
草地贪夜蛾雄蛾交配状态的判断
2022-06-07 09:20:38冯行利曹婷婷杨凤丽吴慧明饶琼郑永利
植物保护 2022年3期
冯行利 曹婷婷 杨凤丽 吴慧明 饶琼 郑永利
摘要 草地贪夜蛾Spodoptera frugiperda是近年入侵我国的重大迁飞性害虫。本研究通过解剖观察不同日龄和交配状态的草地贪夜蛾雄蛾内生殖系统,确定其内含物分级标准,并以此为依据分析性诱剂田间诱捕雄蛾的交配状态。结果表明草地贪夜蛾精巢体积随日龄增加而减小,發育后期已交配的精巢皱缩扁平,日龄对未交配雄蛾单射精管非角质化区(PS)末端内含物无显著影响。根据PS末端内含物的颜色和紧密程度将雄蛾交配状态分为5级(Ⅰ~Ⅴ级)。未交配雄蛾PS末端内含物为褐色且均匀(Ⅴ级),交配后0 h呈透明(Ⅰ级),之后内含物会逐渐回填(Ⅱ~Ⅳ)。田间诱捕雄蛾共921头,其中647头已交配(Ⅰ~Ⅳ级)。不同性诱剂诱捕雄蛾的交配状态无显著差异。本研究结果为草地贪夜蛾的预测预报和性诱剂防控实际控害效果评价提供参考。
关键词 草地贪夜蛾;雄蛾;生殖结构;交配状态;精巢;性诱剂
中图分类号: S436.3
文献标识码: A
DOI: 10.16688/j.zwbh.2021118
Abstract The fall armyworm, Spodoptera frugiperda is an important invasive migratory pest in China. The internal reproductive system of male moths at different day-age and mating status were observed by dissection, and the classification standard was established. Male moths trapped by sex pheromone in field was dissected to analyze the mated status according to the classification standard above. The results showed that testis volume was decreased with the increase of day-age, and the mated testis became shrank and flattened in the later stage of development. But there was no change in the genital contents in primary segment (PS) of single ductus ejaculatorius with the increase of