
粉垄耕作木薯根际与非根际土壤的细菌群落结构多样性差异
2022-05-28 09:30覃锋燕杨慰贤彭晓辉黎亮武阳太亿韦茂贵申章佑
西南农业学报 2022年4期
覃锋燕,杨慰贤,彭晓辉,黎亮武,阳太亿,周 佳,韦茂贵,3,申章佑
(1.广西大学农学院,南宁 530004;2.广西农业科学院经济作物研究所,南宁 530007;3.广西高校作物栽培学与耕作学重点实验室/广西大学,南宁 530004)
【研究意义】耕作方式通过改变土壤理化性质及土壤微生物群落多样性和数量,影响作物对养分的吸收和利用,最终表现为作物产量存在差异[1-3]。不同耕作方式对土壤理化性质和微生物特性的影响存在差异,松耕使土壤养分从表层向深层土壤转移,深翻可显著提高土壤酶活性、微生物数量、微生物量碳和氮[4-5]。粉垄耕作是一种新型耕作技术,可实现全耕层深耕深松,有效加深耕作层、均匀细碎土壤、提高土壤透气性及保水蓄水能力,促进作物根系生长、增加产量及改善品质[6-7]。此外,土壤微生物作为土壤最活跃的部分,参与土壤有机残体的分解与养分循环,在生态系统植物多样性和生产力方面发挥关键作用[8]。土壤细菌作为土壤微生物的重要组成部分,通过参与土壤环境中有机残体的分解和转化过程,合成腐殖质,释放养分,从而调节土壤结构与肥力的稳定性,促进植物生长发育[9]。我国木薯主要种植在南方黄红壤地区,土壤耕层浅薄,加上连作障碍及干旱的双重胁迫[10],土壤板结程度逐年加重,但前期通过粉垄技术栽培的木薯,在减量施肥条件下仍能增产,实现提质增效[11]。因此,揭示粉垄木薯的增产机制,优化现有木薯耕作方式,对助力木薯产业健康发展具有重要意义。【前人研究进展】张薇等[12]、钟文辉和蔡祖聪[13]、Vitali等[14]研究认为,植物群落、土壤类型、耕作措施和土壤理化性质可影响土壤微生物的多样性。近年来关于耕作方式的研究主要集中在免耕、深松深耕和翻耕等耕作类型[15-17],其中,免耕、秸秆还田和深松深耕等保护性耕作措施通过改善土壤质地及促进土壤碳氮转化而增加土壤微生物数量和微生物量碳,在减少碳素亏损的同时提高土壤脲酶活性,从而提高土壤肥力[18-19];在免耕措施下,土壤的微生物多样性、丰富度和均匀度指数与撂荒方式相近,但均高于翻耕和旋耕方式[20]。免耕覆盖和深松免耕覆盖均可明显影响土壤微生物数量和微生物量,即使不改变土壤微生物的组成和种类,仍影响其相对丰度[21]。水稻粉垄耕作可在短时间内改变水稻根际土壤细菌群落多样性,其中,变形菌门和浮霉菌门的相对丰度均表现为粉垄高于免耕和旋耕[22-23]。粉垄耕作相对于旋耕处理能提高小麦成熟期潮土土壤的速效钾含量,显著降低土壤碱解氮、有效磷和有机质含量[24],而速效氮、速效钾和土壤pH等理化性质是影响土壤细菌多样性和丰度的主要因子,其中速效氮和速效钾含量与Shannon和Chao1指数呈显著相关,土壤pH影响放线菌门和拟杆菌门的相对丰度[25]。相对于常规耕作方式,粉垄耕作可有效提高土壤微生物数量和土壤酶活性,从而改变土壤化学性质,改善土壤微生物生存的微环境[26]。可见,由于土壤类型、植物类型和耕作制度等因素的影响,不同耕作方式对土壤微生物数量、微生物量碳和氮及群落多样性的影响效果不同。