological characteristics inP.caipirignumin the future.
Based on the ML trees inferred from LSU and ITS rDNA, the two toxic species of benthicProrocentrumfrom the Xisha Islands could differ from each other(Fig.9 & Supplementary Fig.S1). Many surveys revealP.limacomplex has high genetic diversity and may exist cryptic species. Aligizaki et al. (2009)statedP.limahad a high genetic diversity and replaced the term by ‘P.limacomplex’. Recently, a study in Lesser Antilles, eastern Caribbean Sea revealed thatP.limacomplex divided into five subclades and genetically related toP.caipirignum,P.consutum,P.bimaculatum, andP.hoffmannianum(Chomérat et al., 2019). Nishimura et al. (2020a)collected 244 strains of benthicProrocentrumfrom Japan and showedP.limacomplex had four species complex/ phylotypes. In the present study, the ML tree inferred from LSU rDNA showedP.limacomplex consisted of two clades and four stains from Xisha Islands were located in a same clade (Fig.9),which was the clade 1a by Nishimura et al. (2020a).However, the ML tree based on ITS region showed four strains from Xisha Islands divided into two clades (Supplementary Fig.S1). The strains (3XS34 and 3XS36) from Yongxing Island formed a clade,and Yagong Island strains (XS336 and XS334) were located in the other clade (Supplementary Fig.S1 &Table 2). Our findings confirmP.limacomplex from Xisha Islands also is a complex and genetically related to clade 1a from Japan (Nishimura et al., 2020a). In addition, these findings coupled with the differences in toxin production provide a new proof for Nishimura et al. (2020a) stated there might be the existence of relationship between phylogenetic features and DSP toxins productions (Table 2).
4.1.3Prorocentrumconcavum,Prorocentrumcf.emarginatum, andProrocentrumrhathymum
ProrocentrumconcavumandP.rhathymumfrom Xisha Islands could differ from other benthicProrocentrumspecies in both morphology and phylogeny, and the taxonomic characteristics of them were consistent with the original descriptions and recent findings (Fukuyo, 1981; Gómez et al., 2017;Zou et al., 2020). From a morphological view,P. cf.emarginatumdiffered fromP.fukuyoicomplex in cell size and thecal ornamentation. The former was 29.95-37.65-μm long and 23.22- 33.76-μm wide (L/W, 1.06-1.50) with foveate thecae, andP.fukuyoiwas 26.2-37.9-μm long and 18.0-26.5-μm wide from Beihai,Guangxi, with a smooth ornamentation (Luo et al.,2017). However,P. cf.emarginatumfrom Xisha Islands was similar toP. cf.sculptilefrom eastern Caribbean in morphology (Chomérat et al., 2019).From a molecular point, the ML tree inferred from LSU region showedP. cf.emarginatumfrom Xisha Islands was well divergent fromP. cf.sculptilefrom eastern Caribbean, with a high support (98/1.00, Fig.9).As suggested by Chomérat et al. (2019), the genetic distance between Xisha Islands strains and strains of Martinique were calculated based on LSU region and was 0.26. Considering morphology and phylogeny, we described this species asP. cf.emarginatum.
4.2 Toxin profile
In our research, seven benthicProrocentrumspecies were collected from Xisha islands and only two (P.caipirignumandP.limacomplex) produced DSP toxins. There were contradictory conclusions whetherP.concavumproduced OA and/or DTXs.Previous studies foundP.concavumcollected from the Caribbean Sea could produce OA using highperformance liquid chromatography and mouse assay(Dickey et al., 1990; Juranovic et al., 1997). However,no OA or DTXs were detected inP.concavumisolated from regions such as Okinawa, Japan, and Hainan,China (Yasumoto et al., 1987; Luo et al., 2017).Although no DSP toxins were detected, strains ofP.concavumfrom Okinawa, Japan (Yasumoto et al.,1987) and Hainan Island, China (Zou et al., 2020) was potently toxic to fish, mice, and invertebrates,respectively. Also, no DSP toxins were detected inP.concavumstrains from Xisha Islands (present study). The production of DSP toxins inP.concavummight show a strain-specific variation. Caillaud et al.(2010) stated that LC-MS/MS revealed low quantities of OA inP.rhathymumfrom Malaysia. In addition,An et al. (2010) found that one of five strains ofP.rhathymumfrom Florida Bay produced OA.However, our results showed that no detectable OA was found inP.rhathymumfrom Xisha Islands,which is similar to previous findings (Yasumoto et al.,1987; Aligizaki et al., 2009). Recently, Lim et al.