day-age. According to color and compactness of the genital contents in the end of PS, mated status were classified into five grades (Ⅰ-Ⅴ). The genital contents of unmated male adults were brown or dark brown (grade Ⅴ), with high uniformity. The genital contents were completely excreted and became transparent after mating (grade Ⅰ), and then it began to backfill (grade Ⅱ to grade Ⅳ). There were 921 male moths were trapped in the field by sex pheromone traps, among which the number of mated males (grade Ⅰ to grade Ⅳ) was 647 individuals. There were no significant differences in mated status of the male moth traped by different sex pheromone traps. This study provides a reference for the prediction of S.frugiperda and evaluation of the actual control effect of sex pheromone traps.
Key words Spodoptera frugiperda;male moth;reproductive structure;copulation status;testis;sex pheromone
草地贪夜蛾Spodoptera frugiperda (J.E.Smith), 又称“秋黏虫”(fall armyworm),属鳞翅目夜蛾科,具有较强的迁飞性、暴食性与繁殖能力,是我国重要入侵性害虫[1-3]。2020年全国各地加强了预警防控措施,但草地贪夜蛾发生量仍居高不下,给广大玉米种植者带来严重损失。目前草地贪夜蛾的防控主要依赖于化学防治,但化学药剂的大量使用使得草地贪夜蛾种群抗性风险增高,同时给环境安全和人类健康带来隐患。昆虫性诱剂因其无污染,对环境友好等特点,在害虫预测预报和防治中得到广泛应用,可通过田间诱捕量来判断其诱捕效果,此外性诱剂诱捕雄蛾的交配率也是评判其防治效果的重要参数[4],田间诱集到未交配雄蛾,可有效降低雌蛾交配率,从而降低下一代发生虫量[5-6]。
