
光照强度驱动典型阴生植物三七的生理生态响应特征
2022-05-26 10:02:04双升普张金燕武洪敏孟珍贵李龙根陈军文
生态学报 2022年9期
双升普,张金燕,寸 竹,武洪敏,孟珍贵,李龙根,陈军文,*
1 云南农业大学西南中药材种质创新与利用国家地方联合工程研究中心,昆明 650201 2 云南农业大学云南省药用植物生物学重点实验室,昆明 650201 3 云南农业大学农学与生物技术学院,昆明 650201
光作为光合作用能量的来源,其强度会影响植物的形态建成、光合生理及生物量的产生与分配等过程,从而影响植物生长发育进程[1—2]。而根据植物对光照强度的需求性不同,可以分为喜光植物与喜阴植物[3]。然而在自然界中,光照强度是高度异质化的,植物必须以固着的方式响应光照环境的异质性[4—5]。在没有发生遗传变异的情况下,植物主要通过表型可塑性适应异质化的光环境[6—7]。表型可塑性是植物在形态建成、生长特征和生理功能等方面发生变化从而响应异质环境的一种生态策略[8—10]。可塑性指标的大小可以反映植物对环境变化适应能力的强弱[11—12]。因此,植物表型的可塑性是植物光响应研究的主要指标之一。
光合生理是最先被光照强度驱动来响应变化的生理特征[13—16]。喜光植物通过较高的净光合速率(Pn)、光饱和点(LSP)和光补偿点(LCP)等来增强自身潜在的光合能力,因此能忍受较高的光照强度[17—19]。低光下,喜光植物风吹楠(Horsfieldiaglabra)Pn较低,此时其通过降低LCP和暗呼吸速率(Rd)来减少消耗从而适应遮阴环境;而在高光下可通过增加Pn来适应高光环境[13]。但是,较高的Pn意味着单位面积上需要投入更多的叶绿体、光合蛋白及有机物等物质来构建较厚的叶片[20—21]。喜阴植物由于光合能力较弱,导致用于构建叶片厚度的生物量较少,因此其高光适应能力较弱。另一方面,喜阴植物自身潜在的Pn、LSP、LCP以及Rd相对较低[17—19],当光照强度发生变化时,其光合生理如何在较小的变化幅度内实现最优调整是值得研究的问题。
植物对不同光照强度的响应最终表现在形态建成上。当环境光强减弱时,植物通过增大植株冠幅和增加单片叶面积等方式向着更有利于捕获光能的方向发生改变;而在高光照条件下,植物通过减小单株叶面积和叶片数量等地上部分形态特征来减少光能的吸收[10,22—23]。此外,光照强度对植物地上部分的调节会影响植株碳的获取,从而调节根系生物量的积累,使其更好的适应地上部光环境的变化[1]。前人研究发现喜阴植物可通过形态特征的变化来较快地响应不同的光照环境,但其形态可塑性与喜光植物相比变化幅度较小[10]。因此,了解喜阴植物形态建成如何在有限的变化范围内较快响应异质光环境对于分析其响应策略是非常必要的。
生物量的积累和分配作为植物在异质光环境中内在和外在联系的桥梁,其变化会影响相对生长速率(RGR),是植物响应光照环境的另一重要策略,尤其是不同光照环境下叶生物量比(LMR)、叶面积比(LAR)、比叶面积(SLA)等生物量分配的改变[24—26],其中SLA是与RGR变化相关的主要因子[27—28]。在低光环境下,较大的SLA使得有效叶面积增加,光合碳收获增强,最终引起植物RGR增加[29]。然而SLA对光的适应性调整也是有限度的,当遮阴过度时喜阴植物总的生物量减少,此时分配到地上部分的生物量虽然比例有所增加但数量却有所减少,使得植物捕获的光能相对较少,导致RGR降低。生产实践上,都会追求植物的最大RGR,以达到高产目的,因此,当环境光强发生变化时,阴生植物生物量如何优化分配调整来使RGR尤其是经济器官的RGR保持较大是十分重要的。
