
硫醚类香料对金黄色葡萄球菌和单增李斯特菌的抑制作用
2022-06-14 03:54许宇航毕景然侯红漫郝洪顺张公亮
中国食品学报 2022年5期
许宇航,王 岩,毕景然,侯红漫,郝洪顺,张公亮
(大连工业大学食品学院 辽宁大连 116034)
近年来,由于天然香料的分离技术不断地改进,天然香料在市场中的比例逐渐增加,其中含硫香料在食品香料和食品添加剂中发挥着越来越重要的作用[1]。含硫香料作为一种安全、无毒的天然香料,在美国食品香料和萃取物制造协会(FEMA)公布的一些安全性物质中,也占了很大的比例[2]。硫醚香料作为含硫香料中的代表,主要来源于大蒜素[3-4]。目前,对于硫醚类香料的抑菌构效研究较少,多用于抗癌的研究,一般通过干扰细胞周期来诱导癌细胞的凋亡[5]。
金黄色葡萄球菌和单增李斯特菌是食品中最常见的两种革兰氏阳性致病菌。金黄色葡萄球菌是多种临床感染的病原菌,它能够引发菌血症、感染性心内膜炎和骨关节感染,以及皮肤和软组织感染和化静脓肿形成[6-7]。单增李斯特菌可以进入人和动物的肠道,进而引起脑膜炎等疾病,感染人群主要集中于免疫力低下的人群,如孕妇、老人和新生儿等[8-9]。寻找一种新型、有效,可应用于食品行业的抑制剂,来控制食源性致病菌的传播和感染具有重要的研究意义。
耐热核酸酶与金黄色葡萄球菌侵染宿主细胞的能力有着重要关系,直接影响它的毒性,由nuc基因编码,它的存在能够增强细菌侵袭能力,同时耐热核酸酶也是检测金黄色葡萄球菌污染食品的重要特征标志[10-11]。本研究选取10 种硫醚类香料,通过测量抑菌圈的直径来比较和判断它们的抑菌效果,并讨论硫醚香料的结构与其抑菌能力的关系。同时也筛选出抑菌能力最强的硫醚类香料,研究其对金黄色葡萄球菌nuc 基因表达量的影响,为硫醚类香料作为食源性致病菌的抑制剂在食品行业中的应用提供一定的理论依据。
1 材料与方法
1.1 材料与试剂
本研究根据GB 2760-2014《食品安全国家标准 食品添加剂使用标准》[12]选取了10 种硫醚类香料,均从美国Sigma 公司订购。名称及结构见表1。

表1 10 种硫醚类香料的名称及结构Table 1 Name and structure of ten kinds of sulfide flavors
金黄色葡萄球菌ATCC 6538、单增李斯特菌ATCC 19115 取自大连工业大学食品学院微生物实验室。
RNAprep pure 培养细胞/细菌总RNA 提取试剂盒,天根生化科技 (北京) 有限公司;SYBRRPremix Ex TaqTM、Random primers[pd(N)6]、Reverse Transcriptase M-MLV (RNase H-)、5×MMLV Buffer、dNTP Mixture、Recombinant RNase Inhibitor,宝生物工程(大连)有限公司;RNase-free water、荧光定量聚合酶链式反应(Polymerase chain reaction,PCR)引物,生工生物工程(上海)股份有限公司;胰蛋白胨、酵母浸粉、氯化钠、琼脂粉、大豆蛋白胨、磷酸二氢钾、葡萄糖(分析纯级),大连博诺生物化学试剂公司;无水乙醇(分析纯级),沈阳力诚试剂厂。