鳞翅目昆虫精巢是精子发育和储存的重要器官,在交配过程中,精子从精巢排出,附腺分泌的精液会保护精子通过单射精管进入雌蛾体内,而单射精管非角质化区(primary segment, PS)的分泌物质主要参与精包的形成[7-8],利用单射精管的填充情况和内含物颜色、单射精管厚肌区的形态等可判定雄蛾的交配状态[4, 9-10]。目前有关草地贪夜蛾交配状态判断的研究鲜有报道,亦无判断交配状态的相关标准。本研究通过解剖观察草地贪夜蛾雄蛾内生殖结构,探索其交配状态的判断方法,结合田间诱捕试验调查雄蛾的交配率,为草地贪夜蛾性信息素田间诱捕效果的评判提供参考。
1 材料与方法
1. 1 试虫
供试草地贪夜蛾2020年5月采集于杭州市余杭区余杭街道木桥村玉米种植区(120.27°E, 30.42°N)。将采集到的草地贪夜蛾5~6龄老熟幼虫置于养虫室饲养,温度(28±1)℃,湿度(70±5)%,光周期L∥D=16 h∥8 h。幼虫在3龄前喂食新鲜玉米嫩叶,3龄末期用六孔板单头饲养,喂食人工饲料[11]。幼虫化蛹后鉴别雌雄,分别放入养虫盒内,置于黑暗条件,待羽化。每日9:00收集羽化成虫,鉴别雌雄后分别放入60 cm×60 cm×60 cm养虫笼内饲养,成虫需每日傍晚喂食10%蜂蜜水[11-12]。
1.2 内生殖器解剖方法
参考和伟等[13]对草地贪夜蛾的解剖方法。
雄蛾:将草地贪夜蛾雄蛾用解剖剪从胸腹部连接处剪断,取腹部置于培养皿中,利用镊子小心地将各节的背板和腹板全部去掉,用生理盐水冲洗干净并剔除消化系统和脂肪粒等,最终只保留干净完整的雄蛾内生殖结构进行拍照测量。
雄蛹:首先将雄蛹头胸部剪除,再从腹部将蛹壳剪开,用生理盐水配合解剖镊子多次冲洗腹腔,完全清除腹腔内多余的组织和脂肪粒,保留干净完整的精巢进行拍照测量。
1.3 草地贪夜蛾雄蛾不同日龄和不同交配状态内生殖系统比较
分别收集0~6日龄未交配雄蛾,立即解剖观察。将初羽化雌雄蛾单头配对置于养虫盒(15 cm×8 cm)中,使用摄像机SONY SF-32C4拍摄确定其交配时间,将交配后的雄蛾单独饲养。收集已交配0、1、2、4、6 d的雄蛾解剖,方法同上。
1.4 草地贪夜蛾精巢体积与日龄相关性比较
蛹期选取并解剖化蛹当天至其羽化不同日龄体重体长相近的雄蛹;成虫期选取并解剖1~6日龄且体态相近并具有较强活力的雄蛾;解剖方法同上;使用体式显微镜Nikon SMZ25对精巢进行测量和拍照;精巢体积的测量参考冯波等对二化螟精巢测量的方法[14],分别测量精巢赤道半径a,b(沿着x轴和y轴)和极半径c的长度,按椭圆体积公式V=4πabc/3计算草地贪夜蛾精巢体积。
1.5 田间诱捕草地贪夜蛾雄蛾交配率调查
