
饲料蛋白质水平对三倍体虹鳟肌肉组织特性、蛋白质周转和肌肉生长相关基因表达的影响
2022-06-10 02:31李小花田海宁李长忠孟玉琼
动物营养学报 2022年5期
李小花 田海宁 李长忠 马 睿 孟玉琼*
(1.青海大学省部共建三江源生态与高原农牧业国家重点实验室,西宁810016;2.青海大学生态环境工程学院,西宁810016)
蛋白质是水产动物饲料中必需的核心营养物质,是细胞、组织和机体的重要组成部分,为鱼体提供生长和代谢所需的能量,是决定鱼类生长的关键因素[1]。当饲料中蛋白质水平过低时,会影响鱼类的生长[2];当饲料中蛋白质水平过高时,鱼类会通过氧化脱氨基作用把过量的蛋白质分解代谢用于能量消耗,同时还会增加饲料成本,不利于鱼类的生长[3]。
鱼类的生长实质上是最大限度地促使肌肉组织的快速生长发育,达到速生快长[4]。鱼肌肉生长是一个动态过程,涉及肌纤维的增生和现有肌纤维的增大[5],增生和增大对肌肉质量增加的相对贡献可以影响肌纤维组织形态结构,其中肌纤维直径和肌纤维密度是描述肌纤维组织形态结构的2个主要特征[6-7]。同时,肌肉生长是在生长激素-胰岛素样生长因子(GH-IGFs)、肌源性调节因子(MRFs)等一系列因子的控制下进行的[8]。GH-IGFs包括生长激素受体(GHR)、胰岛素样生长因子(IGF)、胰岛素样生长因子受体(IGFR)和胰岛素生长因子结合蛋白(IGFBP)等,在鱼肌肉的生长和功能成熟中起着关键作用[9]。MRFs包括肌分化因子(Myod)、生肌因子5(Myf5)、肌原调节因子4(Mrf4)和肌细胞生成素(Myog),在成肌细胞增殖和成肌分化中起关键作用[8]。在鲑鳟鱼上研究表明,肌肉中GHR、IGF、IGFBP、Myod和Mrf4基因的表达水平会受到饲料蛋白质水平的调控[10-11]。
肌肉蛋白质是鱼肉组成的重要因素[12]。肌肉生长与肌肉蛋白质沉积有关,肌肉蛋白质沉积与肌肉蛋白质周转(包括蛋白质合成和蛋白质分解代谢过程)密切相关[13]。研究表明,胰岛素样生长因子Ⅰ(IGFⅠ)可引起磷脂酰肌醇3-激酶(PI3K)-p85活化,活化后的p85将信号传递给催化亚基PI3K-p110,从而激活PI3K,进而磷酸化蛋白激酶B(AKT-Ser437),激活雷帕霉素靶蛋白(TOR)[14],活化后的TOR可磷酸化翻译调控因子真核翻译起始因子4E结合蛋白1(4Ebp1)和核糖体蛋白S6激酶1(S6k1)来促进体内蛋白质的合成[15]。