【本研究切入点】目前,关于粉垄耕作应用于旱地作物的研究大多集中在其对作物增产提质及土壤理化性质优化方面[27-28],也有研究课探讨了粉垄耕作对木薯非根际土壤细菌群落多样性的影响[29],但粉垄耕作对木薯根际细菌群落影响的研究未见报道。【拟解决的关键问题】以常规耕作为对照,利用高通量测序技术和生物信息学手段,分析粉垄耕作对木薯根际和非根际土壤理化性质及细菌群落多样性的影响,为阐释粉垄耕作对木薯地细菌群落多样性的形成机制提供科学依据,也为粉垄耕作技术的推广提供理论依据。
1 材料与方法
1.1 试验地概况
试验于2019年4—12月在广西农业科学院武鸣里建试验基地(东经107°49′26″,北纬22°59′58″)进行。耕作前采集耕作层(0~20 cm)土壤,测定其pH为6.24,有机质含量为17.23 g/kg,碱解氮含量为60.38 mg/kg,速效磷含量为45.09 mg/kg,速效钾含量为42.17 mg/kg[30]。
1.2 试验材料
供试木薯品种为广西主栽品种华南205(中国热带农业科学院选育)。
1.3 试验方法
1.3.1 试验设计 设常规耕作和粉垄耕作两个不同耕作方式处理,其中,常规耕作地块先犁地(深30~35 cm),再使用纽荷兰牌110马力拖拉机卧式旋耕耙两轮,耙地深度15~18 cm;粉垄耕作地块直接使用广西五丰机械公司的LG-125型自走式粉垄机,一次完成全土层土壤粉碎并形成小平垄,耕作深度30~35 cm。每个耕作田块里划分为24个70 m2的试验小区(5 m ×14 m),随机挑选4个小区作为重复。
木薯于2019年4月16日种植,种茎长约15 cm,株行距1 m × 1 m。木薯整个生育期的N、P2O5和K2O施用量分别为358.80、89.10和187.50 kg/hm2,其中,于植后60 d撒施60%,植后120 d撒施40%。施肥后用薄土覆盖,试验期间无人工灌水。除耕作方式不同外,两个耕作处理的其他田间管理措施一致。
1.3.2 土壤样品采集与处理 前期研究发现,木薯地在7月的温室气体排放通量最高[30],故土壤样品采集安排在木薯块根形成期(植后90 d)进行。根际土壤采集:每个小区随机选择3株木薯,挖出完整的块根,抖去大块土壤,采用抖根法采集根系上的土壤。非根际土壤采集:各小区在离木薯根际0.5 m处按S形随机选5个点,分别用取土器采集0~20 cm深的土样,混匀后作为该小区非根际土样。土壤样品分别置于塑料薄膜上,剔除植物残体、石砾及其他杂物,混匀后用四分法将土壤样品分为3份:1份立即用液氮速冻,放入冰盒中保存带回实验室于-80 ℃条件下保存,用于测定土壤微生物群落多样性;另1份做好标记,带回实验室于冰箱4 ℃保存,用于测定土壤酶活性;最后1份置于写好标签的自封袋内,常温保存带回实验室,风干后磨碎过筛,用于测定土壤理化性质。
1.3.3 土壤理化性质及酶活性测定 土壤pH采用酸度计法测定(雷磁pHS-2F,上海),有机质含量采用重铬酸钾容量法测定[31],碱解氮含量采用碱解扩散法测定[32],土壤速效磷含量使用钼酸比色法测定,土壤速效钾含量采用NH4OAc浸提火焰光度计测定[33];土壤脲酶活性采用苯酚—次氯酸钠比色法测定[34],过氧化氢酶活性采用紫外分光光度法测定[35],亚硝酸还原酶[36]和硝酸还原酶活性[37][]采用紫外分光光度法测定。