2020年10月在杭州市余杭玉米种植区利用性诱剂进行诱捕试验,选择生长正常、长势均匀、两周内未使用杀虫剂的田块,选择市面上常见的3种诱芯,每种诱芯设3个重复,每个重复设置1个诱捕器并配置1枚诱芯;每个诱捕器间隔30 m以上,距离地面1.5 m左右。3种诱芯生产单位分别为北京中捷四方生物科技股份有限公司(ZJSF)、广州多宇多生物科技有限公司(DYD)及河南省济源白云實业有限公司和中国科学院动物研究所联合出品(JYBY)。本试验所使用的桶形诱捕器为中捷四方生物科技股份有限公司生产,收集诱集到的草地贪夜蛾雄蛾,记录诱集数量并带回实验室解剖检查交配情况。
1.6 数据分析
采用SPSS 25.0对数据进行统计分析,应用Duncan氏新复极差法进行差异显著性检验,两组数据间的相关性分析采用Pearson相关系数进行相关性分析。
2 结果与分析
2.1 草地贪夜蛾雄性内生殖系统组成
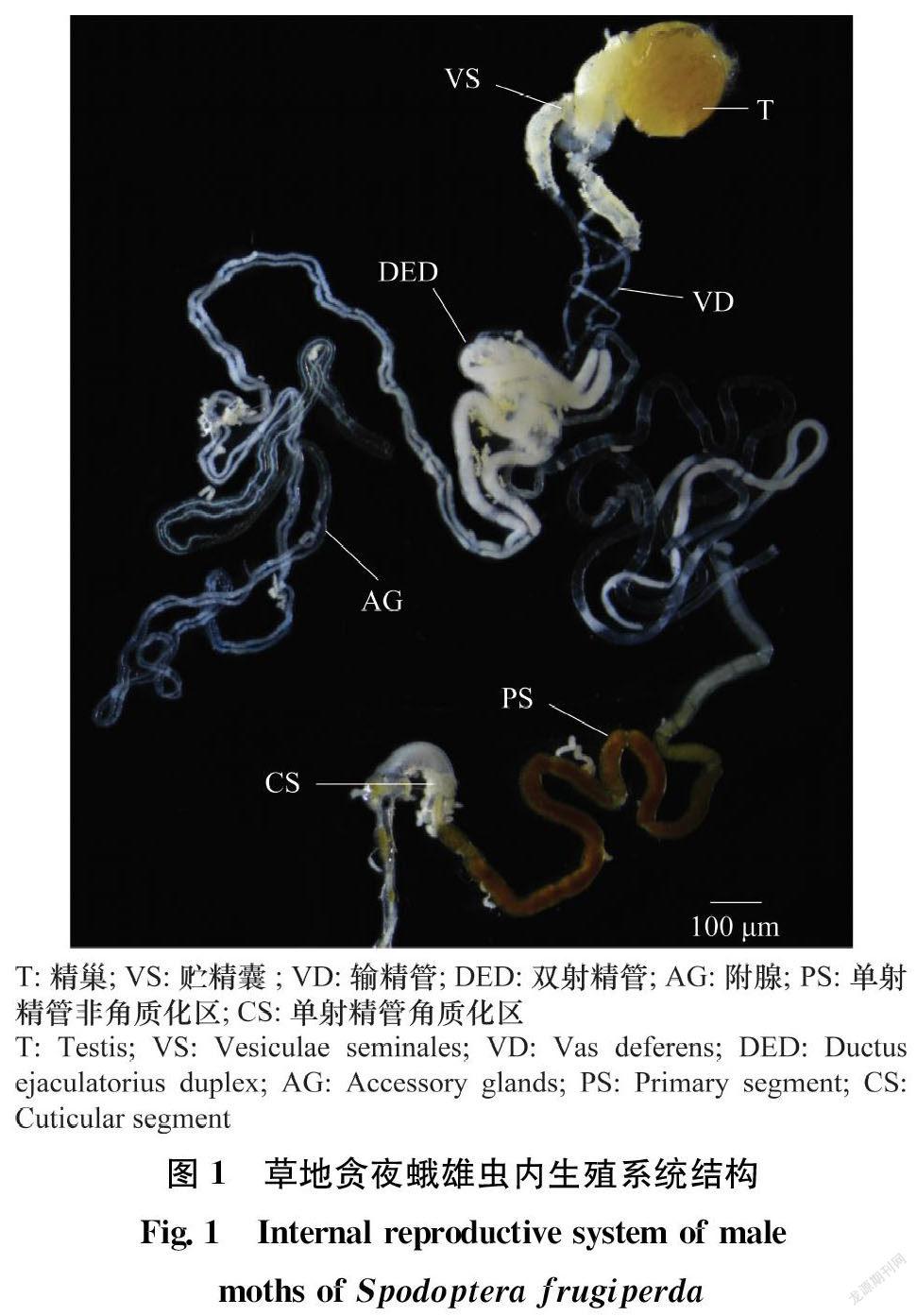
草地贪夜蛾雄性内生殖结构主要包括一个精巢(testis,T)、一对贮精囊(vesiculae seminales,VS)、一对输精管(vas deferens,VD)、一对附腺(accessory glands,AG)、一对双射精管(ductus ejaculatorius duplex,DED)、一条单射精管(ductus ejaculatorius simplex)等,其中单射精管可以分为单射精管非角质化区(primary segment, PS)和角质化区(cuticular segment, CS)(图1)。精巢近椭圆形,黄色不透明,外面包被着一层透明的被膜,主要是产生精子的部位;贮精囊与精巢相连接,通常是输精管膨大形成,中间缢缩,分为两段,尤其在生长发育后期明显膨大;与贮精囊相连的是输精管,输精管是输送精子的通道,成对出现,末端连接双射精管;双射精管1对,乳白色不透明;贮精囊和双射精管是储存精子的主要场所;双射精管一端连接1对附腺,附腺相互独立而又紧密贴合,是分泌精液的主要部位;另一端连接单射精管,精子会在精液的保护下进入单射精管,与单射精管内分泌物质形成精包,通过单射精管到达阳茎,最终在交配时进入雌虫体内完成授精。单射精管PS区为连接双射精管一端柔软细长的管道,PS区末端略微膨大,内含物比较特殊,是判别交配状态的重要部位。未交配时PS末端充满褐色内含物,交配后为透明无色,而后内含物回充。
2.2 草地贪夜蛾雄蛾PS末端内含物分级标准及日龄和交配状态对内含物的影响
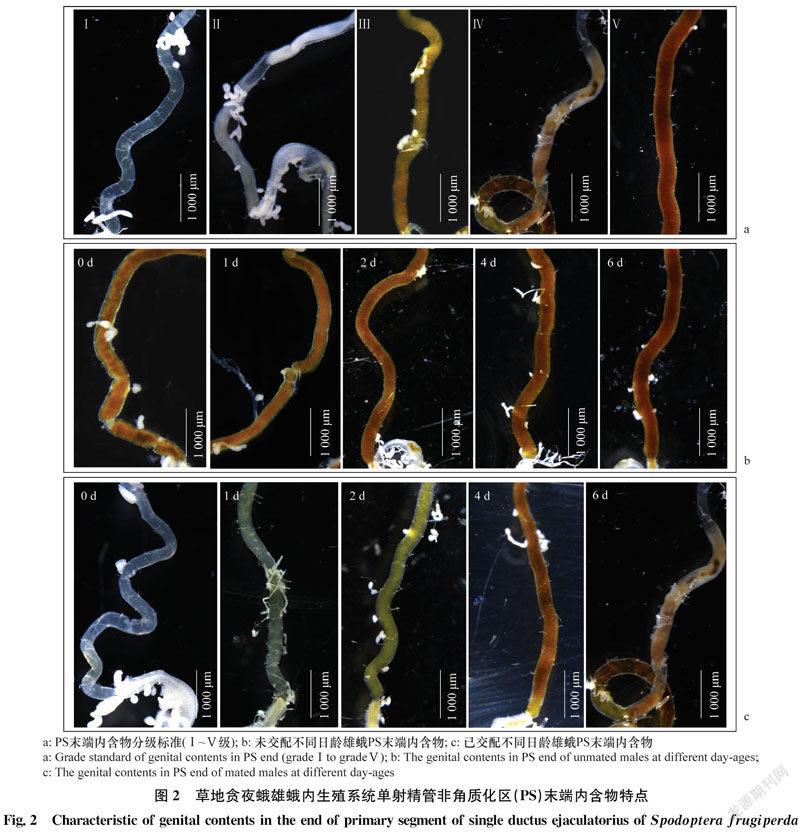
通过对草地贪夜蛾雄蛾内生殖系统的解剖观察,根据PS区末端内含物颜色和形态变化将PS区末端分为5个级别:Ⅰ级,透明状态;Ⅱ级,白色或黄色半充满状态;Ⅲ级,黄色或浅褐色充满状态,质地均匀但无紧致结构;Ⅳ级,褐色未充满状态,内含物质地不均匀、部分出现断裂结块现象;Ⅴ级,褐色或深褐色充满状态,内含物质地均匀、结构紧致(图2a)。