三七(Panaxnotoginseng)为五加科人参属多年生的典型阴生植物,以根和根状茎入药,是中国传统的名贵中药材,在种植过程中需要采用遮阴栽培[30—31]。传统道地产区的三七种植中有“三分阳,七分阴”之说,认为三七在30%透光率(LT)下生长较好,但试验证据相对缺乏。本课题组前期研究发现,当透光率为9.6%—11.5%时,三七具有较高的Pn,同时在1%透光率下三七也能生存,表现出较强适应低光的能力[31—33]。此外,有研究发现,一年生三七通过提高Pn、降低蒸腾速率(Tr)和气孔导度(Gs)以及降低株高和叶面积来适应高光照环境[34—35],但实验使用的材料是处于种苗阶段的一年生三七。本课题组研究还发现,两年生三七会通过降低根冠比(RSR)、根生物量比(RMR),增加茎生物量比(SMR)和LMR以获取更多的光能,从而更好地适应低光照环境;而在高光照下则是通过叶片变厚变小来减少光能捕获,并增加根重比来适应环境[36—38]。然而,以往的相关工作都是对不同生长光照强度下三七形态特征或生理生态特性中某一方面的研究,从形态、生理和生长角度综合深入分析三七对长期光照环境变化的响应特征的研究相对较少,且以前相关研究生长环境光强设置的范围较小。因此,本文以两年生三七为实验材料,设置一系列的生长环境光照强度(46.5% LT、21.8% LT、9.70% LT、5.10% LT和2.80% LT),对生长于不同光照强度下三七的形态特征、生物量积累及分配和光合生理特性进行分析,并进一步进行相关性、可塑性和主成分分析,进而阐明三七对不同光照环境的响应策略,为三七的优质高效种植提供理论依据。
1 材料和方法
1.1 试验地概况
试验设于文山州砚山县盘龙乡(104°32′E,23°53′N)苗乡三七实业有限公司三七科技示范园内。砚山县位于云南省东南部,103°35′—104°45′E、23°18′—23°59′N之间,海拔1539 m,年平均降雨量1200 mm,年平均气温约16 ℃,最冷月平均温度9 ℃,最热月平均温度21 ℃,全年无霜期大于300 d,日均气温10 ℃以上,积温为5000 ℃,属于亚热带大陆性季风气候[39]。砚山县是三七的主要传统产区和道地产区,同时也是三七最适宜生长的地区之一,被誉为“中国三七之乡”。
1.2 试验材料及设计
在空旷地使用三七专用遮阳网搭建5种不同透光率的荫棚,荫棚长4 m、宽4 m、高2 m,设有纱网,通风透气。选择晴天,使用Li-190R 量子传感器(棚内和棚外各三个)和Li-1500 光量子记录仪(Li-Cor,USA),于6:00—19:00每隔10 s采集一次每个荫棚内的光照强度;同时测量全日照强度作为对照。以测量时间段内荫棚光照强度(三个量子传感器的平均值)占同一时刻透光率强度百分比的平均值作为三七生长环境光强的指标;经测量,5个处理的平均环境光照强度分别为:46.5% LT、21.8% LT、9.70% LT、5.10% LT、2.80% LT(图1)。在前期研究发现,透光率为9.6%—11.5%时三七生长发育良好,透光率高于30%三七就会受到显著的高光抑制,而透光率只有3%左右时三七将受到明显的低光限制[36],因此,本实验中9.70%透光率代表前期发现的适宜光照条件,5.10%、2.80%的透光率代表低光条件,46.5%和21.8%的透光率代表高光条件。实验采取盆栽进行,花盆内径30 cm、深25 cm,盆栽土壤为当地生土(红壤),其土壤基本理化性状如表1所示。于1月份选取生长良好、大小一致的三七种苗进行移栽,每个处理下划分为15个重复小区,每个小区放置5盆,每盆3株。试验期间三七常规田间管理措施统一按文山三七科技示范园的三七规范化种植技术规程进行。

图1 不同透光率和全日照下的光量子通量密度Fig.1 Representative diurnal time courses of photosynthetic photon flux density in different light transmittance (LT) and the full sunlight