1.2 仪器与设备
FA(A)型电子天平,上海民桥精密科学仪器公司;01J2003-04 型高压灭菌锅,上海博讯实业有限公司;ZHWY-1008 型恒温培养振荡器、ZHWY-2102C 型双层小容量全温度恒温摇床,上海智城分析仪器制造有限公司;DRP-9162 型电热恒温培养箱,上海森信实验仪器有限公司;UV-1750 型紫外-可见光分光光度计,日本津岛仪器有限公司;MyiQTM2 型荧光定量PCR 仪,美国Bio-Rad 公司。
1.3 方法
1.3.1 硫醚香料的处理 选择乙醇作为硫醚类香料的溶剂,所需原液和无水乙醇的体积可按如下公式计算出,稀释至适当浓度后,置于4 ℃冰箱密封备用。

式中,c——稀释的硫醚类香料的浓度,mol/L;ρ——硫醚类香料的密度,g/mL;V1——硫醚类香料的体积,L;M——硫醚类香料的摩尔质量,g/mol;V2——无水乙醇的体积,L。
1.3.2 培养基的制备 金黄色葡萄球菌用溶菌肉汤(Lysogeny broth,LB 培养基)培养,每配置100 mL 培养基需要添加1 g 胰蛋白胨、0.5 g 酵母浸粉和1 g 氯化钠,用100 mL 去离子水将其混匀后,调节pH 值至7.0,在121 ℃下灭菌20 min。
单增李斯特菌用胰蛋白胨大豆肉汤培养基培养,每配置100 mL 培养基需要添加1.7 g 胰蛋白胨、大豆蛋白胨0.3 g、氯化钠0.5 g、磷酸二氢钾0.25 g、葡萄糖0.25 g,最后用100 mL 去离子水将其混匀后,调节pH 值至(7.3±0.2),在121 ℃下灭菌20 min。
在配制固体培养基时,都需要向两种培养基中添加1.5 g/100 mL 的琼脂粉。
1.3.3 菌种的活化及培养 取-80 ℃冰箱中冻存的菌液100 μL 于100 mL 的液体培养基中进行第1 次传代,在37 ℃下震荡培养24 h 后,在固体培养基上进行第2 次传代,在37 ℃恒温箱培养12 h后,挑取单菌落至液体培养基并在37 ℃下震荡培养12 h。配制无菌的生理盐水并用其将上述得到的菌液稀释至107CFU/mL,保存于4 ℃冰箱中备用。
1.3.4 抑菌圈的测定 抑菌圈的测定采用了滤纸片法[13-14]。取100 μL 上述被稀释后的菌液于固体培养基上,用玻璃珠摇匀,使菌液能够均匀地分布在整个固体培养基上,将准备好的无菌直径为1 cm 的滤纸片放置平板中央,添加6 μL 浓度为5 mol/L 的硫醚类香料于滤纸片上,最后将固体培养基放置于37 ℃的培养箱中培养18~24 h。每种硫醚类香料进行3 次平行试验,最终结果取平均值。
