
砧木对薄壳山核桃嫁接苗生长及叶片矿质元素含量的影响
2022-06-16 06:17:40汤玉洁李晓锐刘俊萍刘壮壮彭方仁
西南林业大学学报 2022年3期
汤玉洁 李晓锐 刘俊萍 刘壮壮 彭方仁
(1.南京林业大学林学院,南方现代林业协同创新中心,江苏 南京 210037;2.江苏省中国科学院植物研究所,江苏 南京 210014)
嫁接作为植物栽培中常用的一种无性繁殖手段,除了具有促进苗木提早开花结果、增强林木综合抗性、合理矮化增加果实产量[1]等优势外,还能提高嫁接苗的养分利用效率[2]。砧木作为嫁接苗的地下部分,承担着对于嫁接苗植株水分和矿物质的供应,因而对营养元素的吸收、运转及利用存在影响效应[3]。研究表明不同砧木的嫁接组合不仅在生长上不同,对养分含量的反应也有所差异,在柑橘[4](Citrus reticulata)、苹果[5](Malus domestica)、黄瓜[6](Cucumis sativus)等植物中均有相关论述,Fazio 等[7]指出砧木会诱导某一元素含量变化从而影响到其他元素含量,所以砧木可以通过影响嫁接苗营养元素的含量,进而调节植物生长发育。其中叶片作为光合作用和养分储存的场所,在碳水化合物的积累与养分运输过程中占据重要地位[8],其元素含量变化能够反映嫁接苗对于养分的分配情况和利用能力,与嫁接苗的生长质量和产量密切相关。氮、磷、钾是植物生长所需的基本营养元素,在有机物合成、能量代谢及提高抗逆性等方面具有重要作用[9],是植物高效生长和优产稳产的保证,其含量高低可以作为判断植物养分利用效率及生长限制元素的依据[10]。碳、氮代谢作为植物生长过程中2 种重要的生理代谢途径,其化学计量比在一定程度上反映了植物体内碳氮营养平衡与叶片的光合固碳能力[11],也是影响花芽分化和成花成果的重要因素。因此,研究叶片中氮、磷、钾及碳氮比含量对于了解植物生长养分需求、元素相互作用规律、提高果实产量及环境适应能力等具有重要意义。
薄壳山核桃(Carya illinoinensis)是胡桃科(Juglandaceae)山核桃属(Carya)一种集果用、油用、材用及绿化价值于一体的干果经济林树种,综合利用效益较高[12]。近年来,由于广泛的资源种植和优良的市场前景,薄壳山核桃在经济林中的产业比重越来越高,嫁接苗木的需求量稳步上升。目前,薄壳山核桃规模化产业种植以方块芽接为主要的嫁接技术手段,其操作体系成熟且简单易行,砧穗愈合生长快速,嫁接成活率高达97.35%[13]。在此基础上的砧木研究多集中于嫁接苗的光合特性、抗逆性等[14−15]差异比较,对于叶片矿质元素的分析较少,缺乏对于嫁接后苗木养分含量变化的了解,因此关于不同砧木的薄壳山核桃嫁接效应研究并不完善。