通过对不同日龄和交配状态的雄蛾解剖观察,日龄对未交配雄蛾PS区内含物等级无显著影响,不同日龄内含物均呈现为褐色或深褐色、不透明、结构紧密的状态,分级为Ⅴ级(图2b)。交配之后PS末端内含物等级发生明显变化(图2c):交配后 PS末端褐色内含物排空呈透明状态,分级为Ⅰ级;交配1 d 后,内含物回充呈白色或淡黄色未充满状态,分级为Ⅱ级;交配2 d后,内含物继续回充呈黄色充满状态,分级为Ⅲ级;交配3~4 d,内含物逐渐充满,分级为Ⅴ级;随着时间推移,交配6 d内含物会发生堆积、断层,分布不均匀等特征,分级为Ⅳ级。
2.3 草地贪夜蛾雄蛾精巢体积与日龄的相关性
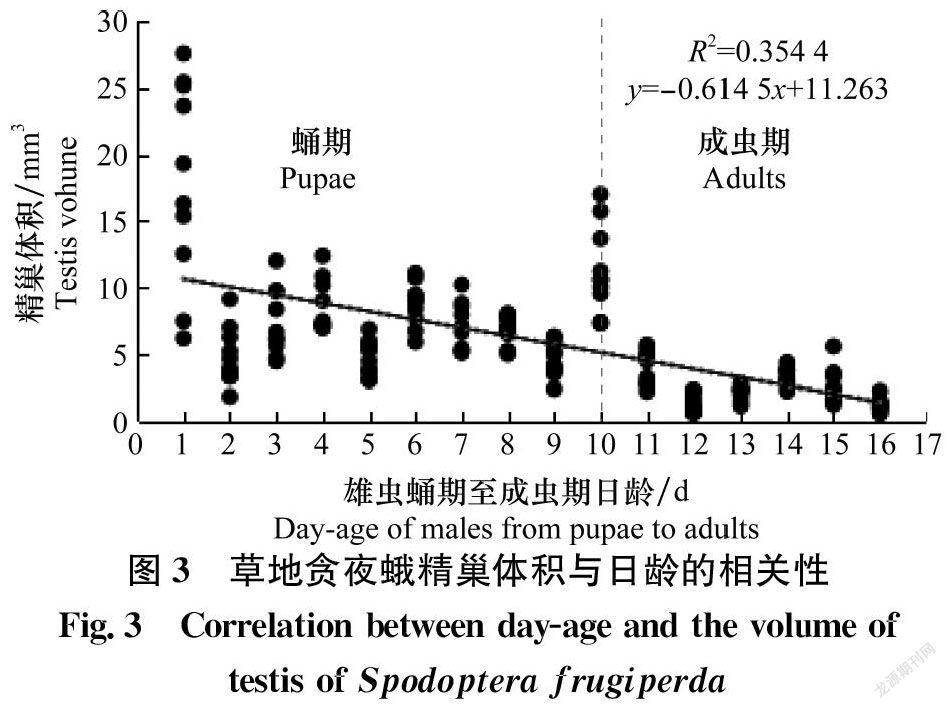
根據对草地贪夜蛾蛹期-成虫期精巢体积与日龄进行Pearson相关性分析,得到Pearson相关系数为-0.595,草地贪夜蛾精巢体积与日龄之间存在显著线性相关性(P <0.05)(图3)。精巢体积在化蛹当天和羽化当天会有两次峰值,即在蛹期1日龄精巢体积达到(17.88±2.40)mm3,随后减小;蛹期10日龄(羽化当日)出现第二次峰值(11.31±1.02)mm3,显著低于1日龄蛹的精巢体积(P < 0.05),显著高于1日龄成虫(图3,11日龄)精巢体积(4.10±0.39)mm3(P < 0.001),6日龄成虫(图3,16日龄)精巢体积减小至(1.23±0.38)mm3。
2.4 性诱草地贪夜蛾雄蛾田间交配率调查
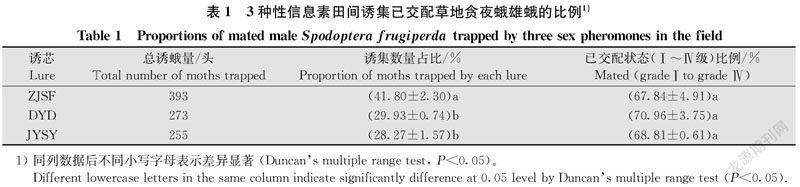
根据对草地贪夜蛾雄蛾田间诱集数据分析,ZJSF诱芯诱集数量显著高于其他两种诱芯(P < 0.05),DYD和BYSY诱芯诱集效果无显著差异(P > 0.05)(表1)。将诱集到的921头雄蛾进行解剖观察,其中647头雄蛾属于已交配状态(Ⅰ~Ⅳ级),3种诱芯田间诱集的雄蛾属于已交配状态(Ⅰ~Ⅳ级)的比例无显著差异(P > 0.05)(表1)。
3 讨论
鳞翅目幼虫期雄虫精巢通常是左右对称的肾形,但随虫体发育,两个肾形精巢会逐渐靠近,然后内膜和膜融合,两个精巢便融合成单个精巢[15];本研究观察到草地贪夜蛾雄虫在预蛹期两个肾形精巢就已经相互靠拢融合,蛹期0日龄精巢已融合成一个完整的精巢结构,之后开始逐渐形成一个完整的内生殖结构,蛹期历期10 d左右。文靓等[16]研究表明,斜纹夜蛾雄虫的精巢体积会随着日龄的增加而变小。这与本研究结果一致。和伟等[13]根据精巢大小对草地贪夜蛾雄蛾制定的划分日龄标准,达到预测雌蛾产卵和幼虫孵化动态的目的,而结合草地贪夜蛾雄蛾的交配状态,可以更准确地预测雌蛾交配时期,从而推测虫量发生为害高峰期。