1.3.5 最低抑菌浓度的测定 根据1.3.4 节试验结果选取抑菌效果最好的硫醚类香料作为研究对象,用二倍稀释法测定它的最低抑菌浓度(Minimum inhibitory concentration,MIC)。用60%的乙醇水溶液将5 mol/L 的硫醚类香料进行2 倍稀释,分别为2.5,1.25,0.625,0.3125 mol/L 以 及0.15625 mol/L。精确量取0.1 mL 菌液添加至0.8 mL 的液体培养基中,再将0.1 mL 不同浓度的硫醚类香料分别添加至液体培养基中,震荡混匀之后,在37 ℃下培养24 h。再取培养完之后的菌液100 μL 接种于固体培养基中,用玻璃珠震荡混匀后,在37 ℃下培养24 h[15]。最后观察固体培养基上是否有菌生长,并且以无菌生长的硫醚类香料的最低浓度为MIC。每个浓度做3 次平行试验,最终结果取平均值。
1.3.6 硫醚类香料对生长曲线的影响 根据1.3.4节和1.3.5 节结果,将抑菌能力最强的硫醚类香料稀释至亚抑菌浓度1/16MIC、1/8MIC、1/4MIC、1/2MIC 和MIC。先取1 mL 的菌液于99 mL 的液体培养基中培养,再将0.1 mL 各亚抑菌浓度下的硫醚类香料分别滴加到液体培养基中,以仅添加0.1 mL 无水乙醇的菌液为对照组,37 ℃下震荡培养24 h,并且每隔2 h 测一次菌液在波长600 nm 下的吸光度值,以吸光度值为纵坐标,时间为横坐标,绘制供试菌的生长曲线。
1.3.7 RNA 的提取和反转录荧光定量PCR 在液体培养基中添加各亚抑菌浓度的硫醚类香料,以未添加硫醚香料的为对照组,培养12 h 后,取适量的菌液,按照RNA 试剂盒的说明书提取RNA 并反转录为cDNA 即可[16-17]。nuc 基因引物:上游5'-GCGATTGATGGTGATACGGTT-3'、下游5'-AGCCAAGCCTTGACGAACTAAAGC-3'[18];16S rRNA 内参基因引物:上游5'-GCTGCCCTTTGTATTGTC-3'、下游5'-AGATGTTGGGTTAAGTCCC-3'[19]。最后在65~90 ℃的范围内建立熔解曲线,采用方程法2-ΔΔCT来确定基因的相对表达量[20]。
1.4 数据分析与处理
所有的数据都以平均值±标准差来表示,并均用SPSS 软件进行显著性分析。
2 结果与分析
2.1 硫醚类香料对金黄色葡萄球菌和单增李斯特菌的抑制效果
将各硫醚类香料都稀释至5 mol/L,在此浓度下对金黄色葡萄球菌和单增李斯特菌的抑制效果见表2。结果表明,在10 种硫醚类香料中,有9 种抑制了金黄色葡萄球菌的生长,有7 种抑制了单增李斯特菌的生长,说明单增李斯特菌对硫醚类香料不如金黄色葡萄球菌敏感,其中DMTS、DADS、DAS、MMTF、MFDS、FIPDS 对金黄色葡萄球菌和单增李斯特菌均有抑制作用,MPDS、DPDS和BMFDS 只对金黄色葡萄球有抑制作用。

表2 10 种硫醚类香料抑制作用Table 2 The inhibitory effect of ten kinds of isulfide flavors
2.2 硫醚类香料的抑菌作用