已有研究表明,饲料蛋白质可激活TOR信号通路,调节肌肉组织中TOR和4Ebp1基因的表达,最终刺激蛋白质合成[16]。另外,肌肉蛋白质沉积是蛋白质合成和蛋白质降解之间平衡的结果。蛋白质分解代谢主要由自噬/溶酶体途径、钙蛋白酶系统和泛肽降解途径三大系统来调控[17]。已有研究表明,饲料蛋白质水平会影响肌肉钙蛋白酶系统和自噬/溶酶体途径[18-19]。
虹鳟(Oncorhynchusmykiss)隶属于鲑科(Salmonidae)、大麻哈鱼属(Oncorhynchus),属于冷水性鱼类[20]。三倍体虹鳟通常是通过运用温度、压力等物化手段形成细胞内具有3套染色体的虹鳟[21],可将性腺发育的全部能量用于躯体生长,因此具有生长速度快、饲料利用高、肉品质好等优点[22],现已成为世界最为广泛的养殖鱼类之一。因此,本研究在前期三倍体虹鳟蛋白质需要量研究[12]的基础上,从肌肉组织特性、蛋白质周转、肌肉生长相关信号通路3个层次研究饲料蛋白质水平对三倍体虹鳟生长的潜在机制,为营养调控鱼类的生长提供理论基础。
1 材料与方法
1.1 试验动物、饲料及养殖管理
本试验所用三倍体虹鳟来自青海民泽龙羊峡生态水殖有限公司。按照之前报道[12]的方法配制3种等脂等能饲料,并测定各饲料水分、蛋白质、脂肪及能量水平。3种饲料蛋白质水平分别为35.4%(低蛋白质组)、45.8%(中蛋白质组)和52.4%(高蛋白质组),其中根据前期研究结果,45.8%蛋白质水平为三倍体虹鳟蛋白质需要量[12]。试验饲料组成及营养水平见表1。
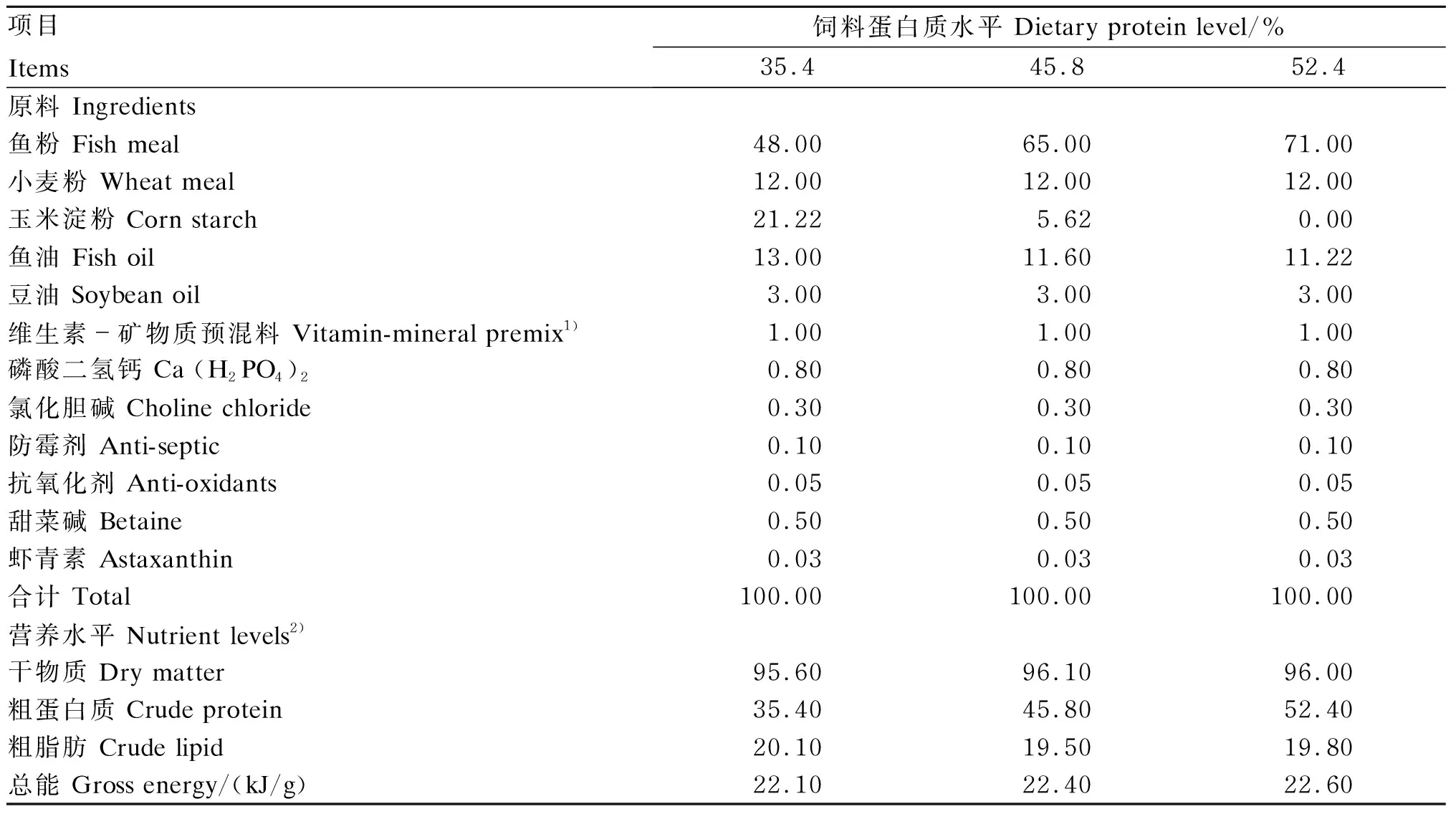
表1 试验饲料组成及营养水平(干物质基础)
将1 200尾初始体重为(232.8±0.1) g的三倍体虹鳟随机分配到12个网箱(3 m×3 m×6 m),每个网箱100尾。每个网箱随机投喂1种饲料,每种饲料有4个养殖重复(n=4),在青海省龙羊峡水库饲养80 d。在此期间,水温在8~16 ℃,溶解氧含量保持在7 mg/L以上。
1.2 样品采集