本研究选择薄壳山核桃'Pawnee'、'Stuart'、'Mahan'和'Jinhua' 4 个品种的实生苗作为砧木,利用方块芽接嫁接相同'Pawnee'接穗,通过比较4 种砧穗组合嫁接苗生长特性及叶片主要矿质元素含量的差异,分析不同砧木对于薄壳山核桃嫁接苗生长以及养分变化的影响,进一步探究养分含量与嫁接苗生长势之间的关系,以期为薄壳山核桃的砧木选择及科学施肥提供依据。
1 材料与方法
1.1 试验地概况
试验地选择位于江苏省句容市后白镇张庙村的南京林业大学薄壳山核桃试验基地(119° 90′50″E,31° 52′47″N)。该试验地地处低山丘陵区域带,夏季高温多雨,冬季低温少雨,四季变化明显,是典型的亚热带季风气候区域。年平均气温为15.6 ℃,活动积温为4 840 ℃,年均降水量可达1 018.6 mm,无霜期至229 d,土壤类型为中性偏酸的黄棕壤土[16]。
1.2 试验设计与材料
供试砧木材料为'Pawnee'、'Stuart'、'Mahan'及'Jinhua'种子培育的实生苗木,接穗均为'Pawnee'枝条。具体培育过程如下:于2017年11 月在上述4 个品种的母树上采集健康种子,室内通风3~4 d 后进行沙藏处理,2018 年1 月下旬经过赤霉素(100 mg/L)浸泡3 d 后播种,4 月中旬将幼苗移栽至容器中,以相同方式进行管理,8 月在每品种实生苗中选择长势一致(地径约1 cm)的苗木作为砧木,并采集同一'Pawnee'母树上当年生的健康枝条作为接穗进行方块芽接。
试验采用完全随机区组设计,4 种砧穗组合('Pawnee/Pawnee'、'Pawnee/Stuart'、'Pawnee/Mahan'、'Pawnee/Jinhua')设置3 次重复,每个重复9株苗木,共计108 株嫁接苗。2019 年4 月初对所有嫁接苗进行萌芽及展叶观察,统计嫁接成活率。于2019年6月16日和9月15日,在每组重复中随机选取3 株成活嫁接苗,进行叶片采集:叶片选择苗木中上部东、南、西、北方向生长健康的完整复叶,采集从顶端至下方的第4 对小叶,每株嫁接苗7~8 对。采集后将试验材料放入自封袋带回实验室,−70 ℃低温保存用于元素含量测定。
1.3 测定方法
生长指标测定:用游标卡尺(精确到0.01 mm)测定穗粗(距离嫁接口上方1 cm)及砧粗(距离嫁接口下方1 cm);卷尺测量穗高(嫁接口基部至苗顶端的长度)、节间长;叶面积仪测定叶面积、叶周长;天平(精确到0.001 g)测定叶鲜质量。