1.3.4 土壤微生物高通量测序及生物信息学分析 委托上海美吉生物医药科技有限公司利用Illumina MiSeq平台进行土壤样品高通量测序。使用FastDNA®Spin Kit for Soil试剂盒(MP biomedicals,USA)提取土壤基因组DNA,以NanoDrop2000检测DNA浓度和纯度,1%琼脂糖凝胶电泳检测DNA提取质量。使用特异引物338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和806R(5’-GGACTACHVGGG TWTCTAAT-3’)对16S rRNA序列的V3~V4可变区进行PCR扩增。PCR扩增产物经2%琼脂糖凝胶回收,用AxyPrep DNA Gel Extraction Kit(Axygen biosciences,Union City,CA,USA)进行纯化,进行2%琼脂糖凝胶电泳检测后,再用QuantusTMFluorometer(Promega,USA)进行定量检测。使用NEXTFLEX Rapid DNA-Seq Kit试剂盒(Bioo scientific,USA)构建文库。利用Illumina MiSeq PE300平台测序后,原始数据上传至NCBI SRA数据库(登录号PRJNA783658,样品序列号SRR17038929-SRR17038944)。
原始测序序列质控和拼接分别使用Trimmomatic(version 0.32)和FLASH(version 1.2.11),使用UPARSE Version 7.1将序列在97%相似度下进行分类操作单元(OTU)聚类。基于Silva数据库,将筛选后的非嵌合序列在RDP classifier(http://rdp.cme.msu.edu/)进行物种分类注释,设置比对阈值为70%。
利用Past 3.0(http://fold.uio.no/ohammer/past)计算各样品的OTU丰富度,利用Mothur Version 1.30.2对OTU进行Alpha多样性分析。主成分分析(PCA)、置换多因素方差分析(PERMANOVA)、冗余分析(RDA),使用R(version 3.3.1)vegan制图,以stats和python的scipy进行组间差异显著性检验。
1.4 统计分析
采用Excel 2010进行数据整理与图表制作,以SPSS 20.0进行方差分析、相关分析和差异显著性检验。
2 结果与分析
2.1 不同耕作方式对木薯根际和非根际土壤养分含量和酶活性的影响
由表1可知,粉垄耕作木薯非根际和根际土壤的pH和有机质含量均显著高于常规耕作土壤(P<0.05,下同);粉垄耕作非根际的土壤速效钾含量显著高于常规耕作的非根际土壤,根际土的土壤速效钾含量高于常规耕作的根际土壤,但差异不显著(P>0.05,下同);两种耕作方式的土壤碱解氮含量间及有效磷含量间均无显著差异;除常规耕作根际与非根际的土壤脲酶活性间差异显著外,其他酶活性间均无显著差异。说明不同耕作处理木薯根际与非根际土壤的养分含量和酶活性存在差异。
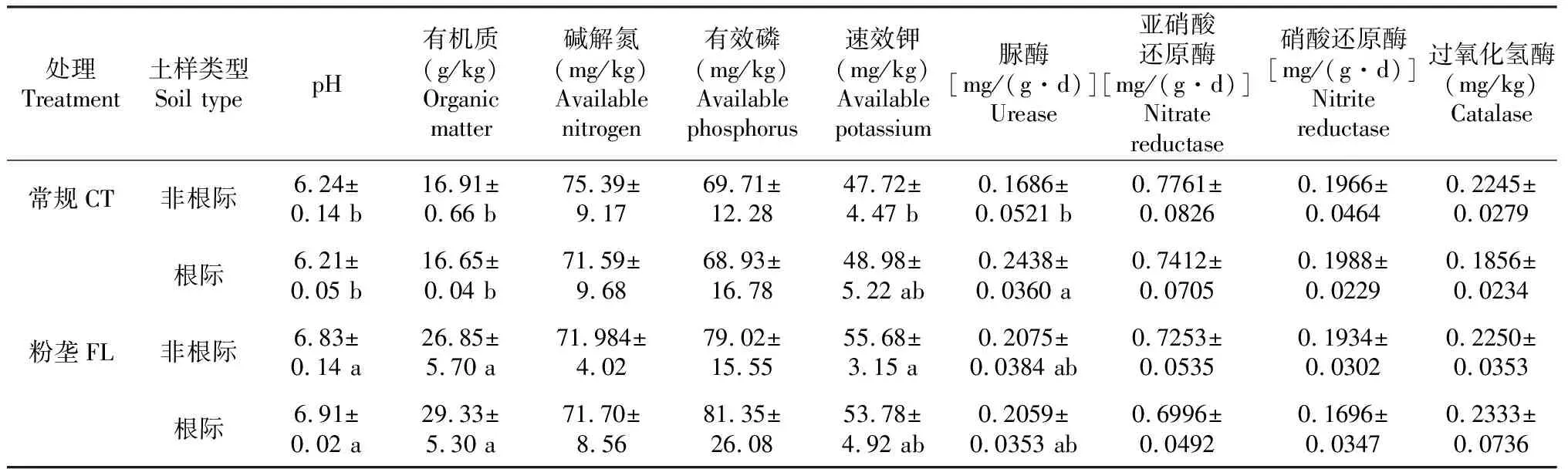