养殖试验结束后,将试验鱼禁食3 d,分别从每个网箱随机选择3尾鱼,用丁香酚(1∶10 000)麻醉(中国上海试剂公司),抽血后剖取鱼肉片,在背部取2 cm×1 cm×1 cm大小的肌肉组织块放于速冻管后放置到液氮中,每条鱼取4个备份,置于-80 ℃超低温冰箱中保存备用。
1.3 指标测定
1.3.1 肌肉组织切片和形态的测定
从-80 ℃超低温冰箱中取出肌肉样品,进行包埋(分清肌肉横纵方向)、切片(厚度是6 μm)、苏木精-伊红染色和封片(中性树脂)。用显微镜(Nikon, DS-Ri2)观察并拍照,每个肌肉组织样品采集10张100、200和400倍照片,采用图像分析软件Image-J测量肌纤维直径和肌纤维数量[5]。
1.3.2 相关基因表达的测定
采用天根RNA提取试剂盒(DP171221)提取三倍体虹鳟肌肉组织样品总RNA,分别用琼脂糖凝胶电泳和分光光度计分析总RNA的质量和浓度,质检合格后按照TaKaRa RNA反转录试剂盒[PrimeScriptTMRT Master Mix(Perfect Real Time)]反转成cDNA。参照虹鳟鱼的相关cDNA序列,设计相关基因的特异性引物(表2),引物均由上海生工生物工程技术服务有限公司合成。然后以β-肌动蛋白(β-actin)作为内参基因,每个样品进行3次技术重复,使用实时荧光定量PCR仪(LightCycler 480 Roche)进行荧光定量PCR分析,并使用2-ΔΔCt方法定量目标基因相对表达量。

表2 相关基因的引物序列
1.3.3 相关蛋白表达的测定
采用全蛋白提取试剂盒(凯基生物KGP2 500)提取三倍体虹鳟肌肉组织样品总蛋白,用酶标仪(Varioskan Lux3 020)和BCA蛋白浓度试剂盒(北京索莱宝PC0020)测定总蛋白浓度。本研究中使用的主要抗体AKT(1∶1 000;总和磷酸化Ser473)、TOR(1∶1 000;总和磷酸化Ser2 448)、4Ebp1(1∶1 000;总和磷酸化Thr37/46)、β-actin(1∶1 000)均作为内参蛋白,每个样品进行3次技术重复,使用Wes试剂盒(Protein Simple SM-W004)和Wes全自动蛋白质印迹定量分析系统(WS-2 724)进行全自动Wes分析,分析条带灰度值,以目的条带/内参条带比值表示目的蛋白相对表达量,计算磷酸化蛋白(p-)/总蛋白比值。