矿质元素测定:利用重铬酸钾滴定法测定全碳(TC)含量,利用H2SO4−H2O2法消煮后扩散法测定全氮(TN)含量、酸溶−钼锑抗比色法测定全磷(TP)含量、火焰光度计法测定全钾(TK)含量[17],并根据全碳和全氮指标计算碳氮比(C/N)。
1.4 数据处理
采用单因素方差分析2 个时期不同砧木嫁接苗的生长指标及叶片矿质元素含量差异,并用Duncan新复极差法进行多重比较;利用主成分分析确定代表生长势的生长指标并与叶片矿质元素含量进行相关性分析。数据统计分析均采用Microsoft Excel 及SPSS 25.0 软件,绘图软件为Origin。
2 结果与分析
2.1 不同砧木对薄壳山核桃嫁接苗生长的影响
2.1.1 不同砧木对薄壳山核桃嫁接苗嫁接成活率的影响
由图1 可知,4 类薄壳山核桃嫁接苗的成活率存在显著差异,其中'Pawnee/Stuart'和'Pawnee/Pawnee'的嫁接成活率均在90% 以上,显著高于'Pawnee/Jinhua'嫁接苗(P0.05)。
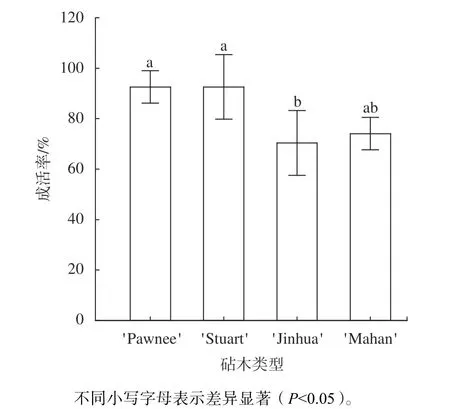
图1 不同砧木薄壳山核桃苗的嫁接成活率差异图Fig.1 Variation in survival rate of pecan seedlings grafted on different rootstocks
2.1.2 不同砧木对薄壳山核桃嫁接苗生长指标的影响
由表1 可知,在不同观测期嫁接苗穗粗、砧粗及穗高指标表现出一致性,均为'Pawnee/Pawnee'最高,其次分别为'Pawnee/Stuart'、'Pawnee/Mahan'和'Pawnee/Jinhua'。其中,'Pawnee/Pawnee'在这3项指标上与'Pawnee/Stuart'无显著差异,而'Pawnee/Mahan'和'Pawnee/Jinhua'均显著低于'Pawnee/Pawnee'和'Pawnee/Stuart'(P0.05)。6 月,'Pawnee/Pawnee'在穗粗、砧粗、穗高上分别高于'Pawnee/Jinhua'20.63%、14.99%、32.92%;9 月则分别高于'Pawnee/Jinhua'18.33%、21.60%、25.22%。
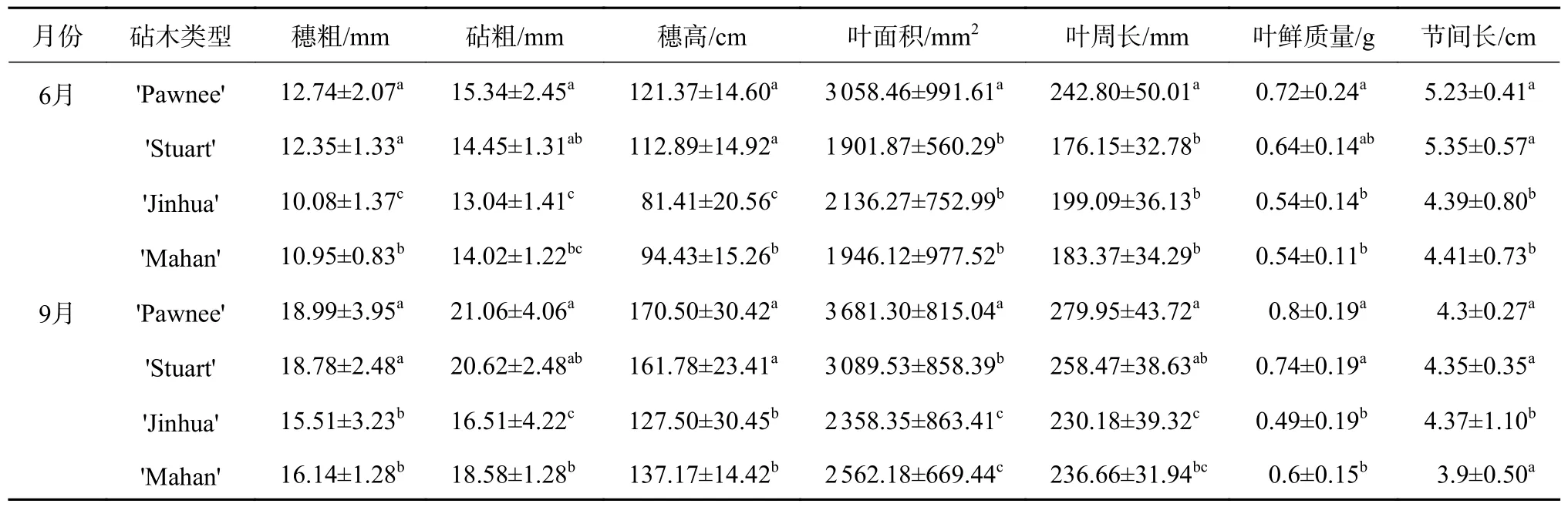
表1 不同砧木薄壳山核桃嫁接苗生长指标Table 1 Growth index of pecan seedlings grafted on different rootstock types
