
木霉T6 生物菌肥对苹果砧木M9T337幼苗生长的促进作用*
2022-06-09 01:08赵国康张树武徐秉良石海春
中国果树 2022年5期
杨 瑞,赵国康,张树武,徐秉良,石海春
(甘肃农业大学植物保护学院,甘肃省农作物病虫害生物防治工程实验室,兰州 730070)
木霉菌是一种广泛存在的生防真菌,不仅具有抗菌作用而且对植物生长有促进作用[1]。木霉生物菌肥具有生产成本低、安全无污染、改良土壤、减轻作物病虫害、增加作物产量等优点。此外,木霉生物菌肥能促进甜瓜、春油菜等作物的生长,能增加青椒、黄瓜的产量和改善其品质[2-5]。曾庆才[6]研究发现,适量的哈茨木霉生物菌肥对茄子和苦瓜的生长有明显的促进作用。木霉菌肥不仅广泛应用于各种蔬菜上,同时也被应用于中药材种植中,叶冰竹等[7]研究发现,深绿木霉D16 生物菌肥能促进丹参苗的生长和有效成分的积累。除此之外,木霉菌肥还可以用于植物病原线虫的防治,王海明[8]研究结果表明,生物炭和麦麸木霉菌肥能够显著降低南方番茄地根结线虫的种群数量并增加番茄的产量。
研究发现,生物菌肥对苹果树幼苗的生长具有明显的促进作用[9-10]。此外,生物菌肥可以显著提高果树果实的含糖量、着色系数[11]。但木霉生物菌肥在苹果树上的应用尚未见报道,因此,本试验基于形态指标和生理生化指标两方面研究了木霉T6 生物菌肥对盆栽苹果砧木幼苗生长及其抗氧化酶活性的影响,确定其最佳的施肥量。研究结果对合理应用木霉生物菌肥、有效促进苹果砧木幼苗生长具有重要意义,以期为大规模应用提供试验数据。
1 材料与方法
1.1 试验材料
木霉T6 生物菌肥由甘肃农业大学植物保护学院植物病理学实验室研制。苹果砧木M9T337 幼苗,购自山东惠农园艺科技有限公司。营养土为购自桃海花卉市场的育苗基质和绿能瑞奇营养基质。
1.2 试验方法
1.2.1 试验处理
选取生长健壮、长势一致(株高约7 cm,茎粗约2.5 cm)、苗龄30 d 的M9T337 幼苗60 株,2019年12 月30 日移栽在花盆(10 cm×10 cm)中。盆土为混入木霉T6 生物菌肥的灭菌土,每盆土重500 g,移栽2 株。移栽后的幼苗置于气温为25 ℃、空气相对湿度为65%的环境下,每隔3 d 浇1 次水。
试验共设5 个处理:木霉T6 生物菌肥施肥量分别为土壤质量的0.5%(T1)、1%(T2)、2%(T3)、3%(T4),对照(CK)不施菌肥。每2 株为1 次重复,重复6 次。
1.2.2 M9T337 幼苗生长指标的调查
M9T337 幼苗移栽3 个月后,测定其鲜重、根重、地径、株高和根长,每个处理重复3 次。
1.2.3 M9T337 幼苗生理生化指标的测定及其方法
M9T337 幼苗移栽6 个月后,分别测定幼苗叶片和根系超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)活性和可溶性蛋白含量,每个处理重复3 次。
结构的主要荷载有:(1)结构自重;(2)活荷载;(3)整体堆煤荷载;(4)局部堆煤荷载;(5)地震作用;(6)温度作用;(7)风荷载。主要工况组合为:(1)整体堆煤荷载组合;(2)局部堆煤荷载荷载;(3)地震作用组合;(4)温度作用组合等[1]。