1.4 数据统计与分析
所有的试验数据采用SPSS 22.0软件进行单因素方差分析(one-way ANOVA),用Tukey检验进行多重比较,试验结果用平均值±标准误(mean±SE)表示。以P<0.05表示差异显著。使用Excel 2010绘图。
2 结果与分析
2.1 饲料蛋白质水平对三倍体虹鳟骨骼肌组织形态学的影响
肌纤维生长特征如表3和图1所示,结果表明,低蛋白质组的肌纤维密度和肌纤维数显著高于中、高蛋白质组(P<0.05);低蛋白质组的肌纤维直径(D)平均数、中位数、最大值和最小值显著低于其他2组(P<0.05);各组肌D≤20 μm的白色肌纤维比例无显著差异(P>0.05);低蛋白质组20 μm
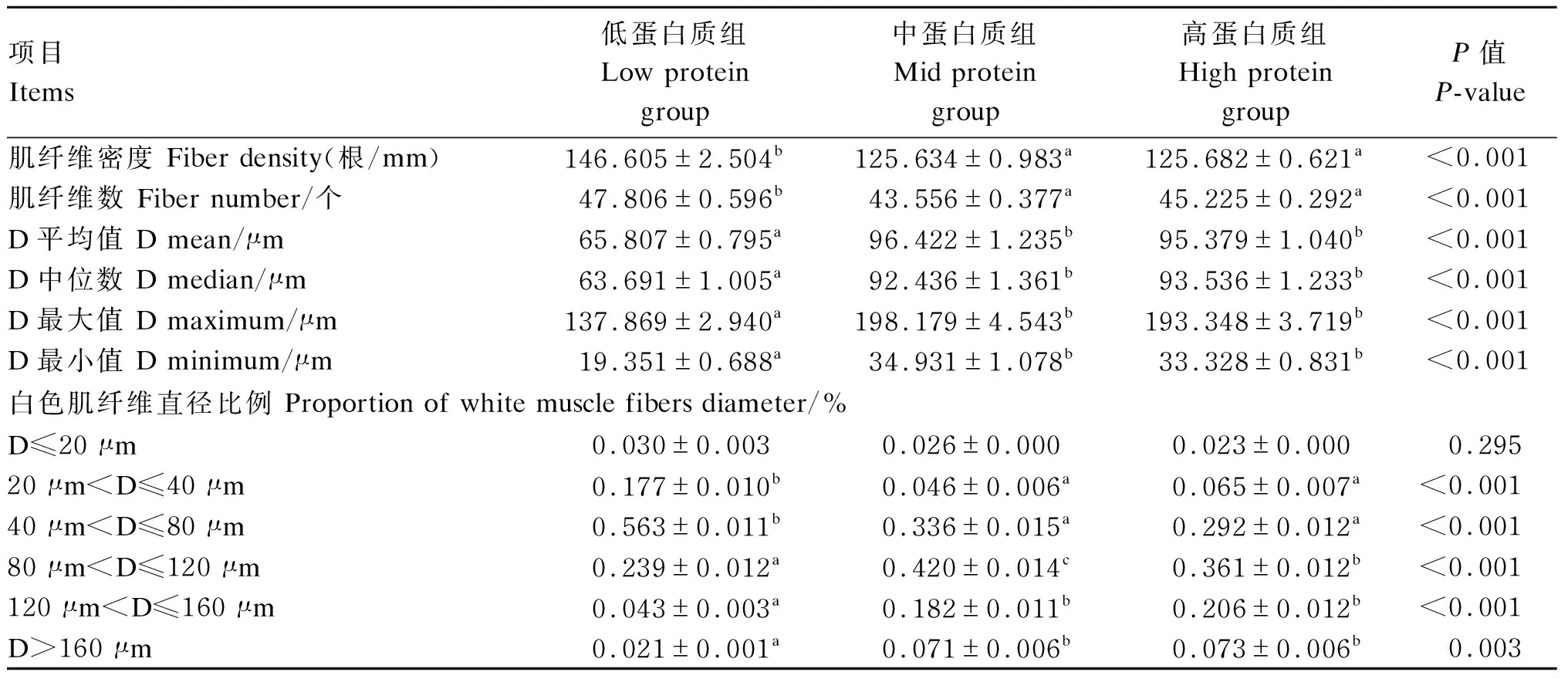
表3 饲料蛋白水平对三倍体虹鳟肌纤维形态参数的影响
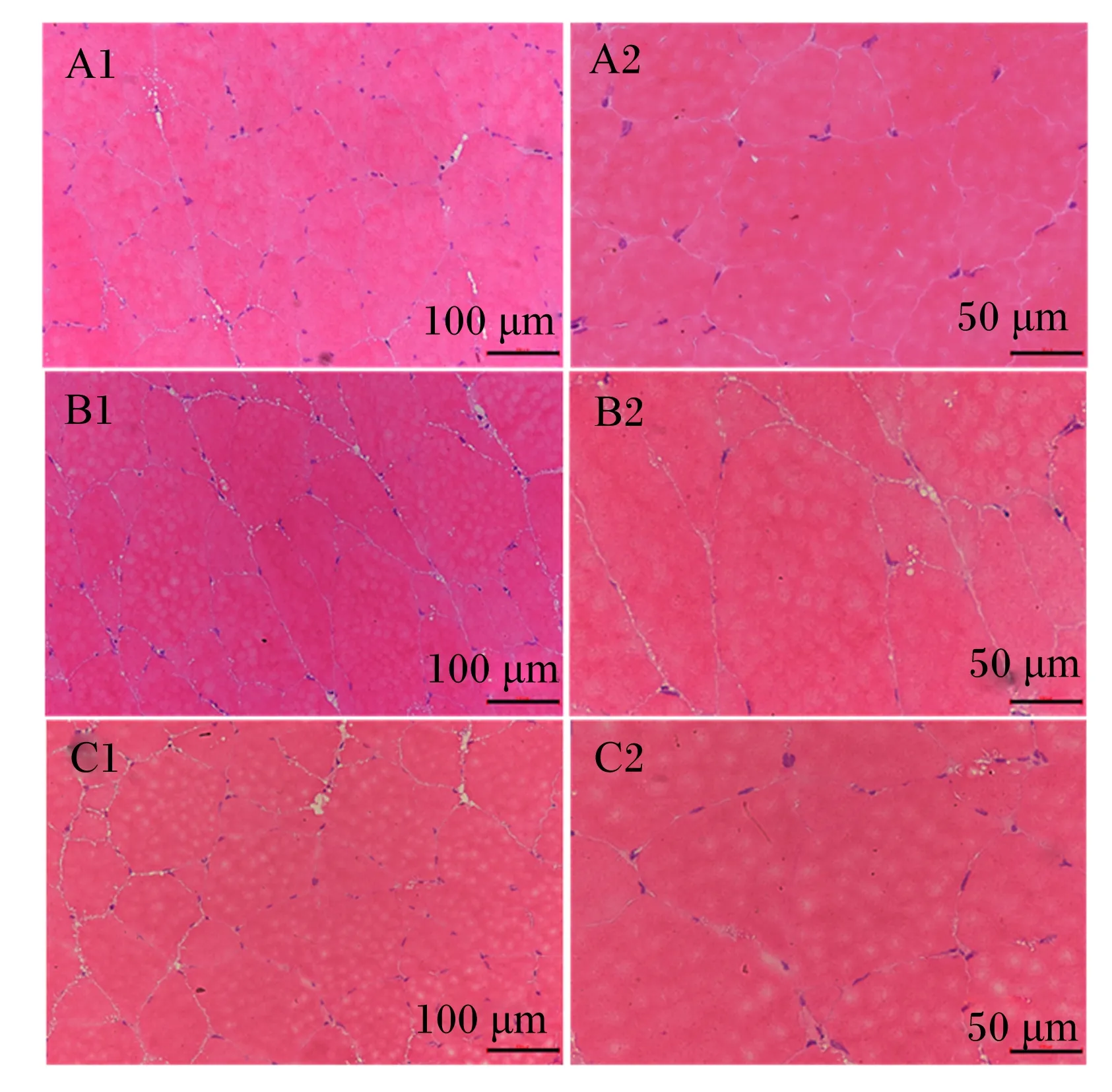
图中A、B和C代表饲料蛋白质水平(35.4%、45.8%和52.4%);1表示放大200倍;2表示放大400倍。
2.2 饲料蛋白质水平对三倍体虹鳟肌肉生长相关基因的影响
2.2.1 饲料蛋白质水平对三倍体虹鳟肌肉MRFs家族基因表达的影响
由图2可知,随着饲料蛋白质水平的增加,肌肉Myod、Myf5、Mrf4和Myog基因相对表达量呈现增加的趋势,其中低蛋白质组基因相对表达量显著低于高蛋白质组(P<0.05);而中蛋白质组和高蛋白质组之间无显著差异(P>0.05)。

Myod:肌分化因子 myodifferentiation factor;Myf5:生肌因子5 myogenic factor 5;Mrf4:肌原调节因子4 myogenic regulatory factor 4;Myog:肌细胞生成素 myogenin。数据柱标注不同小写字母表示差异显著(P<0.05)。下图同。Value columns with different small letters mean significant difference (P<0.05). The same as below.