(2019) and Luo et al. (2017) reported all strains ofP.mexicanum(was synonymized withP.rhathymumby Gómez et al. (2017)) andP.rhathymumproduced no detectable DSP toxins, respectively. The findings that only one of six strains ofP.rhathymumcollected from Florida Bay produced OA (0.153 μg/L in the culture medium) (An et al., 2010). It suggested strainspecific variations also existed in this species.Previous studies showed thatP.borbonicumproduced borbotoxin (Ten-Hage et al., 2002) and its methanolic extract was lethal to mice and Artemia shrimp (Ten-Hage et al., 2000a, 2002; Aligizaki et al., 2009).However, this species did not generate any measurable OA, DTX-1, or DTX-2 (Aligizaki et al., 2009), which was consistent with our results of LC-MS/MS analysis. These results also were corroborated by the descriptions of Aligizaki et al. (2009) who used the protein phosphatase type 2A inhibition assay for their analyses. In addition, we foundP. cf.emarginatumis a nontoxic species, which was suggested by previous reports (Escoffier et al., 2007; Aligizaki et al., 2009).We also determined thatP.elegansdid not produce OA, DTX-1, or DTX-2, which was analogous to the results of David et al. (2014). In the present work,only OA was detected in the Xisha Islands strain ofP.caipirignum. This observation was supported by the results of the Belize study (Nascimento et al.,2017). The OA content of Xisha IslandsP.caipirignumwas 407 fg/cell, which was much lower than that reported in the Belizean strain (20 000 fg/cell)(Nascimento et al., 2017). The difference of OA content between strains of Xisha Islands and Belizean strain might be caused by culture conditions(temperature and light intensity) or strain-specific variations (Aquino-Cruz et al., 2018). In this study, allP.limacomplex strains collected from Xisha Islands contained OA and DTX. It demonstrated thatP.limacomplex was a toxic species worldwide (Bouaı¨cha et al., 2001; Foden et al., 2005; Vale et al., 2009; Luo et al., 2017; Pan et al., 2017). Notably, the types of DSP toxins generated byP.limacomplex showed strainspecific. BothP.limacomplex strains (XS326 and XS336) from Yongxing Island produced OA and DTX-1. The strains of Mexico, the UK, Spain, and Portugal also contained both toxins in different concentrations (Heredia-Tapia et al., 2002; Foden et al., 2005; Vale et al., 2009). However,P.limacomplex strains 3XS34 and 3XS36 from Yagong Island produced an isomer of OA and DTX-2 along with OA but no DTX-1 was detected. Our findings supported the concept that geographically distinct strains ofP.limacomplex not only can produce DSP toxins at different concentrations, but also they can generate various kinds of toxins.
5 CONCLUSION
In the present study, we demonstrated seven species of benthicProrocentrum, includingP.borbonicum,P.caipirignum,P.concavum,P.elegans,P. cf.emarginatum,P.limacomplex, andP.rhathymumusing morphology and phylogenetic analysis in Xisha Islands, South China Sea. All of them inhabited benthic environments. Among them,P.elegansandP.borbonicumwere the first records in Chinese waters. According to the results of LC-MS/MS analysis, no DSP toxins were produced byP.concavum,P.rhathymum,P.elegans,P. cf.emarginatum,P.borbonicum. But the other two species (P.caipirignumandP.limacomplex) could produce OA.P.limacomplex could generate DTX-1 or an isomer of OA and DTX-2. Our results demonstrated the risk of diarrheic shellfish poisoning in this area and it should be paid attention to.
6 DATA AVAILABILITY STATEMENT
The datasets generated and/or analyzed during the current study are available from the corresponding author on reasonable request.
7 ACKNOWLEDGMENT
We sincerely appreciate the three anonymous reviewers’ comments and suggestions, which are helpful for the manuscript. Prof. Yubo LIANG from the National Marine Environmental Monitoring Center is deeply appreciated for providing data about toxins. We also thank Prof. Aifeng LI for providing valuable suggestions for the paper revision.
 Journal of Oceanology and Limnology2022年3期
Journal of Oceanology and Limnology2022年3期
- Journal of Oceanology and Limnology的其它文章
- Typhoon-induced wind waves in the northern East China Sea during two typhoon events: the impact of wind field and wave-current interaction*
- Effect of subsea dispersant application on deepwater oil spill in the South China Sea*
- Geochemical characteristics of cold-seep carbonates in Shenhu area, South China Sea*
- Examination of seasonal variation of the equatorial undercurrent termination in the Eastern Pacific diagnosed by ECCO2*
- Deviation of the Lagrangian particle tracing method in the evaluation of the Southern Hemisphere annual subduction rate*
- Immunostimulatory effect of quaternary degree and acetyl group of quaternized chitosan on macrophages RAW 264.7*