(1)SOD 活性的测定。参考张建文[12]的方法,采用NBT 核黄素比色法测定。称取叶片和根各0.5 g,分别剪碎后,各置于预冷的研钵中,加入2 mL预冷的提取介质,在冰浴条件下研磨成匀浆,将匀浆全部转入到离心管中,最终体积为10 mL,并于4 ℃下12 000 r/min 下离心30 min,获得SOD 粗提液。然后,吸取20 μL 酶液,加入3 mL 反应液,4 000 lx 照光30 min,空白置暗处,对照(CK)与酶液同置于4 000 lx 条件下照光30 min,遮光保存,以空白调零,560 nm 比色。
SOD 总活性(吸光度/g)=(ACK-AE)×V/(FW×0.5×ACK)
公式中,ACK 为照光对照管的吸光度,AE 为样品管的吸光度,V 为样品提取液总体积(mL),FW 为植物样品鲜重(g)。SOD 总活性单位为NBT光还原50%。
(2)POD 活性的测定。采用愈创木酚法测定。称取叶片和根各1 g,分别剪碎后,各放入预冷的研钵中,加入5 mL pH 值6.0 磷酸缓冲液研磨成匀浆,将其移入100 mL 容量瓶中,经磷酸缓冲液将残渣冲洗入容量瓶并定容。混匀后取一定量放入离心管中,并于4 ℃下4 000 r/min 离心10 min,获得POD粗提液。取20 μL 酶液与3 mL 反应液置于比色皿中,在470 nm 比色,每隔1 min 读数1 次,共读数3 次,并以每分钟吸光度变化值表示酶活力的大小。
POD 总活性[ΔA470/(min·g)]=ΔA470×V/Va/FW
公式中,ΔA470 为在波长470 nm 下的吸光值的变化,V 为提取酶液总体积(mL),Va 为测定时所用酶液体积(mL),FW 为植物样品鲜重(g)。
(3)CAT 活性的测定。采用钼酸铵比色法测定。称取叶片和根各0.5 g,各加入pH 值7.8 的磷酸缓冲液及少量石英砂,冰浴研磨成匀浆,分别转移至25 mL 容量瓶中定容,并于4 ℃下4 000 r/min离心15 min,获得CAT 粗提液。然后,将0.1 mL(或50 μL)酶液与2.5 mL 反应液置于比色杯中,240 nm 比色,每隔1 min 读数1 次,共读数3 次。
CAT 总活性[ΔA240/(min·g)]=ΔA240×V/Va/FW
(4)可溶性蛋白含量的测定。采用考马斯亮蓝法测定。称取叶片和根各2 g,分别剪碎,各放入研钵中,加5 倍体积预冷的磷酸缓冲液,研磨成匀浆,全部转移至离心管中,于4 ℃下13 000g离心15 min,获得可溶性蛋白提取液,吸取样品提取液0.1 mL,加入4.9 mL 考马斯亮蓝G-250 溶液,充分混匀并静置2 min 后在595 nm 波长下比色。
可溶性蛋白含量(mg/g)=C×V/Va/FW
公式中,C 为根据标准曲线得到的样品管中蛋白质含量(μg),V 为提取液总体积(mL),Va为测定所用酶液体积(mL),FW 为植物样品鲜重(g)。
1.3 数据处理
利用Microsoft Excel 2010 软件进行图表制作,利用SPSS 22.0 软件中的方差分析(ANOVA)多重比较(Duncan’s)对数据进行分析。