表1 不同耕作处理对木薯根际和非根际土壤养分含量及酶活性的影响
2.2 不同耕作处理对木薯根际和非根际土壤细菌群落多样性的影响
2.2.1 对α多样性的影响 由表2可知,木薯根际土壤与非根际土壤间细菌群落的α多样性存在极显著差异(P<0.01,下同),土壤类型×耕作方式互作也显著影响其细菌群落的α多样性,而2种耕作方式木薯根际土壤与非根际土壤间细菌群落的α多样性无显著差异。从图1也可看出,木薯根际土壤细菌群落的α多样性显著高于非根际土壤。由表3可知,各耕作方式的文库覆盖率均超过95.00%,计算各处理细菌群落的多样性指数(Shannon和Simpson指数)及群落丰富度指数(Chao1和Ace指数),结果表明,常规耕作非根际土壤的Shannon指数大于根际土壤,但差异不显著,而粉垄耕作非根际土壤的Shannon指数小于根际土壤,说明常规耕作非根际土壤的细菌群落多样性大于根际土壤,而粉垄耕作根际土壤细菌群落的多样性大于非根际土壤;2种耕作方式根际土壤的Ace和Chao1指数均大于非根际土壤,但无显著差异,说明木薯根际土壤的细菌群落较非根际土壤丰富。
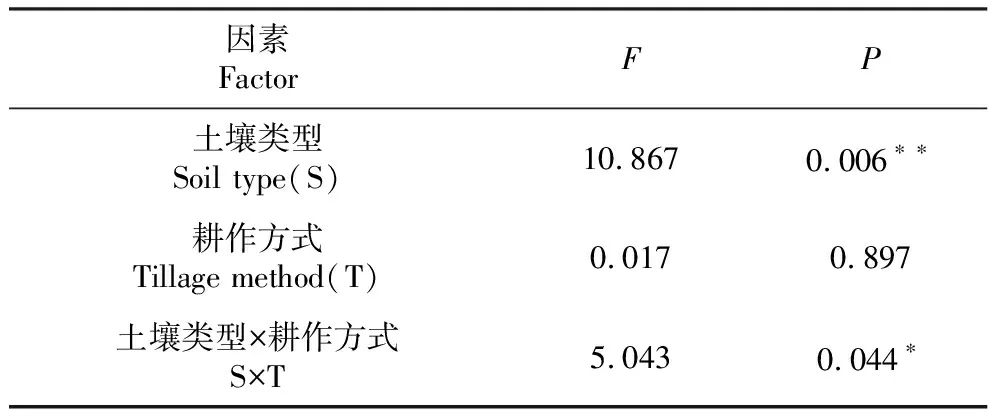
表2 各土壤样本OTU丰富度的方差分析
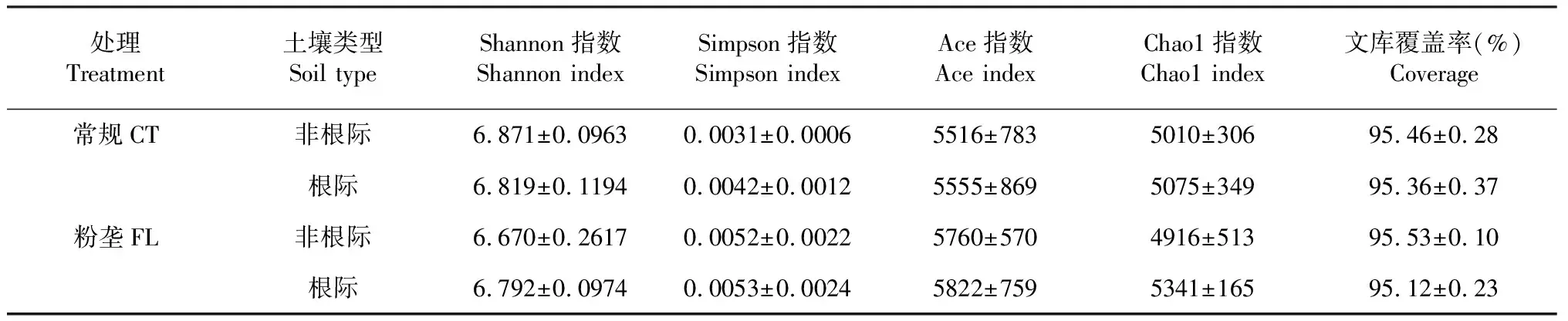
表3 不同耕作处理木薯根际土壤和非根际土壤细菌的α多样性指数比较
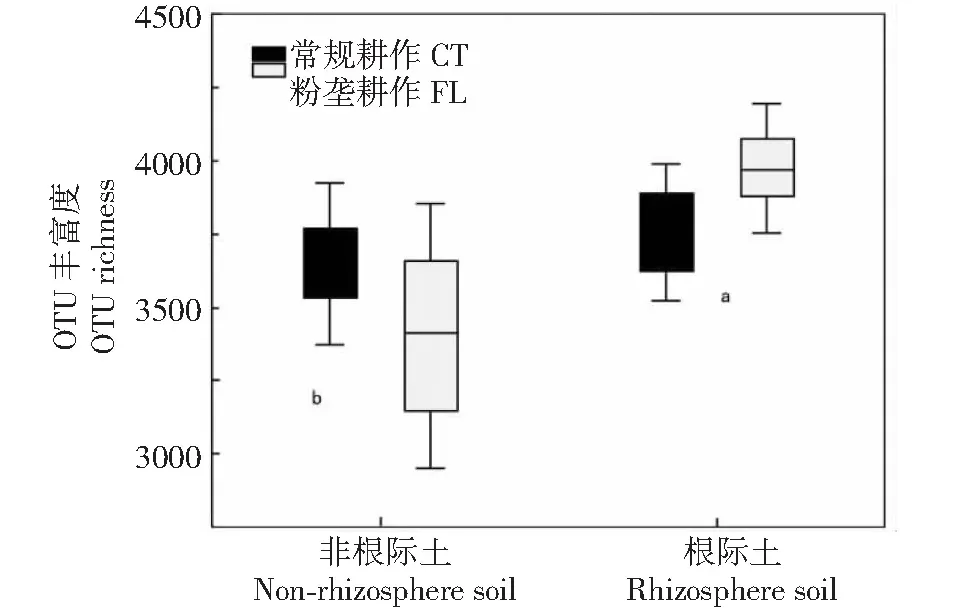
不同小写字母表示根际与非根际间各土壤样本OTU丰富度差异显著
2.2.2 对β多样性的影响 从图2可看出,非根际土壤的主成分分析结果表明,第一主成分(PC1)可解释总方差的58.21%,可能是因粉垄耕作方式中个别重复肥力差异较大引起;第二主成分(PC2)将2种耕作方式分别划分到正、负2个方向,可解释总方差的16.66%,可认定为耕作方式不同引起的差异(图2-A);根际土壤的主成分分析结果显示,不考虑离群样本,第二主成分(PC2)将2个耕作方式分成2个群体,可解释总方差的26.87%,可认定为耕作方式不同引起的差异(图2-B)。
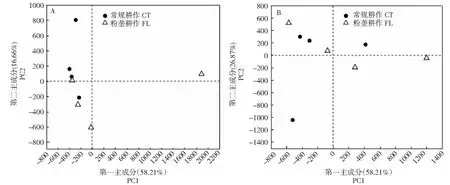
A:非根际土壤;B:根际土壤。图5和图6同
基于细菌菌落半度量距离(Bray-Curtis)和未加权的Unifrac距离,使用PERMANOVA方法,分别计算单个因素对细菌群落变化的贡献。由表4可知,在Bray-Curtis算法中,耕作方式是影响土壤细菌群落分类和系统发育β多样性差异的主要因素,土壤类型对β多样性差异的影响不显著;在未加权的Unifrac算法中,土壤类型和耕作方式对土壤细菌群落分类和系统发育β多样性差异均具有显著影响。
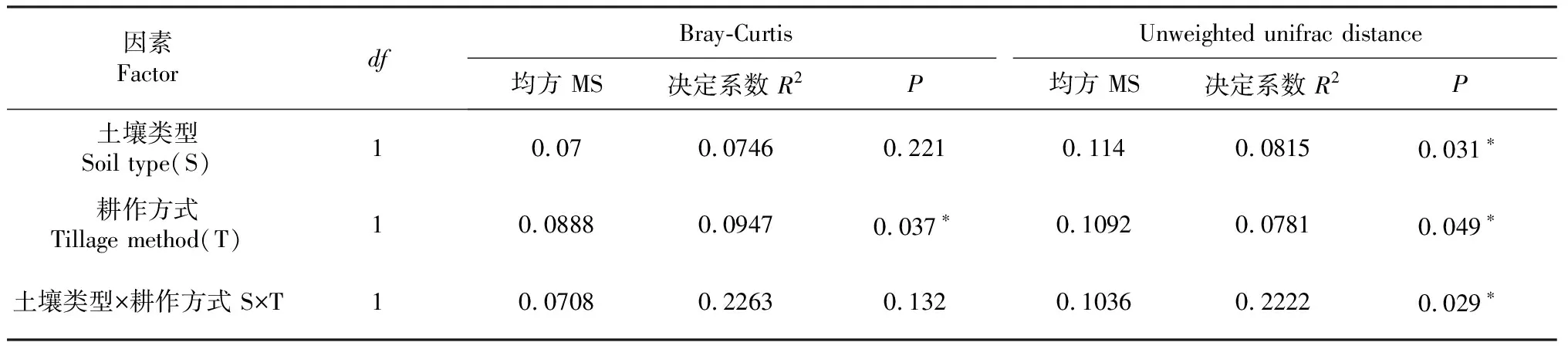
表4 基于Bray-Curtis和Unweighted unifrac距离的PERMANOVA分析
2.3 不同耕作方式对木薯根际和非根际土细菌群落结构的影响