叶表型数据(叶面积、叶周长及叶鲜质量)仍以'Pawnee/Pawnee'最高,在6 月均显著高于其他3 类10%以上(P0.05);而在9 月,'Pawnee/Pawnee'则显著高于'Pawnee/Mahan'和'Pawnee/Jinhua'(P0.05),与'Pawnee/Stuart'差异不显著。
不同于上述指标,4 类嫁接苗间的节间长差异不显著,以'Pawnee/Stuart'最大,其次为'Pawnee/Pawnee'、'Pawnee/Mahan'和'Pawnee/Jinhua'。节 间长作为反映矮化效应的指标[18],其数值越小形成的株木树形越紧凑,因此推测'Pawnee/Jinhua'嫁接苗可能具有矮化特性。
2.1.3 不同砧木对薄壳山核桃嫁接苗生长势的影响
对6 月份单株各生长指标进行主成分分析,选择特征值大于1 且贡献率最高的2 个主成分(贡献率分别为57.2%和21.8%,累积79.0%)进一步绘制关于生长指标的双标图,如图2 所示,向量夹角大小表明各向量指标间均为正相关关系,余弦值表明穗高和节间长、叶周长和叶面积、砧粗与穗粗两两指标间的相关性最高,结合向量长度最终选择穗粗、穗高、叶面积及叶鲜质量4 个指标来代表生长势,以进行后续分析。
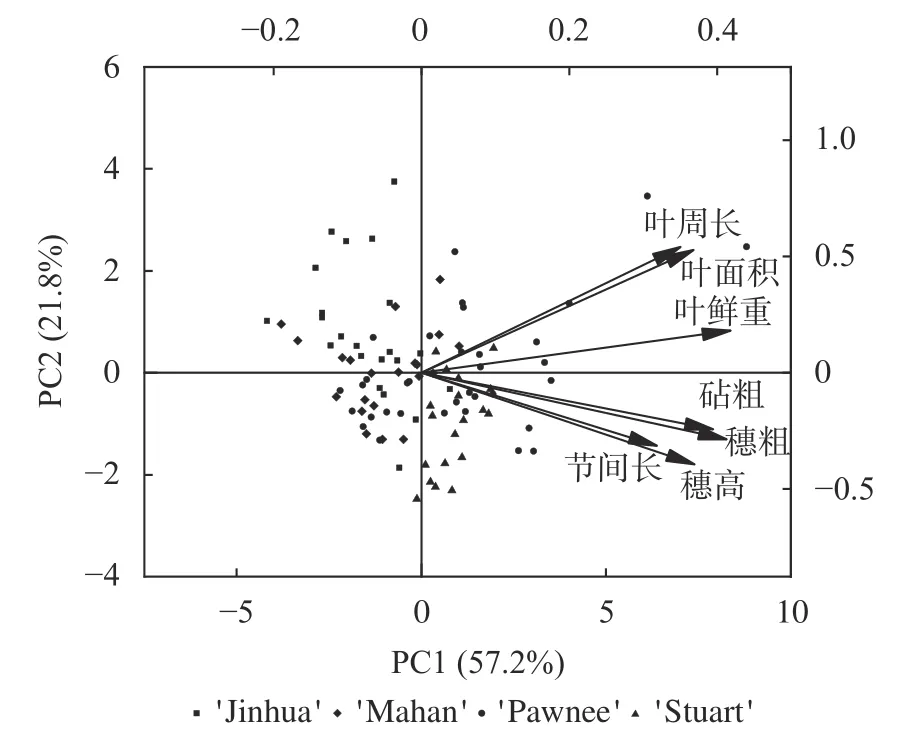
图2 生长指标主成分分析图Fig.2 PCA analysis of growth index
根据F=F1×57.18%+F2×21.81%,以选定主成分的方差贡献率为权数并量化各嫁接苗的2 个主成分得分,计算4 种薄壳山核桃嫁接苗的单株综合得分F,最终得分大小依次为'Pawnee/Pawnee'(1.027)、'Pawnee/Stuart'(−0.030)、'Pawnee/Mahan'(−0.585)和'Pawnee/Jinhua'(−0.666),说明嫁接后的生长情况以'Pawnee'和'Stuart'砧木表现优于'Mahan'和'Jinhua'。
2.2 不同砧木对薄壳山核桃嫁接苗叶片矿质元素含量的影响
2.2.1 不同砧木对嫁接苗叶片TN 含量的影响
由图3 可知,6 月份4 种类型的嫁接苗叶片TN 含量差异不显著,以'Pawnee/Jinhua'最高为18.27 g/kg,'Pawnee/Pawnee'最低为16.71 g/kg。观测期内,4 类嫁接苗的叶片TN 含量均有一定程度的降低,'Pawnee/Jinhua'的下降幅度最大,达到46.84%,'Pawnee/Stuart'和'Pawnee/Pawnee'的下降幅度较小,分别为15.54%、7.36%。9 月TN 含量大小以'Pawnee/Pawnee'和'Pawnee/Stuart'均在15 g/kg以上,显著高于'Pawnee/Mahan'、'Pawnee/Jinhua'(P0.05)。