2.2.2 饲料蛋白质水平对三倍体虹鳟肌肉GH-IGFs家族基因表达的影响
由图3可知,饲料蛋白质水平对肌肉IGFRⅡ和IGFBP1基因相对表达量无显著影响(P>0.05)。随着饲料蛋白质水平的升高,GHR、IGFⅠ和IGFRⅠ基因相对表达量呈现先上升后下降的趋势,在中蛋白质组达到最大值;而IGFⅡ基因相对表达量呈现下降的趋势,其中低蛋白质组显著高于高蛋白质组(P<0.05)。
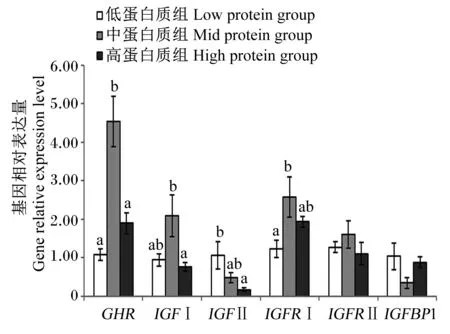
GHR:生长激素受体 growth hormone receptor;IGFⅠ:胰岛素样生长因子Ⅰ insulin-like growth factor Ⅰ;IGFⅡ:胰岛素样生长因子Ⅱ insulin-like growth factor Ⅱ;IGFRⅠ:胰岛素样生长因子受体Ⅰ insulin-like growth factor Ⅰ receptor;IGFRⅡ:胰岛素样生长因子受体Ⅱ insulin-like growth factor Ⅱ receptor;IGFBP1:胰岛素生长因子结合蛋白1 insulin-like growth factor binding protein 1。
2.3 饲料蛋白质水平对三倍体虹鳟肌肉蛋白质周转的影响
2.3.1 饲料蛋白质水平对三倍体虹鳟肌肉蛋白质合成途径的影响
由图4可知,中、高蛋白质组肌肉中p-AKT/AKT显著高于低蛋白质组(P<0.05)。随着饲料蛋白质水平的升高,肌肉中p-TOR/TOR和p-4Ebp1/4Ebp1呈现先上升后下降的趋势,在中蛋白质组达到最大值,显著高于其他2组(P<0.05)。

AKT:蛋白激酶B protein kinase B;TOR:雷帕霉素靶蛋白 target of rapamycin;4Ebp1:真核翻译起始因子4E结合蛋白1 eukaryotic translation initiation factor 4E-binding protein 1;β-actin:β-肌动蛋白。
2.3.2 饲料蛋白质水平对三倍体虹鳟肌肉蛋白质降解途径相关基因表达的影响
2.3.2.1 饲料蛋白质水平对三倍体虹鳟肌肉蛋白质自噬/溶酶体降解途径相关基因表达的影响
由图5可知,饲料蛋白质水平对肌肉微管自噬相关蛋白轻链3β(LC3β)和自噬相关蛋白4b(Atg4b)基因相对表达量无显著影响(P>0.05);中蛋白质组肌肉γ-氨基丁酸受体相关蛋白1(Gabarapl1)基因相对表达量显著高于低、高蛋白质组(P<0.05);随着饲料蛋白质水平的升高,自噬相关蛋白12l(Atg12l)基因相对表达量逐渐增加,组间差异显著(P<0.05);中、高蛋白质组肌肉中组织蛋白酶L(CathepsinsL)基因相对表达量显著高于低蛋白质组(P<0.05)。

LC3β:微管自噬相关蛋白轻链3β microtubule associated protein light chain 3 beta;Gabarapl1:γ-氨基丁酸受体相关蛋白1 gamma-aminobutyric acid receptor-associated protein 1;Atg4b:自噬相关蛋白4b autophagy-related protein 4b;Atg12l:自噬相关蛋白12l autophagy-related protein 12l;Cathepsins L:组织蛋白酶L cathepsin L。