在自然界中,大多数的鳞翅目昆虫雄蛾一生可进行多次交配[17],而草地贪夜蛾雄蛾一生交配多达6次左右[18]。通过单射精管内含物颜色变化和分布状态可判断亚洲玉米螟 Ostrinia furnacalis和二化螟 Chilo suppressalis雄蛾的交配状态[19, 4],本研究认为此方法也可用于草地贪夜蛾雄蛾交配状态的判断。不同鳞翅目种类雄蛾的单射精管内含物颜色不同,如二化螟、玉米螟内含物为乳白色,而草地贪夜蛾内含物为褐色。不同鳞翅目种类雄蛾交配后内含物恢复状态不一致,如二化螟只能区别交配后12 h以内的交配状态[20],亚洲玉米螟内含物在交配60 h后恢复到未交配的状态[19];蔷薇斜条卷蛾Choristoneura rosaceana内含物恢复时间达6 d[5];桃小食心虫Carposina sasakii和梨小食心虫Grapholitha molesta内含物会在72 h后完全恢复[10]。本研究草地贪夜蛾内含物可在2 d内充满,但恢复至未交尾状态需要4 d,交配6 d后的内含物与未交配的内含物状态有明显区别。本试验3种诱芯诱集的草地贪夜蛾雄蛾有69.02%表现出已交配Ⅰ~Ⅳ等级状态,按本研究的分级标准分析,性诱剂田间诱捕到已交配的雄蛾实际比例更高。在草地贪夜蛾测报和防控中,可通过田间诱集雄蛾精巢大小判断日龄[13],结合本研究方法确定其交配状态,可更准确预测草地贪夜蛾的发生时期,为田间防控提供参考。
性诱剂在害虫预测预报和防治中广泛应用,本试验3种草地贪夜蛾性诱剂在田间的诱捕量有显著差异,但诱捕到的已交配雄蛾比例无显著差异,高达67.84%~70.96%,略低于二化螟比例85%[4],显著高于亚洲玉米螟比例39.1%[19],表明草地贪夜蛾在交配后仍有较强活动能力,并具有多次交配的能力。因此,性信息素对草地贪夜蛾的实际控害效果有待进一步研究。
参考文献
[1] PASHLEY D P, JOHNSON S J, SPARKS A N. Genetic population structure of migratory moths genetic population structure of migratory moths: the fall armyworm (Lepidoptera: Noctuidae) [J]. Annals of the Entomological Society of America, 1985, 78(6):756-762.
[2] WESTBROOK J K, NAGOSHI R N, MEAGHER R L, et al. Modeling seasonal migration of fall armyworm moths [J]. International Journal of Biometeorology, 2016, 60(2): 255-267.
[3] 巴吐西, 张云慧, 张智, 等. 草地贪夜蛾对小麦和玉米的产卵选择性及其种群生命表[J]. 植物保护, 2020, 46(1): 17-23.