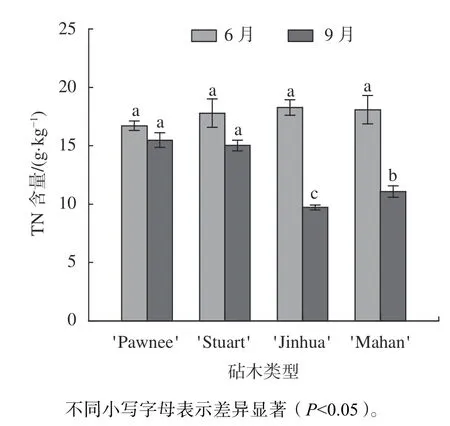
图3 不同砧木薄壳山核桃嫁接苗叶片TN 含量分析Fig.3 Analysis of total nitrogen content of leaf of pecan seedlings grafted on different rootstock types
2.2.2 不同砧木对嫁接苗叶片TP 含量的影响
由图4 可知,6 月份4 种类型的嫁接苗叶片中TP 含量为'Pawnee/Jinhua''Pawnee/Pawnee''Pawnee/Mahan''Pawnee/Stuart',其 中'Pawnee/Stuart'嫁接苗显著低于'Pawnee/Jinhua'20.52%(P0.05)。观测期内,4 类嫁接苗叶片的TP 含量出现不同的变化情况,其中'Pawnee/Pawnee'、'Pawnee/Mahan'和'Pawnee/Stuart'均有不同程度的下降,以'Pawnee/Mahan'13.78%的降幅最高,而'Pawnee/Jinhua'的TP含量变化基本持平。9 月,TP 含量大小以'Pawnee/Jinhua'最高为2.30 g/kg,显著高于'Pawnee/Pawnee'、'Pawnee/Stuart'、'Pawnee/Mahan'(P0.05),差异情况与6 月基本一致。
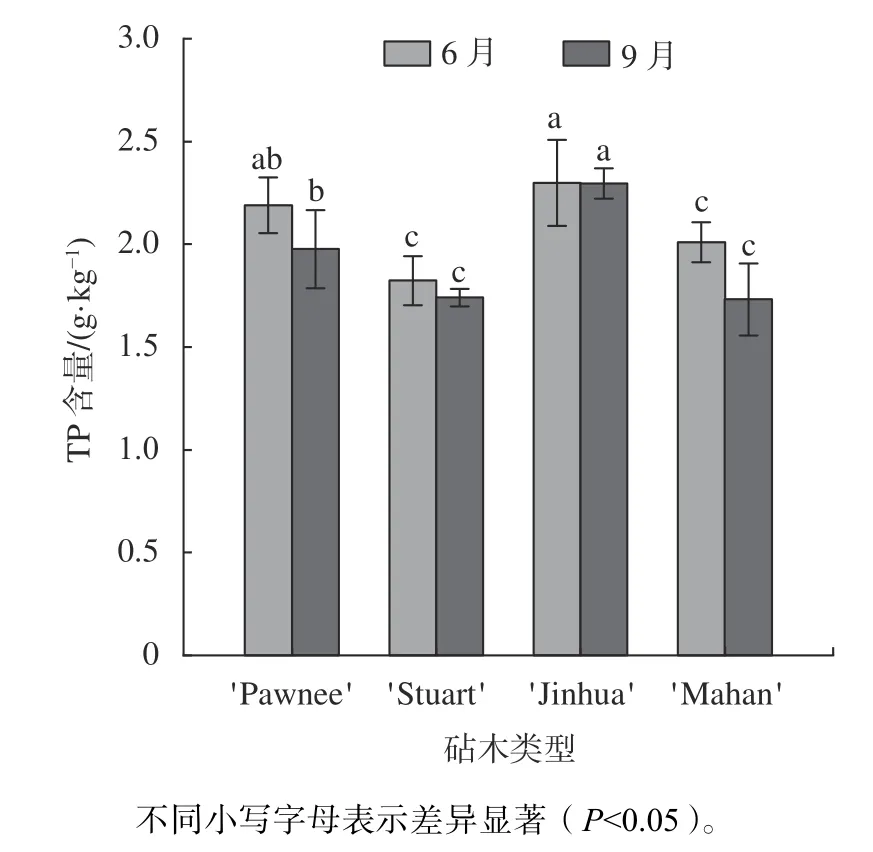
图4 不同砧木薄壳山核桃嫁接苗叶片TP 含量分析Fig.4 Analysis of total phosphorus content of leaf of pecan seedlings grafted on different rootstock types
2.2.3 不同砧木对嫁接苗叶片TK 含量的影响
由图5 可知,6 月份4 种类型的嫁接苗叶片中TK含量为'Pawnee/Pawnee''Pawnee/Jinhua''Pawnee/Stuart''Pawnee/Mahan',其 中'Pawnee/Pawnee'显著高于'Pawnee/Mahan' 26%(P0.05)。观测期内,4 类嫁接苗的叶片TK 含量发生明显的变化,除'Pawnee/Pawnee'有所下降7% 外,其余嫁接苗均呈现上升变化,上升幅度高至'Pawnee/Mahan'30.24%,低至'Pawnee/Stuart' 14.19%。9 月,TK 含量以'Pawnee/Jinhua'最高为10.69 g/kg,显著高于'Pawnee/Mahan'、'Pawnee/Stuart'、'Pawnee/Pawnee'(P0.05)。