1.3 测定指标及方法
1.3.1形态指标的测量
于12月份三七(两年生)收获期,将不同透光率荫棚下的三七进行取样。取样时,将三七整株取出,用水洗净,每个处理选15株(每个小区1株)进行测量,每株重复3次。用电子数显游标卡尺测量茎粗(mm);用直尺测量株高(cm)、每片小叶的长和宽(测叶片最宽的地方,cm),将叶片看作椭圆形,计算每个叶片的叶面积(叶面积=叶长×叶宽×0.75 cm2)[40],并统计出三七的单株叶面积(cm2);然后统计植株的叶片数及分枝数。
1.3.2生物量及分配的测量
每个处理选15株三七(每个小区1株),然后将根、茎、叶分开放在105 ℃下的烘箱中杀青1 h,然后在60 ℃下烘干至恒重。用电子天平称量根生物量(根重,g)、茎生物量(茎重,g)、叶生物量(叶重,g);并统计出总生物量(Mt)、根生物量比(RMR,根重/植株总重)、茎生物量比(SMR,茎重/植株总重)、叶生物量比(LMR,叶重/植株总重)、叶面积比(LAR,总叶面积/植株总重cm2/g)和比叶面积(SLA,总叶面积/叶重cm2/g)。并且在移栽时也需要对三七种苗进行取样,烘干后测量其总生物量(M0),用于计算相对生长速率(RGR)[41]:
RGR=(lnMt-lnM0)/t
其中,Mt表示两年生三七的总生物量(g),M0表示移栽时测定的三七种苗的总生物量(g),t表示两次测定的时间间隔(d)。
1.3.3气体交换参数的测量
于生长旺盛期,使用Li-6400XT光合-荧光记录仪(Li-Cor,USA)测定不同荫棚下三七叶片的光响应曲线。使用CO2注入系统将CO2浓度控制为400 μmol/mol,夹上叶片后于500 μmol m-2s-1光强下稳定一段时间,待数据稳定后启动自动测量程序,光照强度从大到小依次设为:800、500、400、300、200、150、100、80、60、40、20、10、0 μmol m-2s-1,每个光强下稳定2—3 min,采集气体交换参数值。以光照强度(Photosynthetic photon flux density,PPFD)为横轴,Pn为纵轴绘制光响应曲线(Pn-PPFD曲线)。据Bassman的方法拟合Pn-PPFD曲线方程[42]:
Pn=Pn-max-Pn-maxC0e-αPPFD/Pn-max
式中,Pn-max为最大净光合速率,α为表观量子效率(AQY),C0为度量弱光下净光合速率趋近于0的指标。从中可得到单位面积最大净光合速率(Pn-max/area)、单位面积气孔导度(Gs/area)、单位面积暗呼吸速率(Rd/area)=Pn-max-Pn-maxC0、单位面积光补偿点(LCP/area,μmol m-2s-1)=Pn-maxln(C0)/α。然后基于SLA计算单位质量最大净光合速率(Pn-max/mass)=(Pn-max/area)×SLA×1000/10000、单位质量气孔导度(Gs/mass)=(Gs/area)×SLA×1000/10000、单位质量暗呼吸速率(Rd/mass)=(Rd/area)×SLA×1000/10000、单位质量光补偿点(LCP/mass,nmol g-1s-1)=(LCP/area)×SLA×1000/10000。实验中使用统一校准后随机分配的5台Li-6400XT光合仪(Li-Cor,USA)同时测量5个处理,每个处理选取15株(每个小区1株)进行测量,每株选取1叶片,叶片空间取向和角度尽量一致。
1.4 数据处理
用SPSS 25.0软件对数据进行单因素方差分析(One-way ANOVA)及差异显著性分析(n=15),用Pearson法对三七RGR与各指标间的相关性进行分析;用Sigmaplot 14.0和Origin 2020b软件作图。
2 结果与分析