[4] HU Yang, DAI Changgeng, HE Jiachun, et al. A technique for determining the mating status of Chilo suppressalis (Lepidoptera: Crambidae) males [J]. Journal of Entomological Science, 2018, 53(1):11-16.
[5] EVENDEN M L, DELURY L E, JUDD G J R, et al. Assessing the mating status of male obliquebanded leafrollers Choristoneura rosaceana (Lepidoptera: Tortricidae) by dissection of male and female moths [J]. Annals of the Entomological Society of America, 2003, 96(3): 217-224.
[6] 冯波, 张万民, 张丹, 等. 亚洲玉米螟交配率和交配次数与其日龄、性比和精巢大小的关系[J]. 昆虫学报, 2020, 63(1): 54-62.
[7] DELISLE J, BOUCHARD A. Male larval, nutrition in Choristoneura rosaceana (Lepidoptera: Tortricidae): an important factor in reproductive success [J]. Uecologia, 1995, 104(4): 308-314.
[8] 嵇保中, 刘曙雯, 曹丹丹. 昆虫生殖系统[M]. 北京: 科学出版社, 2014: 727-729.
[9] 周娇, 李娟, 翁强, 等. 沙棘木蠹蛾生殖系统形态学和组织学观察[J]. 应用昆虫学报, 2012, 49(6): 1629-1635.
[10]彭云飞. 桃小食心虫和梨小食心虫生殖及消化系统发育的形态学研究[D]. 晋中: 山西农业大学, 2014.
[11]李传瑛, 章玉苹, 黄少华, 等. 草地贪夜蛾室内人工饲养技术的研究[J]. 环境昆虫学报, 2019, 41(5): 986-991.
[12]王世英, 朱启绽, 谭煜婷, 等. 草地贪夜蛾室内人工饲料群体饲养技术[J]. 环境昆虫学报, 2019, 41(4): 742-747.
[13]和伟, 赵胜园, 葛世帅, 等. 草地贪夜蛾种群性诱测报方法研究[J]. 植物保护, 2019, 45(4): 48-53.
[14]冯波, 郭前爽, 姚晓明, 等. 二化螟雄蛾交配行为与精巢大小的关系[J].昆虫学报, 2019, 62(7): 849-856.
[15]刘琳, 冯启理. 鳞翅目昆虫精巢融合的研究[J]. 华南师范大学学报(自然科学版), 2014, 46(5): 1-7.
[16]文靓, 陈丰, 陈亚青, 等. 斜纹夜蛾精巢的发育和精子的发生[J]. 华南师范大学学报(自然科学版), 2019, 51(4): 47- 56.
[17]THORNHILL R, ALCOCK J. The evolution of insect mating systems [M]. Massachusetts: Harvard University Press, 1983.
[18]SIMMONS A M, MARTI O G. Mating by the fall armyworm (Lepidoptera: Noctuidae): frequency, duration, and effect of temperature [J]. Environmental Entomology, 1992, 21(2): 371-375.
[19]馮波, 张万民, 张丹, 等. 日龄和交配状态对亚洲玉米螟雄蛾内生殖器特征的影响[J]. 昆虫学报, 2020, 63(3): 317- 326.
[20]郭前爽, 王春荣, 宋显东, 等. 日龄和交配状态对二化螟雄蛾内生殖器特征的影响[J]. 昆虫学报, 2019, 62(7): 838- 848.
(责任编辑:杨明丽)
猜你喜欢
东坡赤壁诗词(2022年4期)2022-10-30 12:18:17
四川蚕业(2021年3期)2021-02-12 02:38:48
昆虫学报(2020年6期)2020-08-06 06:42:52
农家科技中旬版(2019年8期)2019-09-19 04:26:07
植物保护(2019年4期)2019-09-04 09:34:39
南方农业·下旬(2019年4期)2019-08-26 01:24:22
山东农业科学(2019年6期)2019-08-16 04:05:54
现代农业科技(2019年12期)2019-08-07 01:04:27
安徽农业科学(2018年22期)2018-05-14 08:59:50
蚕学通讯(2017年3期)2017-03-23 08:00:02
