
丹参SmVS 基因的克隆和表达分析
2022-06-10 10:30吴丽萍苏兴隆曹梦阳王兆健邢世海
热带亚热带植物学报 2022年3期
吴丽萍, 苏兴隆, 曹梦阳, 王兆健, 邢世海,2*
(1. 安徽中医药大学药学院,合肥 230012;2. 安徽省中医药科学院中药资源保护与开发研究所,合肥 230012)
丹参(Salvia miltiorrhiza)为唇形科(Labiatae)鼠尾草属多年生直立草本植物,药用部分为干燥根和根茎,为我国常用的传统中药材[1]。丹参主产于安徽、四川、山西、江苏、河北等地,其性微寒、味苦、归心、肝经,是治疗心脑血管疾病的传统中药材[2]。现代药理研究表明,丹参具有保护心血管系统[3]、防止动脉粥样硬化[4]、保护神经系统[5]、改善微循环[6]和抗炎抗菌[7]等多种药理作用,临床常用于心脑血管疾病、癌症、糖尿病和肝病等的治疗[8-10]。现代化学分析表明丹参的化学成分以脂溶性二萜丹参酮类和水溶性丹酚酸类为主,还有多糖类、黄酮类、甾体类等成分[11-12];近年来还报道丹参中含有生物碱成分[13]。目前对丹参的研究大多集中在丹参酮和丹酚酸上,其生物合成途径大部分已被解析[14-15];而关于丹参生物碱的研究甚少,尚无其合成途径的报道。在生物碱中,萜类吲哚生物碱是一类非常重要的具有萜环和吲哚骨架的生物碱化合物,具有显著的药理学活性,临床主要用于抗癌、抗心律失常和抗疟疾等[16]。
单萜吲哚类生物碱阿吗灵,其合成路径已比较透彻[17],其中Vinorine 合成酶(vinorine synthase, VS)是阿吗灵合成途径中的1 个关键酶,催化蛇根精型生物碱16-epi-vellosimine 可逆性地合成阿吗灵型生物碱Vinorine,Vinorine 合成酶连接2 种不同类型的生物碱,即蛇根精型(Sarpagan)和阿吗灵型生物碱(Ajmalan)[17]。Vinorine 合成酶是第一个三维结构被解析的酰基辅酶A 依赖型酰基转移酶(BAHD)家族成员,BAHD 得名于从植物中分离出的前4 种家族酶,该家族成员在多种次生代谢产物的生物合成中起着重要作用[18-19],但目前在丹参中尚未见报道。本研究从丹参中克隆SmVS基因,对其编码蛋白进行生物信息学分析,利用实时荧光定量PCR 技术(RT-qPCR)检测SmVS基因的表达模式,为进一步研究其功能提供理论基础。
1 材料和方法
1.1 材料
材料采自安徽中医药大学药园,生长光/暗周期为16 h/8 h,经安徽中医药大学俞年军教授鉴定为唇形科丹参(Salvia miltiorrhiza)。采集丹参后用自来水清洗表面杂质,置于冰盒中转移至实验室,用蒸馏水冲洗3 次,吸干水分立即用液氮冷冻提取总RNA。
1.2 总RNA 提取和cDNA 的合成
采用改良的TRIzol 法[20]提取丹参根和叶的总RNA,使用FastQuant RT Kit (Tiangen Biotech,北京)试剂盒合成cDNA。
1.3 引物设计和PCR 扩增
根据丹参转录组数据库[21],查找与SmVS同源的cDNA 序列用Primer 5.0 软件设计引物(表1)进行特异PCR 扩增。引物序列由上海生工生物工程股份有限公司合成。

表1 引物序列Table 1 Primer sequences
1.4 生物信息学分析
根据PCR 扩增得到的基因全长序列,利用ORF Finder 网站(http://www.ncbi.nlm.nih.gov/projects/gorf/)寻找开放阅读框(ORF),并翻译出氨基酸序列;在线进行序列同源性比对(http://blast.Ncbi.Nlm.Nih.gov),并进行蛋白质理化性质分析(http://web.expasy.org/protparam); 用TMHMM 在线程序(http://www.cbs.dtu.dk/services/TMHMM-2.0)分析蛋白质的可能跨膜区;利用PSORT 软件(https://www.genscript.com/psort.html)预测蛋白的亚细胞定位;利用GOR4 软件(https://npsa-prabi.ibcp.fr/cgi-bin/secpred_sopma.pl)预测蛋白的二维结构;用SWISS-MODEL 软件(https://swiss model.expasy.org/)进行编码蛋白的三维结构预测;并利用MEGA X 软件(Neighbor-joining,邻位相连法)构建系统进化树。
1.5 SmVS 基因表达的RT-qPCR 分析
采用SYBR Premix Ex TaqTM试剂(TaKaRa)和Roche Z480 实时荧光定量PCR 仪,以GAPDH作为内参基因,设计定量引物SmVS-RT (表1),参照韩荣春等[22]的方法进行RT-qPCR 反应,比较SmVS基因在叶和根中的表达差异,并分析其在叶中表达的日变化规律(n=3)。
2 结果和分析
2.1 SmVS 基因的克隆
采用特异性引物进行PCR 扩增,取5μL 产物进行凝胶电泳检测,结果表明克隆得到的条带与转录组预期基因片段大小一致(图1)。将其核苷酸序列(图2)与其他植物VS基因进行比对,表明该基因与其他植物的同源基因具有很高相似性,因此命名为SmVS。基因全长为726 bp,编码241 个氨基酸(图2)。该基因在GenBank 的登录号为MW802630。

图1 SmVS 基因PCR 扩增产物电泳图。M: DL2000; 1~3: SmVS 基因。Fig. 1 Electrophoretic diagram of PCR amplification products of SmVS. M:DL2000 marker; 1-3: SmVS.