选取具有抑菌能力的硫醚类香料进行抑菌试验,得到的抑菌圈结果如图1所示。在图1a 中,9种硫醚类香料对金黄色葡萄球菌的抑制作用各不相同,抑菌能力依次为:BMFDS≈DADS>MFDS≈DAS >FIPDS ≈MMTF >DMTS ≈MPDS ≈DPDS。BMFDS 和DADS 有显著的抑菌效果,抑菌圈直径分别达到(2.83±0.21)cm 和(2.46±0.35)cm。在图1b 中,7 种有抑菌作用的硫醚类香料对单增李斯特菌的抑菌能力为:DADS >MFDS >MMFDS ≈MMT F≈FIPDS>DAS≈DMTS,其中DADS 有最强的抑菌效果,抑菌圈直径为(1.98±0.22)cm。
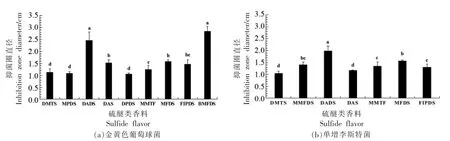
图1 硫醚类香料对金黄色葡萄球菌(a)和单增李斯特菌(b)的抑制作用Fig.1 Inhibitory effects of sulfide flavors on Staphylococcus aureus (a) and Listeria monocytogenes (b)
MPDS 和DADS 虽然含有相同数目的硫原子,但是侧链不一样,MPDS 的-S-S-两侧连接的基团是甲基和丙基,DADS 是烯丙基,DADS 对金黄色葡萄球菌的抑菌效果比MPDS 强 (P<0.05),并且根据2.1 节得出MPDS 对单增李斯特菌无明显抑制作用,可以推测出由于DADS 的侧链中烯丙基或是双键的存在,增强了硫醚类香料的抑菌活性。这与前期研究[21]含有双键可能会影响二硫醚类化合物的抑制效果,存在二烯丙基的单硫醚化合物会有很好的抑制效果的结论相似。
DADS 和DAS 虽具有相同的侧链烯丙基,但是前者拥有的硫原子多于后者,DADS 中有2 个硫原子形成了二硫键。从图1结果可以看出,DADS 对金黄色葡萄球菌和单增李斯特菌的抑制能力均强于DAS(P<0.05),可见对于结构简单的对称型硫醚类香料,侧链的基团相同,随着所含硫原子数目的增多,抑菌能力增强。
MMFDS 和MFDS 是同分异构体,它们的分子式都是C6H8OS2,MMFDS 二硫键连接着呋喃基,MFDS 连接的基团是糠基,MFDS 对金黄色葡萄球菌的抑菌效果更强,而MMFDS 无抑菌作用,显示出对于MMFDS 和MFDS,含有糠基的硫醚类香料的抑菌效果比呋喃基强。对于MMFDS 和MMTF,两者所含的硫原子数目不一样,侧链所含的基团相同,对于单增李斯特菌,两者的抑菌效果无明显差异(P>0.05),说明如果硫醚类香料的结构比较复杂,所含有硫原子数目的多少对其抑制作用没有太大的影响。
由此可见,硫醚类香料的结构对于它们的抑菌活性有着重要的影响,硫原子的数量和侧链的结构在很大程度上决定了它们的抑菌能力,这一结论与前期研究含氧硫醚类香料的抑菌作用的结论类似[22]。根据本研究的结果可以看出,DADS 由于硫原子的数目多和侧链上带有烯丙基的结构,导致了它的抑菌能力明显强于其它的硫醚类香料。与此结论类似的是,有研究表明异硫氰酸酯类含硫香料由于它们侧链的结构不同,同样导致了它们对金黄色葡萄球菌和单增李斯特菌的抑制效果不同,侧链上的苯基能够大幅度提高异硫氰酸酯类香料的抑菌效果[23]。