2.1 不同光照强度对三七生长及形态特征的影响
在高光和低光下,三七各形态特征参数都有所降低(图2)。其中,株高、茎粗和单株叶面积在透光率为9.70% 时最高,且显著高于46.5% 和2.80% (P<0.05);叶片数量和分枝数量在透光率为9.70% 的条件下最多,且显著高于46.5% (P<0.05)。总的来看,在透光率为9.70% 时三七形态特征最好,高光或低光都会显著降低其形态的生长发育。
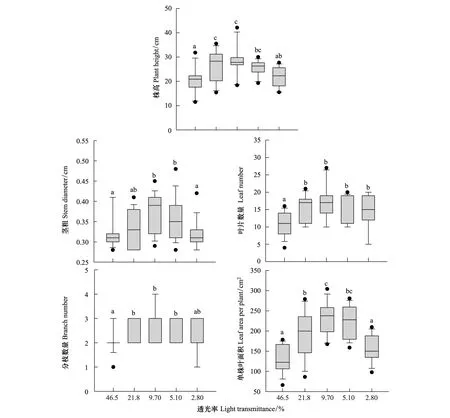
图2 不同透光率对三七株高、茎粗、叶片数量、分枝数量和单株叶面积的影响Fig.2 Effects of different light transmittance on plant height,stem diameter,leaf number,branche number and leaf area per plant of P. notoginseng不同字母表示差异显著(P<0.05)
2.2 不同光照强度对三七生物量分配的影响
高光和低光条件下三七各部分生物量都有所降低(图3)。单株根生物量、单株茎生物量、单株叶生物量和单株总生物量在不同透光率下的变化趋势一致,都在透光率为9.70%时最高,其次是21.8%,这两种条件的各部分生物量都显著高于其他透光率(P<0.05),且透光率为2.80%时各部分生物量最少。总的看来,透光率为9.70%条件下的各部分生物量和单株的总生物量都具有最大值,而透光率为2.80%时最低,并且显著低于其他几个透光率(P<0.05),表明低光下生物量积累较少。

图3 不同透光率对三七单株根生物量、单株茎生物量、单株叶生物量和单株总生物量的影响Fig.3 Effects of different light transmittance on root biomass per plant,stem biomass per plant,leaf biomass per plant and total biomass per plant of P. notoginseng
由图4可知,三七LAR和SLA都在透光率为2.80%下最大,且显著高于其他几个透光率(P<0.05),表明低光下植物会通过增加LAR和SLA来获取更多的光能。另外,不同透光率下三七RMR、SMR和LMR是存在一定差异的。其中,RMR随着透光率的降低而降低,而SMR和LMR则是随着透光率的降低而逐渐增加,且三者在透光率为46.5%、21.8%和9.70%时差异都不显著,但都显著低于其他两个透光率;并且RGR在透光率为9.70%时最大,并显著高于其他几个透光率(P<0.05)。表明低光和高光都会影响植物各部分生物量的分配比例,并抑制植物生长。

图4 不同透光率对三七生物量分配的影响Fig.4 Effects of different light transmittance on biomass allocation of P. notoginseng
2.3 不同光照强度对三七光合能力的影响
由图5可以看出,不同透光率下三七Pn-max/area、Gs/area、Rd/area、LCP/area和Pn-max/mass、Gs/mass、Rd/mass、LCP/mass是存在差异的。与透光率9.70%相比,高光(46.5% LT和21.8% LT)水平下的Pn-max/area、Gs/area、LCP/area和Pn-max/mass、Gs/mass、LCP/mass略高,但与对照差异不显著;低光(5.10% LT和2.80% LT)水平下,上述参数则显著低于9.70% LT下的(P<0.05),且随着透光率的降低而减小。而Rd/area和Rd/mass都是随着透光率的降低而逐渐上升,与透光率9.70%相比,除21.8% LT条件下的Rd/mass与其差异性不显著外,其余参数在高光下都显著下降(P<0.05);而低光下则都显著较高(P<0.05)。这些结果表明9.70%的透光率下三七的光合能力较好,而高光下三七会制造和消耗较多的有机物,低光下有机物的制造和消耗较少。