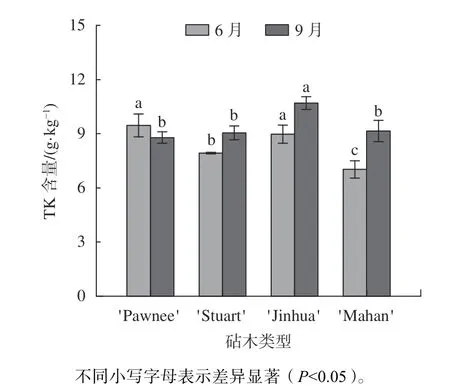
图5 不同砧木薄壳山核桃嫁接苗叶片TK 含量分析Fig.5 Analysis of total potassium content of leaf of pecan seedlings grafted on different rootstock types
2.2.4 不同砧木对嫁接苗叶片碳氮比含量的影响
由图6 可知,6 月份4 种类型的嫁接苗叶片C/N 间无显著差异,平均比值水平为25 左右。观测期内,4 类嫁接苗的C/N 均有所上升,以'Pawnee/Jinhua'、'Pawnee/Mahan'的上升幅度较高,分别为48.25%和36.61%。9 月,C/N 含量以'Pawnee/Jinhua 最高为45.84,显著高于'Pawnee/Mahan'、'Pawnee/Stuart'、'Pawnee/Pawnee'(P0.05),与K含量大小顺序一致。
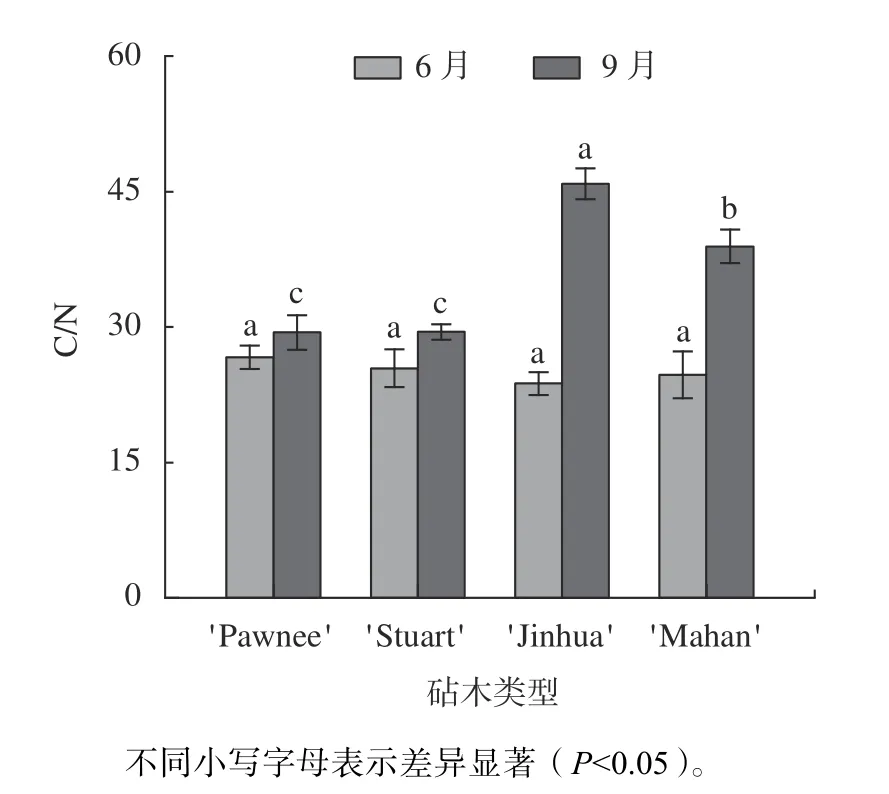
图6 不同砧木薄壳山核桃嫁接苗叶片碳氮比分析Fig.6 Analysis of C/N ratio of leaf of pecan seedlings grafted on different rootstock types
2.2.5 不同砧木嫁接苗叶片元素含量与生长势相关性分析
由表2 可知,叶片的TN 与生长势各指标间呈现正相关,其中与穗粗、穗高指标达到了极显著正相关(P0.01),相关系数分别为0.64 和0.90,说明氮元素的积累有利于促进嫁接苗茎干的生长;TP 含量与生长势关系不显著,而TK 与穗高指标间极显著负相关(P0.01),相关系数为−0.64;与TK 相同,C/N 与穗高指标也呈现相关系数为−0.84 的极显著负相关(P0.01)。
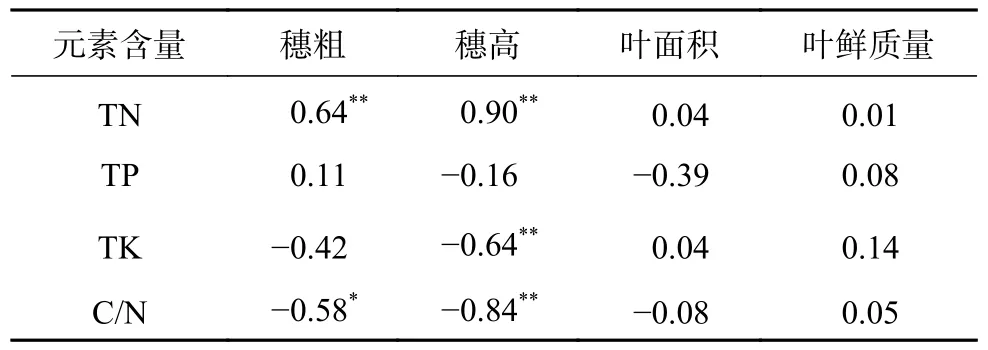
表2 叶片元素含量与生长势之间相关性分析Table 2 Analysis of correlation between element content of leaf and growth potential
3 结论与讨论