2.3.1 对细菌群落组成的影响 从图3可看出,不同耕作方式木薯根际和非根际土壤的细菌门水平和纲水平群落结构组成基本相同;所有土壤样品中均主要含放线菌门(Actinobacteriota)、变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、厚壁菌门(Firmicutes)、酸杆菌门(Acidobacteriota)、芽单胞菌门(Gemmatimonadota)、Myxococcota、Methylomirabilota、拟杆菌门(Bacteroidota)、浮霉菌门(Planctomycetota)、硝化螺旋菌门(Nitrospirota)、蓝藻门(Cyanobacteria)和疣微菌门(Verrucomicrobiota)等。其中,占主导地位的细菌类群有放线菌门(占比20.03%~21.45%)、变形菌门(占比18.13%~21.70%)、绿弯菌门(占比15.92%~16.73%)、厚壁菌门(占比8.68%~14.08%)和酸杆菌门(占比9.59%~13.29%),占总相对丰度的79.89%~81.38%;2种耕作方式非根际土壤放线菌门、绿弯菌门、酸杆菌门和芽单胞菌门的相对丰度均大于根际土壤;粉垄耕作方式根际土壤变形菌门的相对丰度大于非根际土壤,非根际土壤后壁菌门的相对丰度大于根际土壤,而常规耕作方式与粉垄耕作方式相反。在纲分类水平,α-变形菌纲(Alphaproteobacteria)、放线菌纲(Actinobacteria)、芽孢杆菌纲(Bacilli)和γ-变形菌纲(Gammaproteobacteria)为土壤细菌的主要优势菌群(图4)。其中,粉垄耕作方式根际土壤α-变形菌纲的相对丰度大于非根际土壤,而常规耕作方式相反;放线菌纲的相对丰度在粉垄耕作方式中是根际土壤小于非根际土壤,在常规耕作中为根际土壤大于非根际土壤;2种耕作方式土壤芽孢杆菌纲和γ-变形菌纲的相对丰度均以根际土壤大于非根际土壤。综上所述,根际土壤与非根际土壤细菌在门和纲水平的群落组成基本相同,但其细菌群落的相对丰度存在差异。
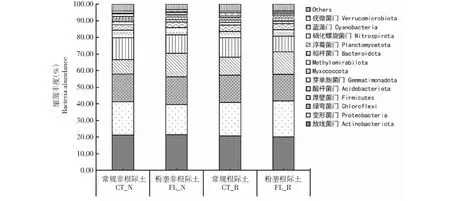
图3 2种耕作处理木薯根际土壤和非根际土壤细菌群落的门水平组成

图4 2种耕作处理木薯根际土壤和非根际土壤细菌群落纲水平的聚类分析热图
2.3.2 细菌群落组成差异分析 从常规耕作和粉垄耕作木薯根际土壤和非根际土壤的细菌群落中选取相对丰度均存在显著或极显著差异排名前10位的OTUs进行细菌群落组成差异分析,结果(图5)表明,常规耕作方式木薯根际土壤和非根际土壤细菌群落的OTUs被分类为黄色杆菌科(OTU3214)、芽单胞菌科(OTU2248)、硝化螺旋菌属(OTU6678)、苯基杆菌属(OTU8035)、Azohydromonas属(OTU4105)、假诺卡氏菌属(OTU9628)、TK10纲(OTU476)、Gaiellales目(OTU6143)、酸杆菌目(OTU7972)和Conexibacter属(OTU5869,分别属于变形菌门、芽单胞菌门、硝化螺旋菌门、放线菌门和绿弯菌门(图5-A),其中,OTU8035、OTU4105、OTU9628、OTU7972和OTU5869占主导地位,且在细菌群落组成中均显著或极显著高于非根际土壤;从图5-B可看出,粉垄耕作方式木薯根