图5 不同透光率对三七光合生理的影响Fig.5 Effects of different light transmittance on photosynthetic physiology of P. notoginsengPn-max/area:单位面积最大净光合速率Maximum net photosynthetic rate unit area;Gs/area:单位面积气孔导度Stomatal conductance unit area;Rd/area:单位面积暗呼吸速率Dark respiration rate unit area;LCP/area:单位面积光补偿点Light compensation point unit area;Pn-max /mass:单位质量最大净光合速率Maximum net photosynthetic rate unit mass;Gs/mass:单位质量气孔导度Stomatal conductance unit mass;Rd/mass:单位质量暗呼吸速率Dark respiration rate unit mass;LCP/mass:单位质量光补偿点Light compensation point unit mass
2.4 不同光照强度下三七形态和生理特征与RGR的相关性分析
2.4.1形态特征与RGR的相关性分析
RGR与株高、茎粗以及单株叶面积呈正相关关系,相关性均达到极显著水平(P<0.01),但相关性系数较低,分别为0.116、0.093和0.270,其中RGR与单株叶面积相关性最大,而与叶片数量的相关性则不显著(图6)。这些结果说明,在形态特征方面,单株叶面积是影响三七生长发育的主要因素,但形态特征各参数的相关性系数相对较小,表明与光合生理及生物量分配相比,形态特征对三七生长发育的影响较弱。

图6 三七株高、茎粗、叶片数量、单株叶面积与相对生长速率的相关性Fig.6 Correlation between plant height,stem diameter,leaf number,leaf area per plant and relative growth rate (RGR) of P. notoginseng
2.4.2生物量分配与RGR的相关性分析
RGR与LAR、SLA、SMR、LMR呈负相关关系,相关性均达到极显著水平(P<0.01),其相关系数分别为0.493、0.461、0.390、0.472,而与RMR则是极显著的正相关关系(P<0.01)相关系数为0.553。说明RGR与RMR相关性最大,和LAR的相关性次之(图7)。总的来看,生物量分配对三七的生长发育有着较大的影响,其中又以控制分配到根部生物量的RMR和调控有机物生产和积累的LAR的影响最大。说明生物量分配中三七主要通过RMR和LAR来响应不同光照强度。

图7 三七叶面积比、比叶面积、根生物量比、茎生物量比、叶生物量比与相对生长速率的相关性Fig.7 Correlation between leaf area rate,specific leaf area,root mass ratio,stem mass ratio,leaf mass ratio and relative growth rate of P. notoginseng
2.4.3气体交换参数与RGR的相关性分析