2 结果与分析
2.1 木霉T6 生物菌肥施入量对M9T337 幼苗生长的影响
施用不同量木霉T6生物菌肥对M9T337幼苗株高、鲜重、地径、根长和叶片数均有促进作用。处理3 个月后,与对照相比,木霉T6 生物菌肥施入量为土壤质量的2%(T3)和3%(T4)时,对苹果砧木M9T337 幼苗的生长促进作用最显著。木霉T6生物菌肥施入量为2%(T3)时,幼苗株高、鲜重、地径、根长和叶片数的增长率分别为38.01%、143.40%、30.21%、85.43%、206.38%;施入量为3%(T4)时,幼苗株高、鲜重、地径、根长和叶片数的增长率分别为59.45%、120.75%、46.84%、83.27%、162.66%(表1)。

表1 施不同量木霉T6 生物菌肥M9T337 幼苗的生长指标
2.2 木霉T6 生物菌肥施入量对M9T337 幼苗SOD活性的影响
从图1 可以看出,与对照相比,不同木霉T6生物菌肥施入量均能显著提高M9T337 幼苗叶片和根系的SOD 活性。其中,就叶片而言,木霉T6 生物菌肥施入量为土壤质量的2%(T3)和3%(T4)时,SOD 活性最高,显著高于其他2 个木霉T6 生物菌肥和对照;就根系而言,SOD 活性随木霉T6生物菌肥施入量的增加而提高,木霉T6 生物菌肥施入量为土壤质量的3%(T4)时最高,对照最低,各处理间SOD 活性差异均显著。

图1 施入不同量木霉T6 生物菌肥M9T337 幼苗叶片和根系的SOD 活性
2.3 木霉T6 生物菌肥施入量对M9T337 幼苗POD活性的影响
从图2 可以看出,M9T337 叶片和根系POD 活性均随木霉T6 生物菌肥施入量的增加而提高,均以木霉T6 生物菌肥施入量为土壤质量的3%(T4)处理最高,对照最低;且各处理间叶片和根系POD活性差异均显著。
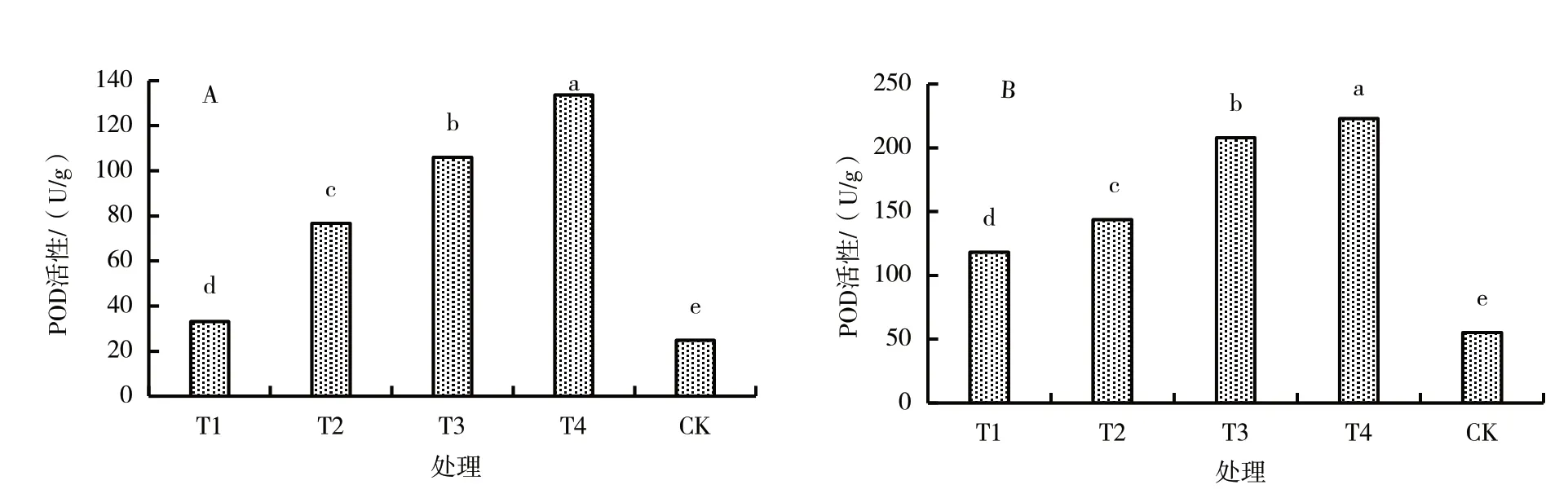
图2 施入不同量木霉T6 生物菌肥M9T337 幼苗叶片和根系的POD 活性
2.4 木霉T6 生物菌肥施入量对M9T337 幼苗CAT活性的影响
从图3 可以看出,M9T337 叶片和根系CAT 活性均随木霉T6 生物菌肥施入量的增加而提高,均以木霉T6 生物菌肥施入量为土壤质量的3%(T4)处理最高,对照最低;各处理间叶片和根系CAT 活性差异均显著。
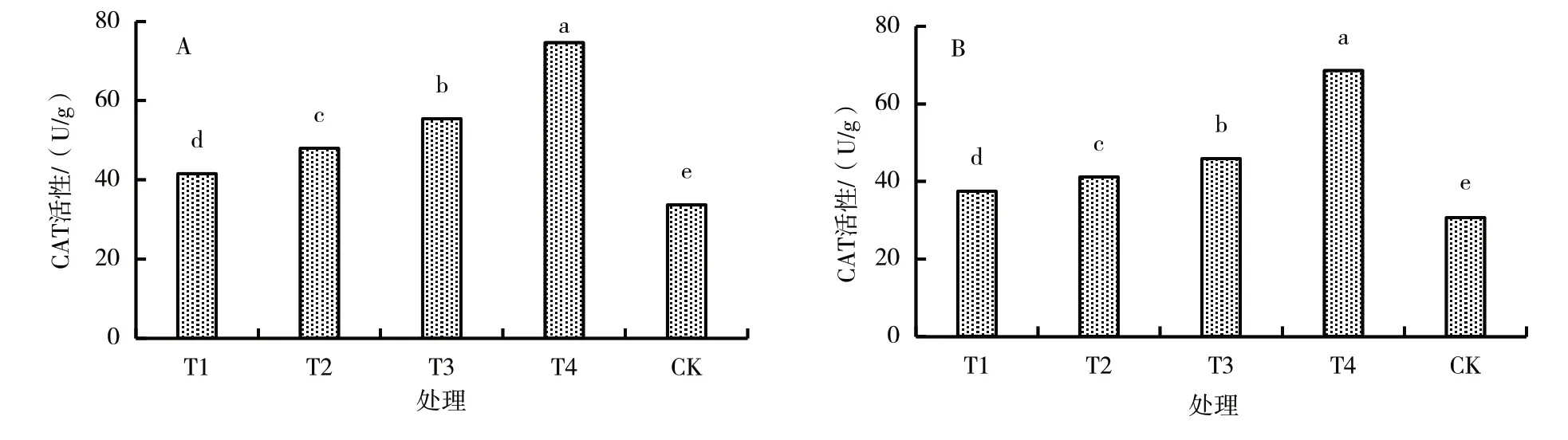
图3 施入不同量木霉T6 生物菌肥M9T337 幼苗叶片和根系的CAT 活性
2.5 木霉T6 生物菌肥施入量对M9T337 幼苗可溶性蛋白含量的影响
从图4 可以看出,M9T337 叶片和根系可溶性蛋白含量均随木霉T6 生物菌肥施入量的增加而提高,均以木霉T6 生物菌肥施入量为土壤质量的3%(T4)处理最高,对照最低;除T1 和T2 处理间根系可溶性蛋白含量差异不显著外,其他各处理间叶片和根系可溶性蛋白含量差异均显著。

图4 施入不同量木霉T6 生物菌肥M9T337 幼苗叶片和根系的可溶性蛋白含量
3 讨论与结论