际土壤和非根际土壤细菌群落的OTUs被分类为芽孢杆菌属(OTU7337)、鞘氨醇单胞菌属(OTU350)、芽单胞菌属(OTU3052)、亚硝化螺菌属(OTU4016)、IMCC26256目(OTU9034)、Ellin6067属(OTU6431)、Vicinamibacteraceae科(OTU6922)、A21b科(OTU7905)、马赛菌属(OTU7179和梭状芽孢杆菌属(OTU2218),分别属于厚壁菌门、变形菌门、芽单胞菌门和放线菌门,其中,OTU7337、OTU3052、OTU9034、OTU6431、OTU7905和OTU7179占主导地位,且在细菌群落组成中均显著或极显著高于非根际土壤。可见,常规耕作木薯根际土壤细菌群落组成排名前10位的OTUs中有5个显著高于非根际土壤,粉垄耕作木薯根际土壤细菌群落组成排名前10位的OTUs中有6个显著或极显著高于非根际土壤。
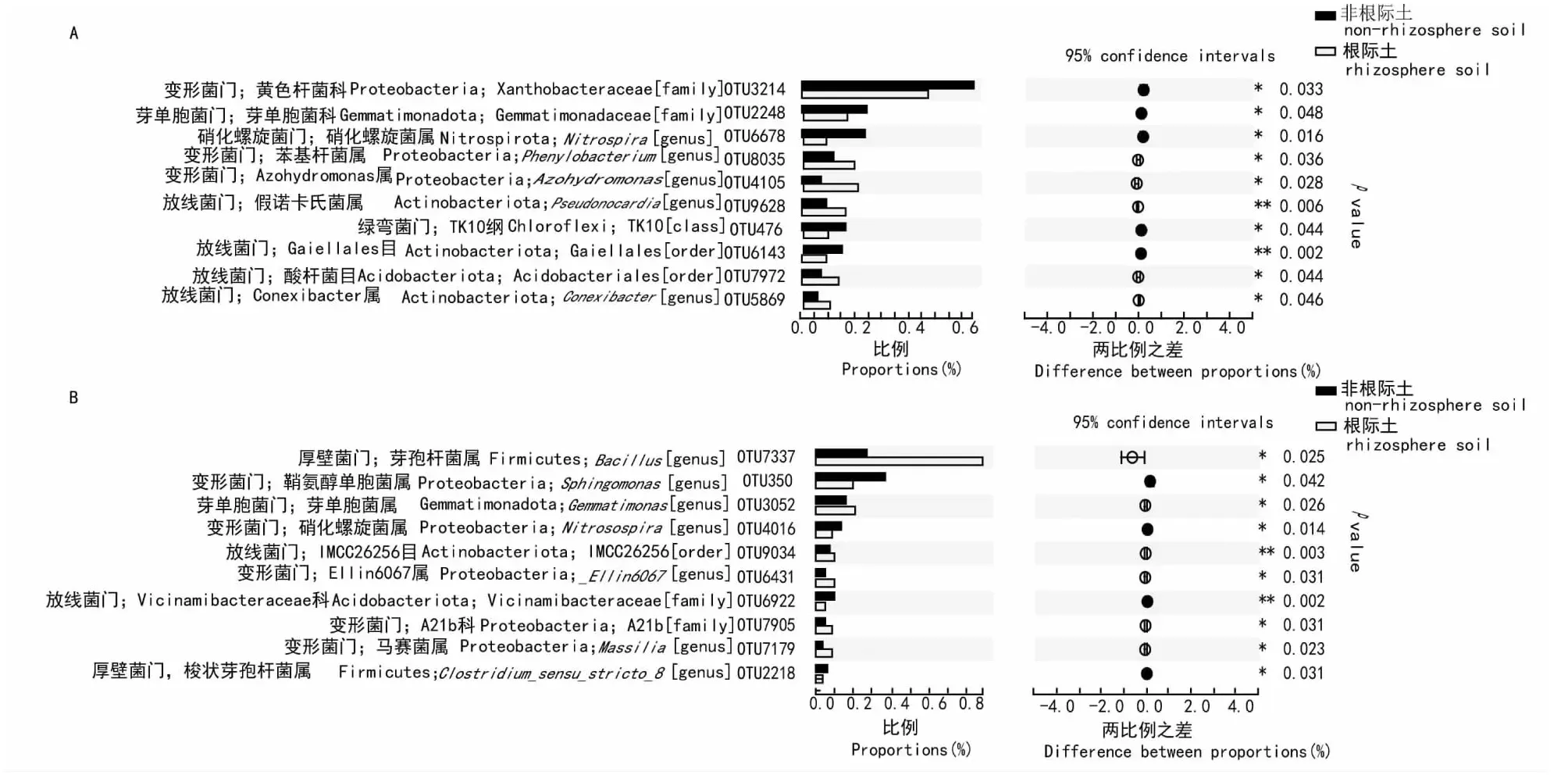
图5 2种耕作处理木薯根际和非根际土壤细菌群落中相对丰度存在显著或极显著差异排名前10位的OTUs差异比较
2.4 不同耕作方式对土壤养分含量与细菌群落组成相关性的影响
从图6可看出,在非根际土壤中(图6-A),厚壁菌门与土壤pH和有机质含量、拟杆菌门与有效磷含量、浮霉菌门和Armatimonadota与硝酸还原酶活性及硝化螺旋菌门、Latescibacterota和脱硫杆菌门与亚硝酸还原酶活性分别呈显著正相关,Methylomirabilota与有机质含量、绿弯菌门与碱解氮含量分别呈显著负相关;在根际土壤中(图6-B),硝化螺旋菌门、unclassified_k_norank_d_Bacteria和蓝藻门分别与脲酶活性、过氧化氢酶活性和速效钾含量分别呈显著负相关,有效磷含量与浮霉菌门、绿弯菌门、疣微菌门和Latescibacterota呈显著负相关。可见,土壤细菌群落组成同时受土壤理化性质和酶活性的影响,但影响程度存在差异。
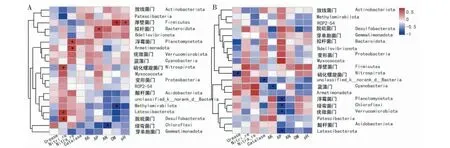
图6 土壤样品细菌群落相对丰度与环境因子在门水平的相关分析
从图7可看出,丰度值靠前的5个细菌门分别为放线菌门、变形菌门、绿弯菌门、厚壁菌门和酸杆菌门,RDA1和RDA2的解释变量分别达35.24%和20.50%;土壤有机质含量、pH、有效磷含量、亚硝酸还原酶活性、过氧化氢酶活性与碱解氮含量是对细菌群落结构影响较大的环境因子,其中,有机质含量与pH和有效磷含量呈正相关,有机质含量和pH是影响厚壁菌门相对丰度的原因之一,过氧化氢酶活性和有效磷含量是影响变形菌门相对丰度的主要原因,亚硝酸还原酶活性是影响酸杆菌门和绿弯菌门相对丰度的主要因素,过氧化氢酶对放线菌门相对丰度有一定影响。粉垄耕作方式土壤中厚壁菌门的相对丰度较高,主要是其较高的有机质含量和pH影响脲酶活性;而常规耕作方式土壤中绿弯菌门和酸杆菌门的相对丰度较高,进而提高了土壤亚硝酸还原酶活性。说明土壤细菌群落的分布与土壤环境因子相互影响。
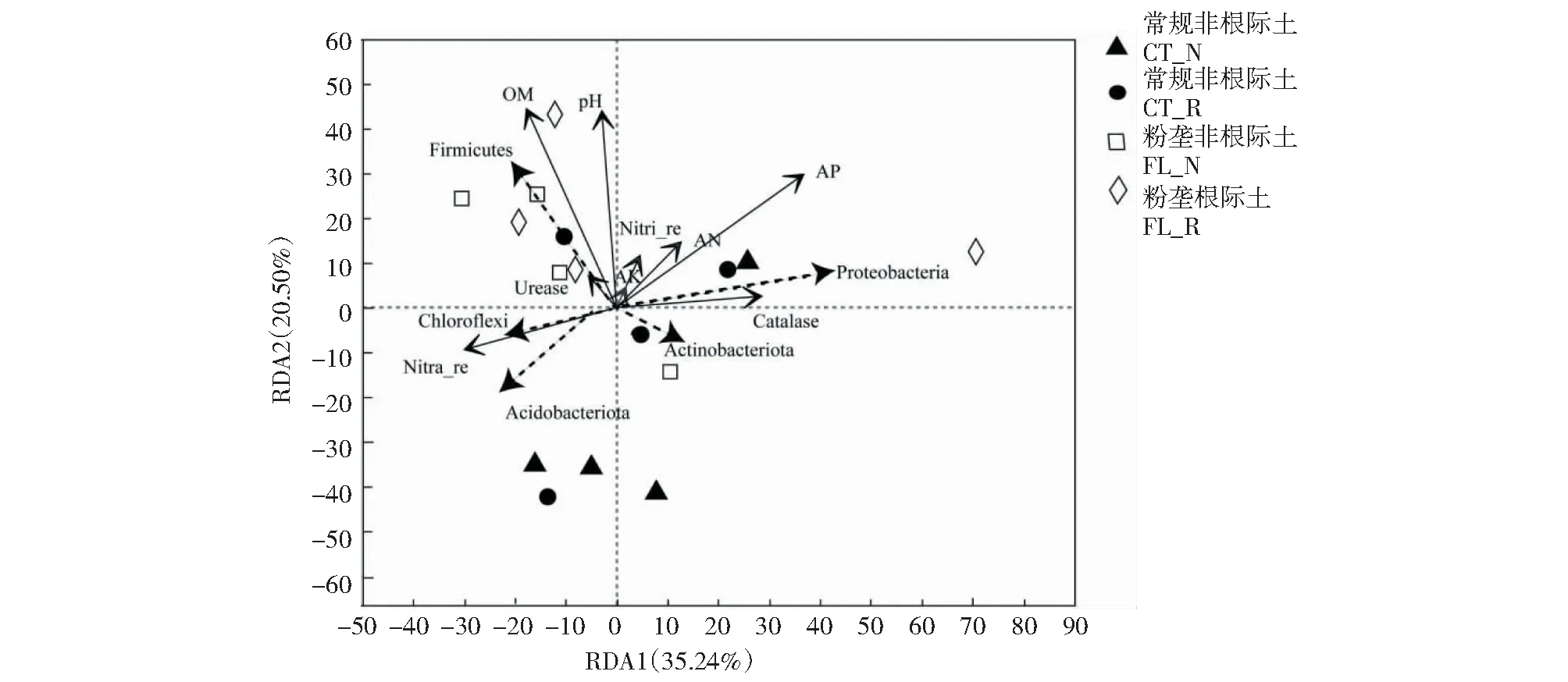
实线箭头代表物种;虚线箭头表示环境影响因子,箭头长短代表环境因子对物种数据的影响程度