由图8可知,RGR与Pn-max/area、Gs/area、LCP/area、Pn-max/mass、Gs/mass、LCP/mass呈正相关关系,相关性均达到极显著水平(P<0.01),相关系数分别为0.581、0.558、0.574、0.350、0.274、0.300,而与Rd/area、Rd/mass则是极显著的负相关关系(P<0.01)相关系数分别是0.472、0.333;说明RGR与Pn-max/area相关性最大,和Gs/area、LCP/area的相关性次之。总之,三七的RGR与Pn、Gs、LCP、Rd有很大的关联,说明光合生理是植物响应光照环境的一个重要策略,是影响三七生长发育的主要因素。

图8 三七单位面积最大净光合速率(Pn-max/area)、单位面积气孔导度(Gs/area)、单位面积暗呼吸速率(Rd/area)、单位面积光补偿点(LCP/area)和单位质量最大净光合速率(Pn-max/mass)、单位质量气孔导度(Gs/mass)、单位质量暗呼吸速率(Rd/mass)、单位质量光补偿点(LCP/mass)与相对生长速率的相关性Fig.8 Correlation between maximum net photosynthetic rate unit area (Pn-max/area),stomatal conductance unit area (Gs/area),dark respiration rate unit area (Rd/area),light compensation point unit area (LCP/area),maximum net photosynthetic rate unit mass (Pn-max /mass),stomatal conductance unit mass (Gs/mass),dark respiration rate unit mass (Rd/mass),light compensation point unit mass (LCP/mass) and relative growth rate of P. notoginseng
2.5 不同光照强度下三七形态和生理特征的可塑性
由图9可知,在所有性状指标中,茎粗的可塑性指数最小,单株根生物量可塑性指数最大;从形态特征、生长特征和生理特征三方面来看,形态特征中没有一个参数的可塑性指数大于0.5,其可塑性最小,生理特征参数的可塑性指数大于0.5的最多,其可塑性最大;而从形态特征分组可以看出,各参数的可塑性指数均小于0.5,其中单株叶面积的可塑性最高;在生长特征分组中,单株根生物量及LAR的可塑性指数大于0.5,其中单株根生物量的可塑性最大,RGR的可塑性最小;生理特征中可塑性指数大于0.5的参数最多,有5项(Pn-max/area、Gs/area、Rd/area、LCP/area、Rd/mass)占比62.5%,其中Rd/area的可塑性最大,Gs/mass的可塑性最小。这些结果表明三七主要是通过改变光合生理、根的生物量以及LAR来响应光照强度的变化。
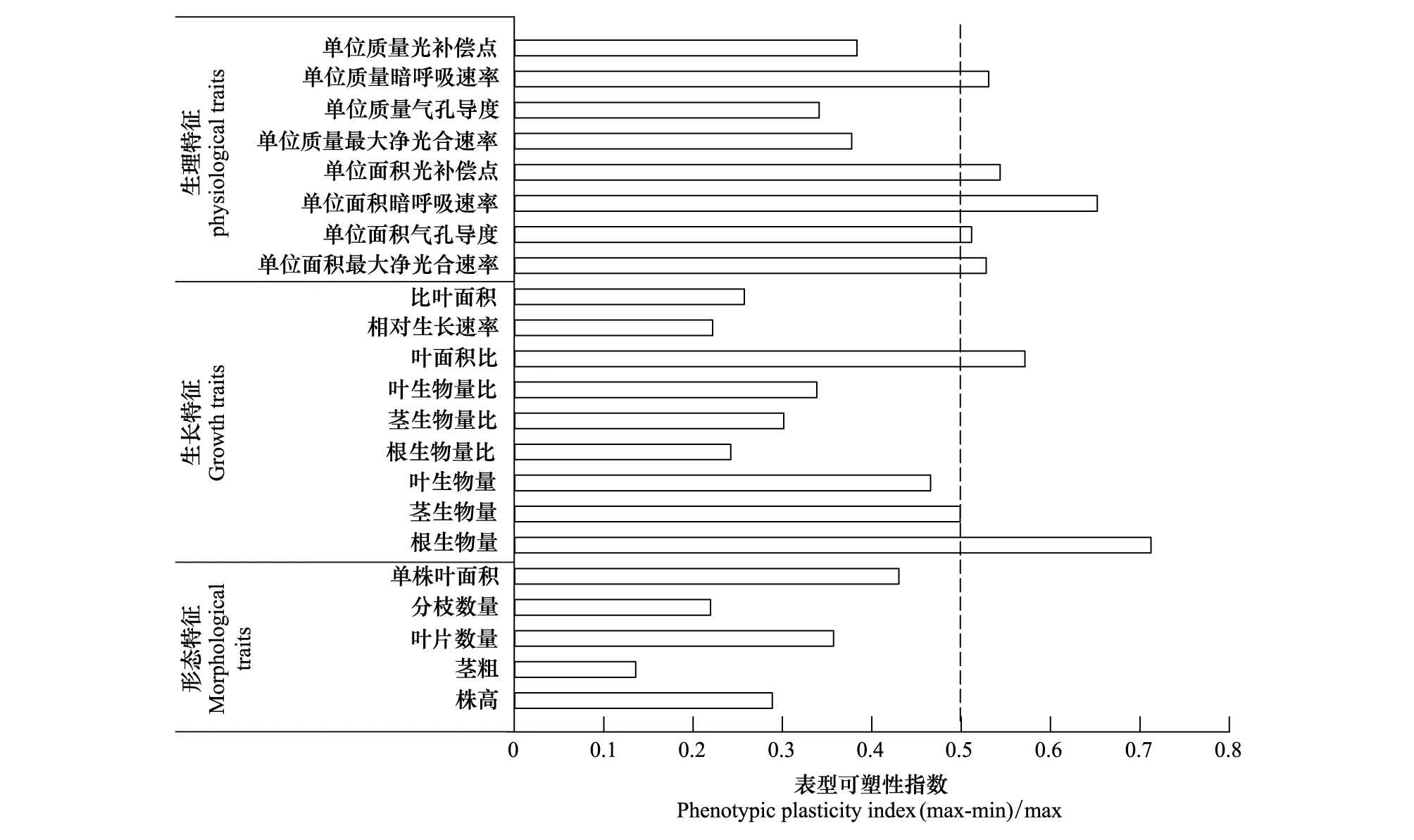
图9 三七形态特征、生长特征和生理特征等各参数的表型可塑性指数Fig.9 Phenotypic plasticity index of parameters of morphological characteristics,growth characteristics,and physiological characteristics of P. notoginseng
2.6 不同光强响应特征的主成分分析
对不同光照强度下三七的23个形态特征、生长特征和生理特征参数进行主成分分析,得到各主成分对三七响应不同光强的特征值和贡献率(表2)。从表中可以看出,PC1和PC2的累计贡献度达70.61%,且特征