2.3 DADS 对金黄色葡萄球菌和单增李斯特菌的MIC
选取对金黄色葡萄球菌和单增李斯特菌抑菌效果最好的DADS 作为研究对象,测定其MIC,结果见表3。当DADS 的浓度大于或者等于1.25 mol/L 时,两种菌都无法生长,该浓度的DADS 对这两种革兰氏阳性致病菌都有着很强的抑制作用。当DADS 的浓度低于1.25 mol/L 时,可以观察到单增李斯特菌能生长,因此,单增李斯特菌的MIC 为1.25 mol/L;当DADS 的浓度低于0.3125 mol/L 时,可以观察到金黄色葡萄球菌能生长,因此,金黄色葡萄球菌的MIC 为0.3125 mol/L。
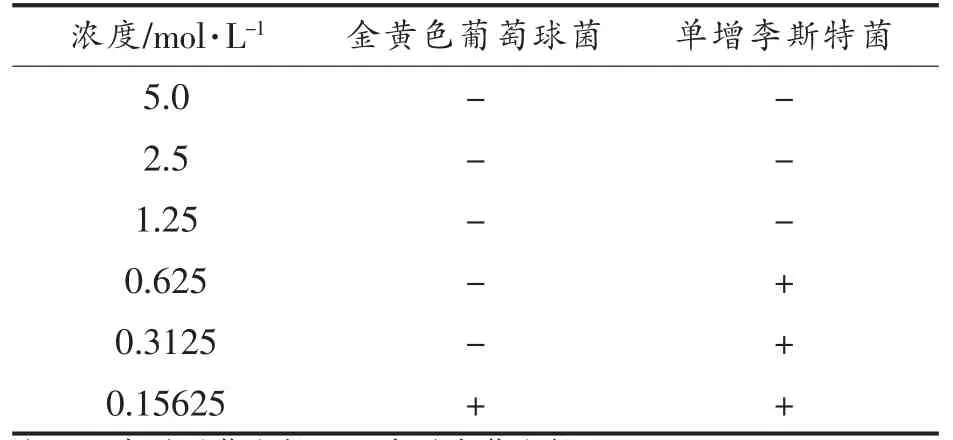
表3 DADS 对两种革兰氏阳性致病菌的MICTable 3 MIC of DADS against two Gram-positive pathogens
2.4 DADS 对两种革兰氏阳性致病菌生长曲线的影响
在亚抑菌浓度下的DADS 对两种革兰氏阳性菌正在曲线的影响见图2。当DADS 的浓度在1/16MIC 和1/8MIC 时,金黄色葡萄球菌和单增李斯特菌的生长并没有受到明显的影响,它们的生长曲线与对照组几乎一致;当DADS 的浓度提高到1/4MIC 时,此时能够观察到两种菌的生长开始受到抑制,只是在该浓度下,DADS 对金黄色葡萄球菌的抑制作用并不明显,而该浓度的DADS 对单增李斯特菌已经有了相当明显的抑制效果,当进入稳定期后,OD600nm的值仅有对照组的一半,菌液的密度已经大幅度减少;当DADS 的浓度提高到1/2MIC 时,此时两种革兰氏阳性菌的生长均受到了明显的抑制;当DADS 的浓度继续提高至MIC 时,可以看到两种菌几乎停止了生长。由此可见,DADS 的浓度在1/16MIC~MIC 时,它对金黄色葡萄球菌和单增李斯特菌的抑制作用都具有浓度依赖性。与此结论类似的是,DADS 对大肠杆菌的抑制同样具有浓度依耐性[24]。
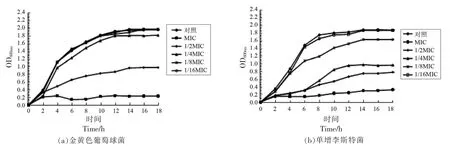
图2 DADS 对金黄色葡萄球菌(a)和单增李斯特菌(b)生长曲线的影响Fig.2 Effect of DADS on the growth curve of Staphylococcus aureus (a) and Listeria monocytogenes (b)
2.5 DADS 对金黄色葡萄球菌毒力基因nuc 表达的影响