土壤微生物与土壤肥力关系密切,对维持土壤的生态平衡有着重要作用[13]。吴晓娴[14]研究表明,施入生物菌肥后,果园土壤中有益微生物增多,苹果生长及产量显著增加;王洋娟[9]发现,施用微生物菌肥可以明显促进苹果幼苗的生长,其株高、茎粗、枝长等均有不同程度的增加。本研究结果表明,施用不同量木霉T6 生物菌肥均能显著促进苹果砧木M9T337 幼苗的生长,当施肥量为土壤质量的3%时M9T337 幼苗株高、鲜重、地径、根长和叶片数均最大。陈伟等[15]研究发现,木美土里生物菌肥可以显著改善苹果树的根系生长及枝条生长。本试验结果与前人研究结果基本一致。
SOD、CAT 和POD 是生物演化过程中建立起来的生防生化系统关键酶,均能使植物体内的活性氧及过氧化氢等物质转变为活性较低的物质,降低或消除它们对植物膜脂的攻击能力,使膜脂不被氧化而受到保护[16-18]。植物体内可溶性蛋白含量的变化可以反映植物体总代谢情况,植物体内的可溶性蛋白大多是各种代谢所需的酶,也是重要的营养物质以及渗透调节物质,对细胞的生物膜和生命物质起到保护作用[19-20]。
曾庆宾等[1]研究了苗床添加木霉菌肥对烟苗生长和相关抗逆酶活性的影响,表明苗床添加中高浓度木霉菌肥可提高红花大金元和云烟85 幼苗叶片和根系的CAT、POD 活性,促进烟草苗的生长和养分的吸收。小梅[21]研究发现,施用菌肥能显著提高降香黄檀幼苗SOD、CAT 和POD 等保护酶的活性。本研究表明,不同木霉T6 生物菌肥施肥量处理的M9T337 幼苗叶片和根系SOD、POD、CAT 活性均显著高于对照,且在施肥量为土壤质量的3%时达到最高值,表明木霉T6 生物菌肥能提高植物的抗逆性,促进苹果砧木幼苗的生长,这与前期研究结果基本一致。同时,本研究中发现3%木霉T6 生物菌肥处理后,苹果砧木幼苗叶片和根系的可溶性蛋白含量均显著上升。相关研究表明[22],人参叶片中可溶性蛋白含量随着生物菌肥浓度的增加呈显著增加的趋势,当施肥量为80 g/m2时达到最大值。
小梅[21]研究表明,降香黄檀幼苗保护酶活性和可溶性蛋白含量随着水溶性复合微生物菌肥施用量的增加呈现出先上升后下降的趋势,而本研究发现苹果砧木幼苗保护酶活性随着菌肥施用量的增加呈现逐渐上升的趋势,这可能与不同植物、不同处理和不同菌肥有关。木霉T6 生物菌肥能够显著提高M9T337 幼苗的株高、鲜重、地径、根长和叶片数,提高苹果砧木幼苗的抗氧化酶活性和可溶性蛋白的含量,进而促进苹果砧木幼苗的生长。本试验仅在盆栽条件下研究了木霉T6 生物菌肥对苹果砧木幼苗生长的影响,其实际应用效果还需进一步通过田间试验进行验证。
猜你喜欢
今日农业(2022年15期)2022-09-20
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
新农村(2018年24期)2018-09-14
科学种养(2018年1期)2018-01-20
农家科技下旬刊(2017年10期)2017-12-06
江苏农业科学(2017年19期)2017-11-22
新农业(2017年3期)2017-05-17
农家顾问(2016年7期)2016-05-14
农家科技(2016年1期)2016-03-03