表2 各成分初始特征值及累积贡献率Table 2 The initial eigenvalues and the accumulated variance contribution of each component
值都大于1,因此这两个主成分能有效反映参数特征和有效解释三七的响应特征。
由图10可知,PC1中光合生理特征参数(Pn-max/area、Gs/area、Rd/area、LCP/area、Pn-max/mass、Gs/mass、Rd/mass、LCP/mass)和生长特征参数(单株根生物、单株总生物量、RMR、SMR、LMR、LAR、SLA、RGR)权重系数较大;PC2中生长特征参数(单株茎生物量、单株叶生物量)和形态特征参数(株高、茎粗、叶片数量、分枝数量、单株叶面积)权重系数较大。实验点在PC1/PC2平面上较好的集聚为五簇,分别代表五个不同光强处理。同时PC1非常明显的将透光率2.80%与其他四个透光率分离开,将透光率5.10%与透光率46.5%、21.8%、9.70%明显分离开,而透光率46.5%、21.8%、9.70%三者间没有明显的区分,表明低光与其他几个光强之间有着明显的差异,而高光与透光率9.70%的差异较小。由载荷系数可以看出,Pn-max/area、Gs/area、LCP/area、RGR、根生物量、总生物量与PC1有着正相关关系,且对PC1的贡献较大;而LAR、Rd/area对PC1也有着较大的贡献,但为负相关关系;叶片数量、分枝数量、单株叶面积与PC2为负相关关系,并且对其贡献较大。并且Pn-max/area、Gs/area、LCP/area、RMR、根部生物量及总生物量与RGR相关性较高,且为正相关关系,而Rd/area、LAR与RGR则有着较高的负相关关系,说明在不同光照强度下三七主要靠调整光合生理特征、LAR、根生物量以及RMR来响应变化。

图10 不同透光率下三七形态和生理指标的主成分分析Fig.10 Principal component analysis of morphological and physiological indexes of P. notoginseng under different light transmittance LAR:叶面积比Leaf area rate;SLA:比叶面积Specific leaf area;RMR:根生物量比Root mass ratio;SMR:茎生物量比Stem mass ratio;LMR:叶生物量比Leaf mass ratio;图中点的颜色代表不同透光率;椭圆为不同透光率下各参数的置信区间;箭头代表各指标与主成分的关系
3 讨论
3.1 高光环境下阴生植物三七采取快速碳获得和碳消耗的冒险响应策略
形态特征作为判断植物生长发育状况的一项重要指标,可以直观反映植株对不同强度光照环境的适应性[43]。植物形态特征的改变是通过生物量积累和局部特化的生物量分配变化来实现的,因此,生物量在各器官间的分配反映植物生长策略的调整[22,44]。高光环境下阴生植物三七LMR、SMR、SLA和LAR较低,RMR较高(图4),这与魏巍等研究发现高光下阳生植物薇甘菊(Mikaniamicrantha)SLA 、LAR降低一致[24]。较低的SLA和LAR使叶片变厚变小,即单位生物量所承载的光合叶面积减少,使其捕光面积降低[45]。较厚的叶片在强光下单位面积叶片的光合能力较强[46],从而三七在高光下保持较高的Pn(图5),消耗更多的光能,生产更多的有机物。同时高光下三七Rd较大(图5),使得有机物消耗较多,从而维持Pn在一个较高水平来利用更多的光能,防止光合机构遭受破坏[42,47],这些特征有利于植物在强光下保持较高的生存能力,提高对强光的适应性。与此同时,SLA和LAR降低表明植物生物量分配发生变化,高光下较多的生物量分配到三七地下部分,抑制地上部分的生长,提高RMR,有利于吸收更多的水分和无机物,为植物蒸腾和光合作用提供更多的原料,这与叶片变小时三七蒸腾失水减少相互协调,成为阴生植物三七响应高光环境的重要策略[45,48]。总的来看,高光下三七较高的Pn可以制造更多的有机物,而Rd较大则会消耗大量的光合产物,并且此时生物量积累较少(图3),这说明高光下阴生植物三七同时进行快速碳获得和碳消耗的冒险策略[49]。