对不同耕作方式的土壤细菌群落(门分类水平相对丰度大于0.1%)与理化性质进行相关分析,结果(表5)表明,有8个细菌门与土壤理化性质呈显著相关。其中,变形菌门与有效磷含量呈极显著正相关;绿弯菌门与碱解氮含量和有效磷含量呈极显著负相关、与过氧化氢酶活性呈显著负相关;厚壁菌门与pH呈显著正相关、与有机质含量呈极显著正相关;酸杆菌门与硝酸还原酶活性呈显著正相关;Methylomirabilota与有机质含量呈极显著负相关;硝化螺旋菌门与脲酶活性呈显著负相关、与亚硝酸还原酶活性呈显著正相关;蓝藻门与脲酶活性呈显著正相关。说明对细菌群落影响较大的环境因子为有机质含量、有效磷含量和脲酶活性,而影响最小的因子为速效钾含量。
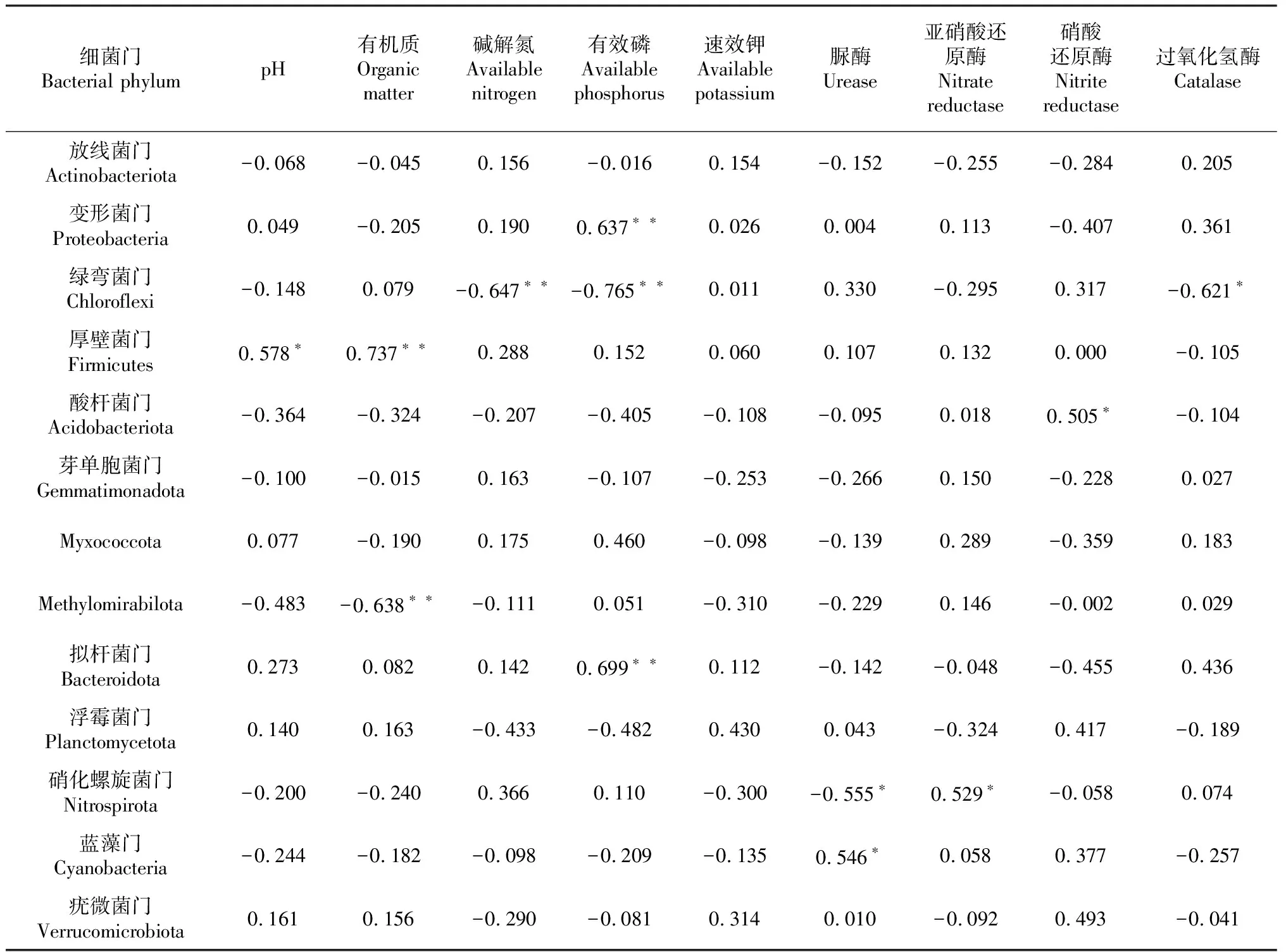
表5 不同耕作处理木薯土壤细菌主要菌群与土壤理化性质的相关分析结果
3 讨 论
粉垄耕作通过深耕深松土壤但土层不变的方式,保留原耕作层和犁底层的良好结构,具有更好的协调土壤水肥气热作用,可减少木薯受旱受渍机率[30]。本研究中,粉垄耕作木薯根际土壤的pH、有机质含量和有效磷含量均大于非根际土壤,而常规耕作木薯根际土壤的pH、有机质含量和有效磷含量均小于非根际土壤,可能是因为在木薯旺盛生长的7月降水量较大,粉垄耕作的土壤疏松有利于雨水下渗,木薯未受逆境胁迫,而常规耕作方式的耕作层土壤湿度明显高于粉垄耕作,土壤水分出现过饱和现象,进而导致木薯受逆境胁迫,诱导根系分泌物有机碳和有机酸,与前人关于冠麦草在受涝害胁迫时分泌有机酸的研究结果一致[38]。土壤酶活性可反映土壤各种生物化学过程,是评价土壤质量的重要指标之一[39]。本研究中,除常规耕作木薯的根际土壤脲酶活性显著高于非根际土壤外,2种耕作方式的其他几种酶活性在非根际土壤与根际土壤间均无显著差异,可能与常规耕作木薯处于水分胁迫下分泌更多的有机酸和氨基酸,引起木薯根际土壤脲酶活性变高有关,与Schlute等[40]、赵宽等[41]的研究结果一致。
土壤微生物参与并主导土壤生态系统的一系列活动,维持土壤生态系统的稳定性[42]。本研究结果表明,木薯根际土壤细菌群落的α多样性显著高于非根际土壤,而粉垄耕作和常规耕作方式木薯根际土壤细菌群落间的α多样性间差异不显著,与高晶晶等[43]、李丽娜等[44]的研究结果一致,说明土壤细菌群落结构的多样性和丰富度差异主要受木薯根系分泌物影响,耕作方式的影响效应较小。本研究PERMANOVA分析结果表明,土壤细菌群落β多样性受耕作因子影响显著,基于未加权Unifrac距离的PERMANOVA分析结果也显示,非根际土壤和根际土壤细菌群落β多样性具有显著差异,与Griffiths和Philippot[45]的研究结果相似,说明常规耕作对木薯土壤耕层的扰动完全破坏了土壤耕作层,而粉垄耕作通过不改变土层的方式深耕深松土壤,仍保留原耕作层的结构,因此,耕作方式对土壤细菌群落系统发育β多样性的影响存在差异。