图2 SmVS 基因的核苷酸序列和编码的氨基酸序列Fig. 2 Nucletide sequence of SmVS and encoded amino acid sequence
2.2 生物信息分析
利用ProtParam 软件对SmVS编码的蛋白进行理化性质分析, 结果表明,SmVS 蛋白分子式为C1200H1857N329O347S13,由20 种共241 个氨基酸组成,相对分子量为26 861.72,原子总数为3 746,等电点(pI)为5.66,不稳定系数为43.95,预估在哺乳动物体外的网织红细胞的半衰期是30 h、酵母体内大于20 h、大肠杆菌体内大于10 h,脂肪族指数74.02,总平均亲水性为-0.208。跨膜区分析表明SmVS 未形成跨膜区结构域,属于非跨膜蛋白。预测SmVS蛋白的亚细胞定位于线粒体,可能性为73.9%;定位于细胞质的可能性为17.4%;定位于细胞质膜和细胞核的可能性较低,均为4.3%。
蛋白质二级结构域预测表明(图3: A),约有35.27%的氨基酸(85 个)以α螺旋形式存在,17.01%的氨基酸(41 个)为延伸主链,4.98%的氨基酸(12 个)为β转角,42.74%的氨基酸(103 个)为无规则卷曲,表明该蛋白二级结构以α螺旋和无规则卷曲为主。用SWISS-MODEL对SmVS蛋白的氨基酸序列进行三维结构预测(图3: B),该蛋白的主要结构元件还是螺旋、折叠和卷曲,与二级结构的预测结果相符。
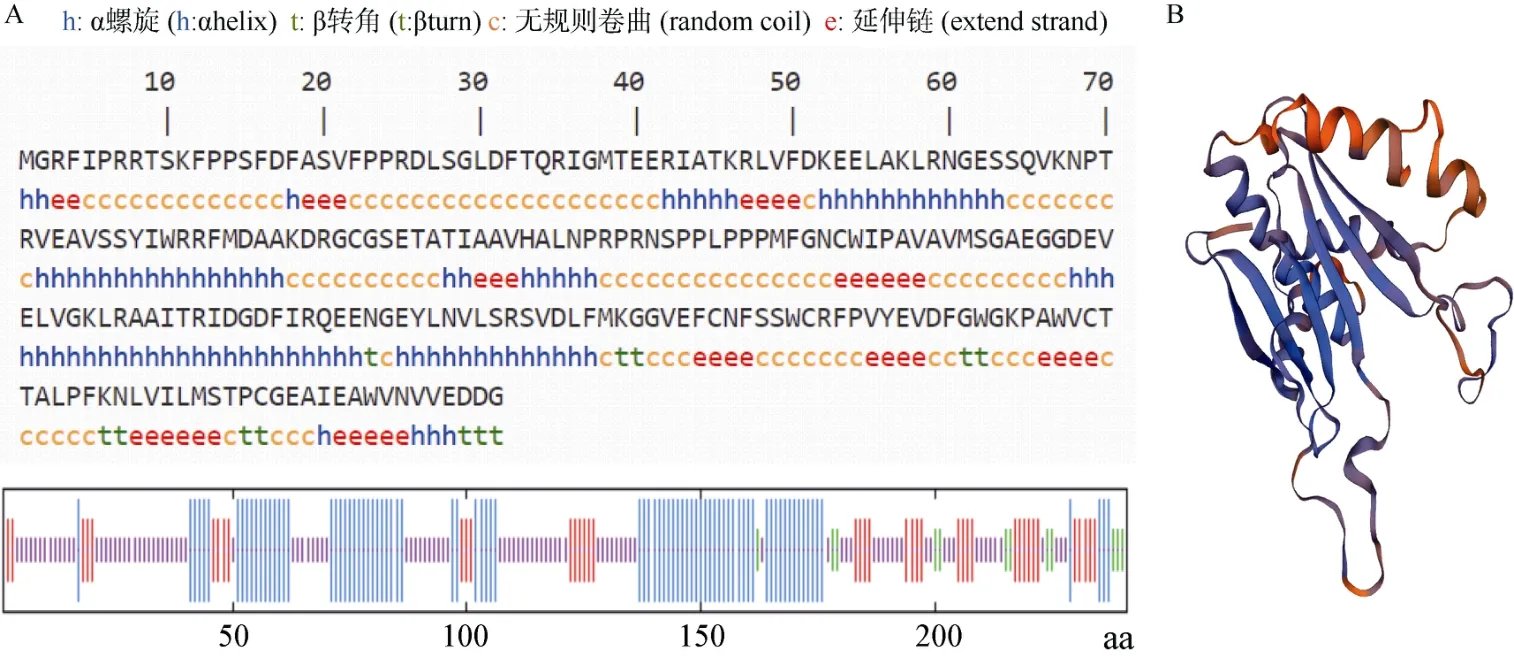
图3 SmVS 蛋白二级结构(A)和三维结构(B)预测Fig. 3 Prediction of secondary (A) and three dimensional (B) structures for SmVS
2.3 SmVS 蛋白的系统进化分析
选取6 种植物的VS 氨基酸序列,利用DNA MAN 软件进行序列多重比对分析(图4: A),结果表明,这些VS 蛋白间具有较高的相似性。同时,SmVS蛋白功能域的氨基酸组成与其他植物几乎一致,具有相同的保守域(Motif 1)(图4: B),其中DFGWG结构域是几乎所有BAHD 家族成员都有的,然而该结构域在不同酶中会有些许差异,如DFGWG 结构域在杨树(Populussp.)的BAHD 家族基因中成为DYGWG、DFGFG、DFGWA、NFGWG、DFGWK、DLGFG 和NLGWG[18]。