如图3所示,与未添加DADS 的对照组相比,添加不同亚抑菌浓度的DADS 后,毒力基因nuc的表达水平均显著降低(P<0.01)。DADS 的亚抑菌浓度由1/64MIC、1/32MIC、1/16MIC、1/8MIC 增加至1/4MIC,与未添加DADS 相比,nuc 的相对表达水平分别为77.08%,57.59%,48.7%,28.29%和15.22%。结果表明:逐渐增加DADS 的浓度,nuc基因的相对表达水平逐渐降低,即DADS 浓度增加,对nuc 基因的抑制作用增强,具有明显的浓度依耐性。与本研究结论类似的是,有研究表明大蒜素同样以一种剂量依赖性的方式来抑制金黄色葡萄球菌hla 基因的表达水平,而大蒜素的主要成分就是二烯丙基二硫醚等硫醚类香料[19]。也有研究表明,DADS 对单增李斯特菌毒力基因inlA、hylA 和prfA 的表达水平也具有一定的抑制作用,并且也呈浓度依耐性[25]。DADS 对大肠杆菌的毒力基因同样也存在着抑制作用,随着DADS 的浓度提升,大肠杆菌的毒力基因的表达水平逐渐降低[24]。综合来看,DADS 对食源性致病菌的毒力基因的表达具有很强的抑制作用,随着浓度的提升,对其表达水平的抑制也增强,具有浓度依耐性。将DADS 与其它含硫香料相比较,在周倩倩等[26]的研究中,异硫氰酸苄酯(Benzyl isothiocyanate,BITC) 的浓度为1/4MIC 时,此时副溶血弧菌tdh基因的表达水平相比于对照组降低了4.8 倍,与本研究结果相类似,可见硫醚类香料和异硫氰酸酯类香料对于食源性致病菌毒力基因的表达均有着很强的抑制能力。
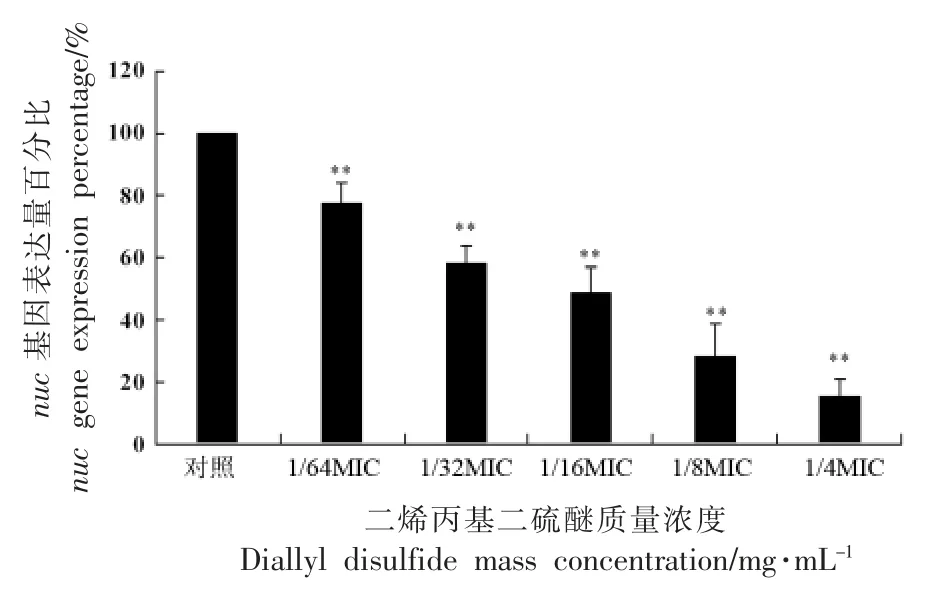
图3 二烯丙基二硫醚对nuc 基因表达的影响Fig.3 Effects of diallyl disulfide on expression of nuc gene
3 结论
在本研究选取的10 种硫醚类香料中,DADS、MFDS 和DAS 均对金黄色葡萄球菌和单增李斯特菌有着不错的抑制效果,其中抑制能力最强的是DADS,原因可能是DADS 侧链中烯丙基的存在以及硫原子的数目较多,从而增强了硫醚类香料对这两种革兰氏阳性致病菌的抑制能力。DADS 对金黄色葡萄球菌和单增李斯特菌的MIC 分别为0.3125 mol/L 和1.25 mol/L。DADS 对两种菌的抑制作用均呈现浓度依赖性。DADS 在亚抑菌浓度下同样以浓度依赖性的方式降低了金黄色葡萄球菌耐热核酸酶基因nuc 的表达水平。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
小学科学(学生版)(2021年9期)2021-11-02
山东农业大学学报(自然科学版)(2021年3期)2021-07-29
——香草传
动漫星空(兴趣百科)(2020年11期)2020-11-09
疯狂英语·新悦读(2020年7期)2020-07-30
癌变·畸变·突变(2020年1期)2020-02-12
疯狂英语·新阅版(2019年10期)2019-09-10
水运管理(2018年5期)2018-08-20
健康之路(医药研究)(2015年2期)2015-10-21
中国新技术新产品(2015年6期)2015-07-20