本研究中,2种耕作方式木薯根际土壤与非根际土壤的优势菌门均为放线菌门、变形菌门、绿弯菌门、厚壁菌门和酸杆菌门,与张贵云等[21]对旱地麦田、周佳等[23]对水稻根际土细菌群落结构的研究结果相似;粉垄耕作方式木薯根际土壤中厚壁菌门芽孢杆菌属(OTU7337)、芽单胞菌门芽单胞菌属(OTU3052)、放线菌门IMCC26256目(OTU9034)及变形菌门Ellin6067属(OTU6431)、A21b科(OTU7905)和马赛菌属(OTU7179)的相对丰度显著大于非根际土壤,结合微生物主要菌群与土壤理化性质的相关分析和RDA分析结果推测,可能与粉垄耕作可减轻木薯涝害,减少根际分泌物有机酸的产生,从而促使木薯根际土壤pH、有机质含量和有效磷含量增加有关[21]。芽孢杆菌属中有很多能降解土壤中苯酚类和酚酸类有毒物质且具有拮抗病原菌活性的益生菌,能改善植物根际生态环境,增强对外界环境的适应和抵御能力[46],可能也是粉垄耕作中根际土壤的芽孢杆菌属显著高于非根际土壤的原因。本研究还发现,常规耕作和粉垄耕作木薯非根际土壤酸杆菌门的相对丰度均高于根际土壤,与柳春林等[47]得出酸杆菌门是具有嗜酸特点的一类细菌,酸性土壤环境有利于其代谢活动的观点一致。
木薯块根形成期正值夏季,雨水较多,而木薯忌渍,土壤中过多的水分会对木薯产生涝害胁迫。粉垄耕作可长时间保持土壤疏松,有利于雨水下渗,可避免或有效减轻木薯在块根形成期遭受涝害胁迫。本研究发现,在木薯非根际土壤中,有机质含量、亚硝酸还原酶活性和硝酸还原酶活性对土壤细菌群落结构的影响较大;在木薯根际土壤中,有效磷含量对土壤各细菌群落结构的影响最大;RDA分析结果表明各细菌群落与环境因子间关系不尽相同,放线菌门的相对丰度与有机质含量和pH呈负相关,与有效磷含量、碱解氮含量和过氧化氢酶活性呈正相关,与Juhnke等[48]的研究结果存在差异,可能是由于植物类型及土壤质地不同所引起;绿弯菌门与pH、有效磷含量和碱解氮含量呈负相关,与Kim等[49]的研究结果一致;厚壁菌门与有机质含量、有效磷含量和碱解氮含量呈正相关,可能与粉垄耕作可优化土壤物理结构、提高土壤有机质含量[50],进而提高厚壁菌门的相对丰度有关;酸杆菌门的相对丰度与有机质含量、pH、有效磷含量和碱解氮含量呈负相关,与Lauber等[51]发现酸杆菌门和放线菌门的相对丰度随着pH的升高而降低的研究结果相似。
4 结 论
木薯粉垄耕作通过改善土壤耕作层物理结构,减少木薯逆境胁迫,使其根际土壤的pH、有机质含量、有效磷含量和速效钾含量大于非根际土壤,进而引起根际土壤细菌群落多样性和丰富度均大于非根际土壤,其中根际土壤细菌群落的α多样性显著高于非根际土壤。木薯根际土壤与非根际土壤细菌群落多样性的差异主要体现在细菌群落相对丰度上,根际土壤中芽孢杆菌属、芽单胞菌属、Ellin6067属和马赛菌属的丰度显著高于非根际土壤。
猜你喜欢
科学技术与工程(2022年26期)2022-11-01
农业工程学报(2022年8期)2022-08-08
中国土壤与肥料(2022年4期)2022-06-14
中国农学通报(2022年14期)2022-06-01
草地学报(2022年4期)2022-04-25
土壤学报(2022年1期)2022-03-08
干旱地区农业研究(2022年1期)2022-01-28
湖北农业科学(2019年22期)2019-12-23
世界热带农业信息(2019年2期)2019-05-17
广东第二课堂·小学(2017年5期)2017-05-27