2.3.2.2 饲料蛋白质水平对三倍体虹鳟肌肉蛋白质钙调蛋白酶降解途径相关基因表达的影响
由图6可知,饲料蛋白质水平对肌肉钙依赖性蛋白酶1(Capn1)基因相对表达量无显著影响(P>0.05);肌肉钙依赖性蛋白酶2(Capn2)基因相对表达量在高蛋白质组显著高于其他2组(P<0.05);随着饲料蛋白质水平的增加,肌肉钙蛋白酶抑制蛋白短亚型(Cast-s)和钙蛋白酶抑制蛋白长亚型(Cast-l)基因相对表达量呈现先上升后下降的趋势,其中在中蛋白质组达到最大值,显著高于其他高蛋白质组(P<0.05)。
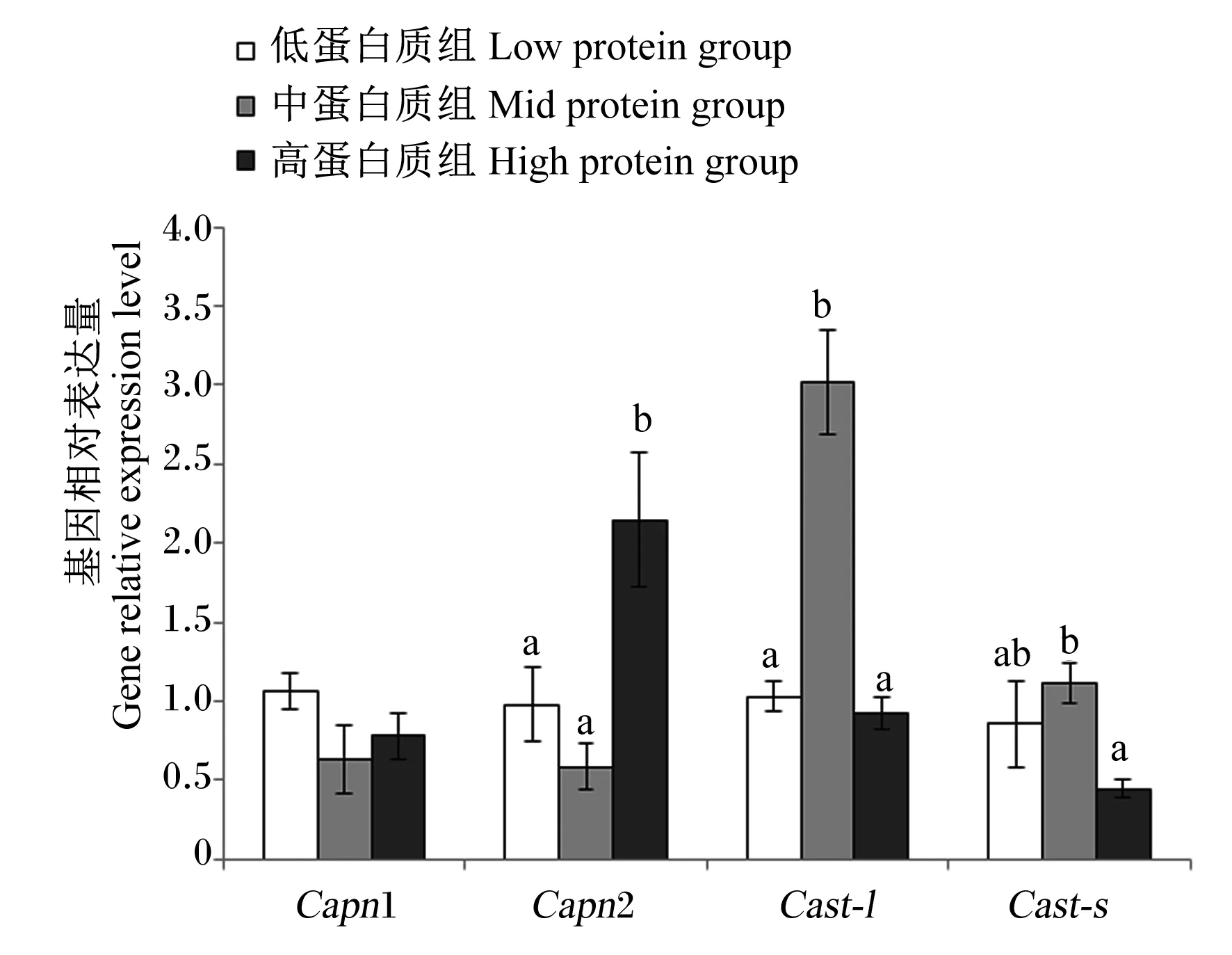
Capn1:钙依赖性蛋白酶1 calcium-dependent protease 1;Capn2:钙依赖性蛋白酶2 calcium-dependent protease 2;Cast-l:钙蛋白酶抑制蛋白长亚型 long isoforms of calpastatin; Cast-s:钙蛋白酶抑制蛋白短亚型 short isoforms of calpastatin。
2.3.2.3 饲料蛋白质水平对三倍体虹鳟肌肉蛋白质泛肽降解途径相关基因表达的影响
由图7可知,饲料蛋白质水平对肌肉特异性环指蛋白1(Murf1)和萎缩F-box蛋白32(Fbx32)基因相对表达量无显著影响(P>0.05);随着饲料蛋白质水平的增加,肌肉萎缩F-box蛋白25(Fbx25)基因相对表达量呈现先下降后上升的趋势,其中中蛋白质组显著低于低蛋白质组(P<0.05)。
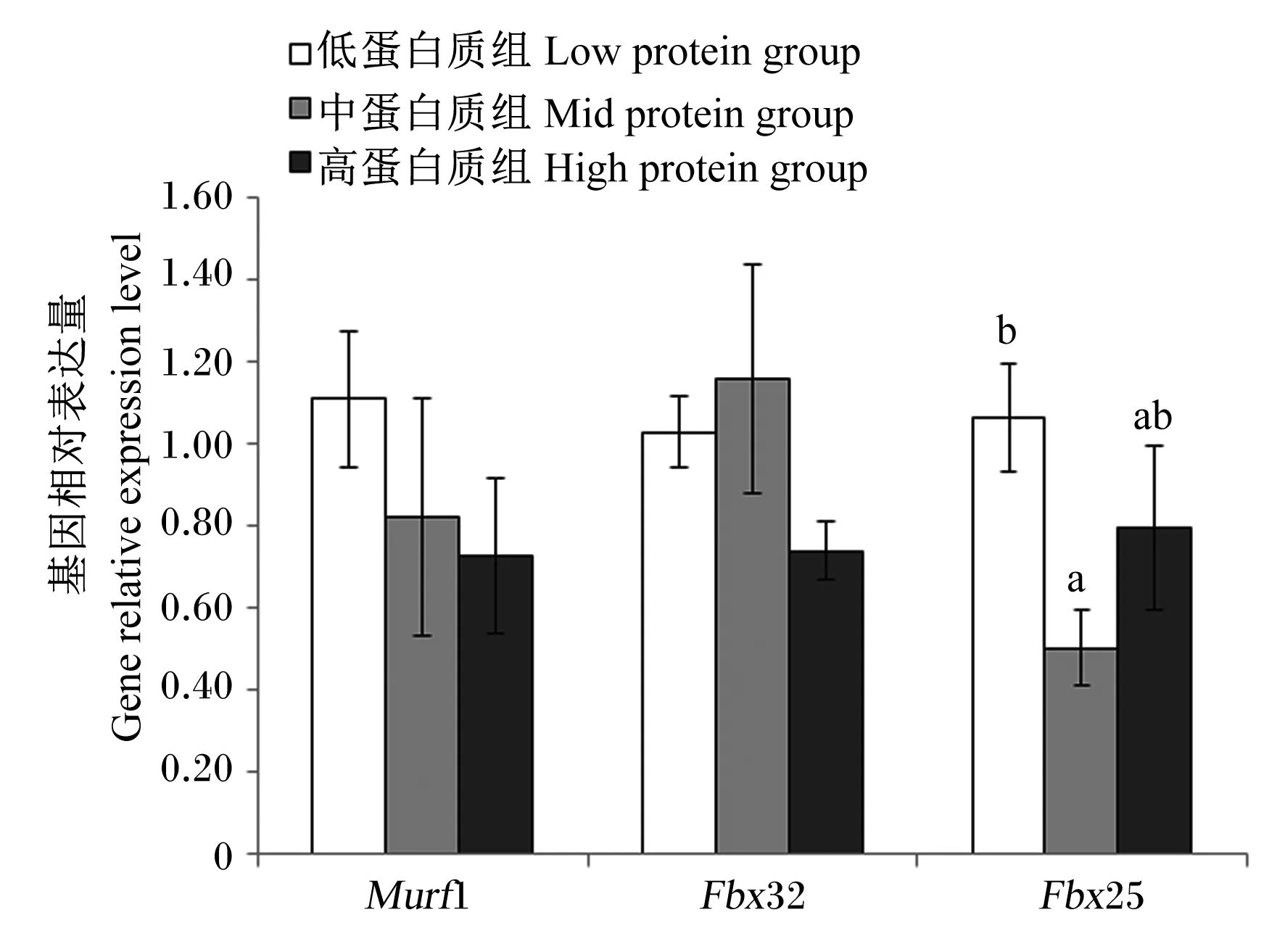
Murf1:肌肉特异性环指蛋白1 muscle specific ring finger protein 1;Fbx32:肌肉萎缩F-box蛋白32 muscle atrophy protein F-box32;Fbx25:肌肉萎缩F-box蛋白25 muscle atrophy protein F-box25。
3 讨 论
肌纤维组织形态结构包括肌纤维直径和肌纤维密度等[6-7]。前期研究表明,饲料蛋白质水平能够影响蜀宣花牛、金头鲷和大菱鲆肌纤维的密度和直径[23-26]。本研究通过对三倍体虹鳟骨骼肌的组织学观察,发现45.8%以上饲料蛋白质水平能促进三倍体虹鳟肌纤维增大,从而导致同一显微镜视野下肌纤维密度降低。这与程贵兰等[27]在麒麟鸡、Saavedra等[28]在大西洋白姑鱼(Argyrosomusregius)的研究结果一致。
鱼类肌肉生长是一个动态过程,包括2个机制:肌纤维的增大和增生[5]。肌纤维的增大是从胚胎后期开始一直到肌纤维达到功能需要的最大直径,而肌纤维的增生涉及到增加肌纤维的数量来补充新的纤维[29]。因此,肌纤维的特性可以作为衡量细胞增大或增生的指标。骨骼肌生长发育过程中,如果存在大量D<20 μm的肌纤维,代表肌纤维数增加的生长方式较活跃[30-31],如果存在D>120 μm的肌纤维,则代表肌肉对肌纤维的募集能力消失,肌肉纤维生长主要以增大为主,肌纤维数不再增加[32-33]。本研究结果表明,饲料蛋白质水平对三倍体虹鳟肌纤维增生无显著影响,而较高饲料蛋白质水平(45.8%以上)能促进鱼体肌纤维增大。