图4 VS 的氨基酸序列比对(A)和保守域(B)分析Fig. 4 Alignment of amino acid sequence and conserved domain of VS
根据NCBI Blast 对比结果,SmVS 蛋白的氨基酸序列与其他植物的同源性较高,与野生油橄榄(Olea europaeavar.sylvestris)的VS 氨基酸序列相似度为64%。在MEGA X 软件平台上采用NJ 法(bootstrap 设置为1 000)对11 种植物的VS 氨基酸序列进行系统进化分析(图5),包括丹参(MW802630)、野生油橄榄(LOC111382475)、欧基尼奥伊德斯种咖啡(Coffea euge-nioides, LOC113763265)、烟草(Nicotiana tabacum, LOC 107765976)、茶树(Camellia sinensis, LOC114266018)、牵牛(Ipomoea nil, LOC109155698)、芝麻(Sesamum indicum, LOC105168028)、苦瓜(Momordica charantia,LOC111007275)、南瓜(Cucurbita moschata, LOC11 1439839)、哥伦比亚锦葵(Herrania umbratical, LOC11 0418295)和胡桃(Juglans regia, LOC108985508)。结果表明,SmVS 与野生油橄榄的VS 聚为一支,亲缘关系最近, 其次是欧基尼奥伊德斯种咖啡。系统进化和序列比对结果一致, 相似性越高亲缘关系越近,可见Vinorine 合成酶在进化过程中是相对保守的。

图5 基于氨基酸序列的VS 系统进化树Fig. 5 Phylogenetic tree of VS based on amino acid sequences
2.4 实时荧光定量表达分析
为了解SmVS基因在丹参中的表达模式,以GAPDH为内参基因,采用RT-qPCR 技术检测SmVS基因的相对表达量。可见,SmVS基因在根和叶中均有表达,根中的表达是叶中的近2 倍(图6: A), 且叶中SmVS基因在21:00 达最大值(图6: B)。

图6 SmVS 基因的表达模式。柱上不同字母表示差异显著(P<0.05)。Fig. 6 Expression pattern of SmVS. Different letters upon column indicate significant differences at 0.05 level.