在嫁接过程中,砧木造成嫁接苗地上部分生长的差异,接穗影响嫁接苗根系发育情况,这种砧穗间的互作效应会直接表现在嫁接苗木的形态特征上[19]。本研究中,在砧粗、穗高、穗粗指标上占优的'Pawnee'、'Stuart'砧木嫁接苗在叶表型指标上同样具有明显优势,综合主成分得分判定四类嫁接苗生长势大小为'Pawnee/Pawnee''Pawnee/Stuart''Pawnee/Mahan''Pawnee/Jinhua',说明不同砧木对嫁接苗的生长形态产生了影响,推测这种长势差异与砧穗间的亲和性有关。研究表明遗传距离越近,砧穗的嫁接亲和性及相容性越高,嫁接过程中越易愈合成为共生体成活[20],最终影响嫁接苗的正常生长。黄展文等[21]在研究油茶(Camellia oleifera)与杜鹃红山茶(Camellia azalea)的砧穗组合时指出应该将早期嫁接苗成活率与后期生长状况进行综合判定,从而评价砧穗亲和性的优劣程度。因此综合生长势情况,嫁接成活率均在90%以上的'Pawnee'和'Stuart'砧木与'Pawnee'接穗的亲和性更高,可作为生产上的优良砧木,而'Jinhua'与'Pawnee'接穗的亲和性最差。'Pawnee'、'Stuart'、'Mahan'均为从美国引进的薄壳山核桃品种,而'Jinhua'是国内浙江地区选育品种,李晖等[22]在构建薄壳山核桃种质资源的亲缘关系树时发现,地理来源相同的品种大多可以聚在同一类群,所以地理间的差异可能是影响遗传距离远近的原因之一。
在本研究中,不同砧木对薄壳山核桃嫁接苗的叶片矿质元素含量具有影响,这与前人[23]关于砧木影响效应的研究是一致的。一般认为,砧木造成的矿质元素含量差异是嫁接苗营养生长、生殖生长及其他代谢生理过程出现差异的基础[24],因此本研究结果能够反映砧木基于部分营养机理对嫁接苗生长势的影响。
在观测生长期内,生长势与叶片TN 呈现正相关,与C/N 负相关,其中'Pawnee'和'Stuart'砧木嫁接苗的TN 较高,C/N 较低。Chaer 等[25]的研究指出叶片中的高氮含量有利于促进苗木内氮素循环,提高植株对于氮素的利用效率;低水平的C/N提供了低碳高氮的叶片环境,增加了养分供应量[26],从而促进地上部分的营养生长和生物量的增加。所以'Pawnee'和'Stuart'砧木嫁接苗在各项生长指标中均占有一定优势,生长势高于其他嫁接苗。碳代谢过程需要氮代谢中相关的酶及光敏色素等作为物质基础,氮代谢则需要碳代谢提供所需的能量[27],两者关系密切,因此当叶片C/N 过低时,会出现碳素不足从而限制氮素利用的情况。刘奇峰等[28]的研究发现增加氮素供给有利于提高碳代谢,加快光合产物的积累和植株的生长发育,所以嫁接苗生长过程中可以通过适当增加氮肥处理协调树体的碳氮平衡。