鱼类骨骼肌的生长受一系列信号通路的调控,如GH-IGFs和MRFs[34-35]。前期在斑马鱼、金头鲷和虹鳟研究表明,IGFs通过调节肌肉蛋白质沉积、肌细胞增生和肥大在促进肌肉生长中发挥重要作用[36-37]。GH-IGFs生长轴是鱼类生长发育的重要调控系统,该系统包括GH、GHR、IGF、IGFBP和IGFR等[38-40]。一般认为,该系统的调控机理为:由垂体分泌的GH通过与靶细胞GHR结合,启动细胞内的信号传导机制,促进IGF的表达与合成;IGF通过血液循环到达机体的局部组织,作为胞外配体通过与细胞膜上的受体结合,促进组织细胞分裂、分化和生长[41]。马骏等[10]在大西洋鲑(Salmosalar)中研究发现,饲料蛋白质水平可显著影响肌肉中GHR和IGFⅠ基因表达水平。与其结果类似,本研究也发现肌肉GHR、IGFⅠ和IGFRⅠ基因相对表达量在45.8%饲料蛋白质水平组最高,这也与本课题组之前研究结果一致:随着饲料蛋白质水平从31.6%增加到45.8%,三倍体虹鳟生长速度明显提高;而当饲料蛋白质水平大于45.8%以后生长速度不再增加[12]。然而在本研究中IGFⅡ基因相对表达量随着饲料蛋白质水平的增加呈现显著下降的趋势,可能与受到IGFⅠ的负反馈调节有关,类似的结果也发现于Sterle等[42]在猪上的研究。肌肉中特异性表达的MRFs家族成员有4个:Myod、Myog、Myf5和Mrf4。Myod和Myf5是肌卫星细胞分化发育成为肌细胞必需的,而Myog和Mrf4则通过招募肌球蛋白等结构蛋白在肌纤维分化中发挥作用。饮食中常量营养物质的变化,包括蛋白质来源和氨基酸补充,会影响MRFs的水平[43]。在虹鳟上的前期研究表明,饲料蛋白质水平或氨基酸水平与肌肉MRFs家族基因表达成正相关[11,44]。类似的结果在本研究中同样发现。然而在本研究中,饲料蛋白质水平促进了MRFs家族基因的表达,但没有促进肌纤维增生。这可能的原因与IGF能诱导MRFs相关基因表达有关[45],潜在机制仍需进一步研究。
肌肉蛋白质沉积与肌肉蛋白质周转(包括蛋白质合成和蛋白质分解代谢过程)密切相关[13]。研究表明,在机体内蛋白质合成代谢依赖于TOR途径[46]。TOR通路可汇聚和整合多种细胞刺激信号(如生长因子、营养因子、能量等)来调控蛋白质代谢及许多重要生理过程[47]。IGFⅠ可以通过IGFⅠR-PI3K-AKT途径促进肌肉生长[48]。原因与该途径磷酸化TOR后使其活化,进而磷酸化翻译调控因子4Ebp1和S6k1来促进体内蛋白质的合成有关[15]。已有研究表明,饲料蛋白质能够提高TOR信号通路相关基因的表达水平,促进蛋白质的合成代谢[49]。在本研究中,当饲料蛋白质水平从35.4%增加到45.8%时,AKT、TOR和4Ebp1的磷酸化水平显著提高,这与Li等[50]在猪背肌上的研究结果一致。然而,当饲料蛋白质水平从45.8%增加到52.4%,TOR和4Ebp1的磷酸化水平降低,这与过高饲料蛋白质水平抑制GH-IGFs家族相关基因表达结果一致,使得肌肉乃至鱼体生长不再进一步增加。蛋白质分解代谢主要由自噬/溶酶体途径、钙蛋白酶系统和泛肽降解途径三大系统来调控[17]。自噬/溶酶体途径是一个进化的保守过程,可将降解长寿命的蛋白质和消除多余或受损的细胞结构[51]。