3 结论和讨论
本文从丹参中克隆得到了SmVS基因,全长为726 bp。生物信息学分析表明,SmVS基因编码241个氨基酸,SmVS 蛋白的分子量为26 861.72,等电点PI 为5.66,为非跨膜蛋白,推测其定位于线粒体。氨基酸同源性分析表明,SmVS 蛋白功能域的氨基酸组成与其他植物的VS 有较高的一致性,具有相同的保守域(Motif 1),SmVS 蛋白具有BAHD 家族的保守结构域DFGWG,因此,推测SmVS基因可能属于BAHD 家族成员。系统进化树分析表明丹参的SmVS 蛋白与野生油橄榄亲缘关系较近。RT-qPCR结果表明,SmVS基因在丹参根中的表达量比叶中高,说明其在丹参中具有组织特异性;叶中SmVS在21:00 时表达最高。这些为丹参生物碱的代谢调控研究提供了基础数据。
单萜类吲哚生物碱阿吗灵,1931 年首次从蛇根木(Rauvolfia serpentina)树皮中分离得到[23],1959年上市,属于Ia 类抗心律失常药物,临床上主要用于治疗预激综合征伴发的心律失常[24]。乙酰辅酶A依赖酶的Vinorine 合成酶是从蛇根木(Rauvolfifia serpentina)和直瑞兹亚(Rhazya stricta)的杂交细胞悬浮培养物中分离出来的[25]。乙酰辅酶A 依赖的酰基转移酶在植物细胞的次生代谢中起重要作用[26]。用于癌症治疗的二聚生物碱长春碱和长春新碱的生物合成前体单萜吲哚生物碱长春碱在长春花(Catharanthus roseus)的生物合成过程中,发生了乙酰转移[27-28];二萜生物碱紫杉醇在东北红豆杉(Taxus cuspidata)中的生物合成中,涉及到几种酰基转移酶,其中有2 种乙酰辅酶A 依赖性酶[29]。在阿吗灵生物合成途径中,乙酰辅酶A 依赖型Vinorine合成酶通过连接2 种不同类型的生物碱, 即蛇根精和阿吗灵生物碱,在合成代谢途径中起重要作用。
SmVS编码的氨基酸序列和其他6 种植物具有相同的保守域,即SmVS 蛋白具有BAHD 家族成员的保守结构域DFGWG,因此,推测SmVS 属于BAHD 家族的一员。最近几年报道的BAHD 家族成员数目不断增长,目前在水稻(Oryza sativa)、杨树、拟南芥(Arabidopsis thaliana)中分别有119、94 和64个相关基因[30-31],而丹参中还未见报道。BAHD 家族中大多数功能已知的成员参与很多次生代谢过程,如花香、花青素的生物合成, 尤其是许多具有临床治疗价值的生物碱的合成,如吗啡、紫杉醇、长春花碱和阿吗灵等[30]。该家族成员有很多保守区域,第1 个是位于酶中心HXXXD 结构域,对于乙酰基转移酶的反应非常重要,第2 个是位于碳端的DFGWG 结构域,是所有BAHD 成员都拥有的结构域[17]。
表达分析表明,SmVS基因的转录水平受不同组织和时间的影响,在根中表达量比叶中高,说明丹参生物碱多在根中合成。在基因工程中利用发根农杆菌诱导丹参发根,比利用根癌农杆菌诱导能产生更多的阿吗灵型生物碱vinorine;叶片中SmVS基因在白天的转录水平极低,夜间逐渐升高,21:00达到最大值,可见该生物碱主要在夜间合成。植物的生长发育受外界很多因素影响,如光照、低温、干旱等,其中光照是一种重要的环境因子,植物体内存在很多光响应的转录因子,能够感受光的刺激而诱导下游基因的表达,最终影响植物的生长发育和代谢等[32-33]。Liu 等[34]报道长春花叶中生物碱vindoline 的合成受光调控,其光应答转录因子CrGATA1 可以调控vindoline 生物合成,CrGATA1的瞬时过表达导致长春花文多灵途径基因的上调,最终增加了vindoline 的积累。因此,我们推测SmVS基因也是一个光应答基因,其转录水平受光的调控。
猜你喜欢
今日农业(2022年4期)2022-11-16
湖北农业科学(2022年11期)2022-07-18
中草药(2022年5期)2022-03-03
Digital Chinese Medicine(2021年2期)2022-01-19
Digital Chinese Medicine(2021年3期)2021-07-23
昆明医科大学学报(2021年2期)2021-03-29
烟台大学学报(自然科学与工程版)(2021年1期)2021-03-19
中文信息(2017年2期)2017-04-13
江苏农业科学(2016年11期)2017-03-21
分析化学(2014年10期)2014-10-24