另一方面,在营养生长阶段,植物会将氮素优先分配给地上部分,保证叶片中氮含量足够促进苗木发育[29],研究表明高氮环境会抑制地下根系的生长[30]。本研究中'Pawnee'和'Stuart'砧木嫁接苗在叶片TN 高的情况下表现出了较低的TP、TK,可能是因为嫁接苗地下生长受到了抑制,影响了根系对于磷、钾元素的吸收从而导致叶片TP、TK 含量较低。低磷、低钾的环境不利于后期花芽形成和成花成果[31−32],因此在生产实践中对这两类嫁接苗要注重磷肥和钾肥的施用。
另外,薄壳山核桃矮壮苗具有生长矮小,节间紧凑的特点[33]。研究表明矮化砧木嫁接后嫁接苗叶片的养分含量及营养物质积累诱导成花,促使果实早产[34],同时氮含量会有所下降[35],抑制营养生长。在本研究中,'Pawnee/Jinhua'嫁接苗在生长上表现为生长势较弱,节间长最小,树形紧凑,观测期内叶片中有利于生殖生长的TP、TK 及C/N 高,TN 较低,因此推测'Jinhua'砧木可以作为接穗'Pawnee'的矮化砧木,但由于营养元素并不能完全解释矮化机理,所以其矮化特性仍需进一步研究。
本研究中不同砧木对薄壳山核桃嫁接苗的生长及叶片矿质元素含量的影响存在差异。4 种砧木中,'Pawnee'和'Stuart'可作为接穗'Pawnee'生长表现较好的优良砧木,其叶片矿质元素特征为TN 含量高,在TP、TK 含量及C/N 能力上较弱,因此在生长后期,可通过调节氮、磷、钾配比的施肥方案来协调苗木营养平衡。但由于试验周期较短,分析仅限于观测生长期,所以日后可通过大田试验,进一步探究不同砧木嫁接叶片营养元素规律和对果实产量的影响,综合筛选出用于生产的薄壳山核桃优良砧木。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12 07:08:54
河北果树(2021年4期)2021-12-02 01:14:48
数学大王·中高年级(2021年3期)2021-03-10 11:19:30
中南林业科技大学学报(2020年9期)2020-09-28 09:00:20
农民致富之友(2018年18期)2018-07-14 05:54:18
小学生作文(中高年级适用)(2018年4期)2018-05-14 06:30:13
大科技(2016年4期)2016-07-13 07:24:33
广西林业科学(2016年1期)2016-03-20 05:33:14
中国蔬菜(2015年9期)2015-12-21 13:04:40
作文与考试·小学高年级版(2015年5期)2015-05-30 10:48:04