自噬过程需要大量自噬相关因子(LC3β、Gabarapl1、Atg12l、Atg4b)参与,被证明在控制骨骼肌质量方面发挥重要作用[52]。在虹鳟中研究发现,肌肉蛋白质自噬降解相关基因(LC3β、Gabarapl1、Atg12l、Atg4b)受饲料蛋白质和碳水化合物水平的影响[18-19]。在本研究中,45.8%及以上饲料蛋白质水平能上调Gabarapl1、Atg12l和CathepsinL基因相对表达量,促进三倍体虹鳟肌肉蛋白质降解。钙蛋白酶系统是一种广泛分布于哺乳动物组织中高度复杂和高度调控的蛋白质降解系统[53]。钙蛋白酶家族是一类细胞内的半胱氨酸蛋白水解酶,共有16个成员:钙蛋白酶1-钙蛋白酶15(CAPN1-CAPN15)和钙蛋白酶调节亚基2(CAPNS2)。骨骼肌中主要表达Capn1、Capn2及内源性钙蛋白酶抑制剂(Cast)。饲料蛋白质水平与肌肉钙蛋白酶降解途径在陆生动物中的研究结果存在争议,王耀梅等[54]在绵羊上的研究发现,一些肉品质相关基因表达量随饲料蛋白质水平的升高分别升高和降低;而马丽娜等[55]在滩羊肌肉组织中发现,Capn1和Capn2基因在高饲料蛋白质水平组中表达上调。本研究中,52.4%饲料蛋白质水平显著上调肌肉Capn2基因相对表达量,进而激活钙蛋白酶降解途径,可能与过高蛋白质水平抑制了Cast有关。泛肽降解途径是一种具有选择性的和高度特异性的蛋白质降解途径,实现这一目标主要依赖于能特异性识别特定蛋白质底物的连接酶E3[56]。目前骨骼肌所特有的E3主要有:肌肉萎缩F-box蛋白(MAFbx或Fbx)和Murf1。在本研究中,虽然肌肉Murf1基因相对表达量无显著变化,但Fbx25基因相对表达量在低蛋白质组最高,Li等[50]研究发现,能量状态限制可以通过增加E3连接酶(Murf1和MAFbx)的基因表达水平促进蛋白降解。Baptista等[57]也表明增加氨基酸水平可以抑制Fbx的表达。鉴于此,当饲料蛋白质水平无法满足鱼体需要时,鱼体启动泛肽降解途径选择性得降解蛋白质提供能量。
4 结 论
① 当饲料蛋白质水平达到三倍体虹鳟蛋白质需要量45.8%时,能促进肌纤维的增大,对肌纤维增生无显著影响;进一步提高饲料蛋白质水平对肌纤维生长无显著影响。
② 饲料蛋白质水平通过调控MRFs和GH-IGFs通路进而调控肌纤维增大。
③ 当饲料蛋白质水平增加到45.8%时,可促进肌肉蛋白质合成代谢;然而进一步提高饲料蛋白质水平会抑制该作用。
④ 肌肉蛋白质各分解途径在不同饲料蛋白质水平时存在差异。当投喂过低蛋白质水平(35.4%)饲料时,三倍体虹鳟会启动泛肽降解途径选择性得降解蛋白质提供能量;当投喂的饲料其蛋白质水平超过45.8%时,肌肉蛋白质降解以自噬溶酶体途径为主;当投喂过高蛋白质水平(52.4%)饲料,鱼体还会启动钙调蛋白酶降解途径。
猜你喜欢
浙江农业学报(2022年1期)2022-11-07
肝博士(2022年3期)2022-06-30
江西水产科技(2022年2期)2022-05-17
中国饲料(2022年5期)2022-04-26
海外星云(2021年9期)2021-10-14
食品安全导刊·中旬刊(2020年2期)2020-06-01
灌篮(2020年36期)2020-05-16
文苑(2018年22期)2018-11-19
大众健康(2015年12期)2015-09-10