3.2 低光下阴生植物三七采取缓慢碳获得和碳消耗的保守响应策略
低光下由于光能不足,导致碳获得显著下降(图5),此时各部分的生物量和总生物量的积累都显著降低(图3),这与阳生植物大豆(Glycinemax)在遮阴处理下光合作用减弱,导致生物量积累减少的结果相一致[50]。遮阴过度时,三七吸收的光能无法满足其正常光合作用所需,因此Pn降低,产生和积累的有机物减少,营养生长受到抑制,最终导致株高和叶面积降低(图2),此时Rd降低,有机物的消耗减少,这与较低的光合速率相匹配。此外,三七通过减少根生物量的分配,将更多的资源分配到地上的茎和叶部分,增加SLA和LAR(图4),植株叶面积增大,使其在低光下捕获更多的光能,合成更多的光合产物来提高其耐阴性,以此来实现对弱光环境的适应[51—53]。另一方面,LCP的高低可以反映植物对弱光的利用能力,低光下三七具有较低的LCP,降低了对光强利用和净碳获得的阈值,这也是植物对低光环境的一种响应策略。因此,低光下阴生植物三七通过减少呼吸消耗来应对较少的光合有机物产量,从而维持碳平衡,此时,三七采取缓慢的碳获得和碳消耗的保守策略来响应低光环境。
3.3 阴生植物三七光合生理活性及生物量分配的改变是其响应光强变化的主要策略
RGR与植物形态特征、生理特征及生物量的积累与分配等有着密切关系。前人研究发现,RGR受光合碳获得和LAR的共同影响,较高的Pn和LAR均可提高RGR[54]。在本研究中,Pn、Gs和LCP与RGR呈正相关关系,Rd与RGR呈负相关关系(图8),这与前人研究发现,阳生植物麻栎(Quercusacutissima)有着较高的Pn和较低的Rd从而使得生长速率较快的结果相一致,这是因为高的Pn能够使植物制造更多的光合产物,低的Rd则有利于减少对有机物的消耗,最终获得最大的碳积累,使植物生物量积累和生长更快[19,55]。但在本研究中LAR与RGR呈极显著的负相关关系(图7),这与喜光植物桤木(Alnuscremastogyne)较大的LAR使其表现出较高的RGR有所差异[56],可能是因为阴生植物三七为了适应低光照环境而增大LAR,但此时光照强度过于偏低,植物光合效率较低,使得有机物的产生和积累减少,从而导致RGR降低。此外,本研究中RMR与RGR呈极显著的正相关关系(图7),较高RMR意味着植物将更多的生物量分配到根部,使得根部生长发育健壮,能吸取更多的水分和养分,而根部通常被认为是用于储存有机碳的器官,增强RMR有利于有机物的积累[57],且三七是根茎类药材,因此RMR与RGR有着较大的相关性。总之,阴生植物三七的Pn、Gs、LCP、Rd、LAR和RMR对RGR的影响较大。
植物依赖于自身的表型可塑性来调整对自然界中各种资源的获取和消耗,以维持自身正常的生长和代谢,从而响应不同的环境条件[58]。可塑性指数可以直观反映植物对环境变化适应能力的大小[8—10]。研究发现,阳生植物比阴生植物表现出较大的表型可塑性,且可塑性方式存在差异[59]。张常隆等研究表明,阳生植物紫茎泽兰(Ageratinaadenophora)的叶面积、株高、冠宽、分枝数、LAR以及茎、叶生物量等指标的可塑性指数较大,这说明其主要通过调节植株形态特征以及生物量积累与分配来响应不同的环境[7]。本研究中,由可塑性分析(图9)和主成分分析(图10)发现,典型阴生植物三七与阳生植物有所差异,三七的形态特征可塑性相对较小,与RGR相关性不显著,而其光合生理(Pn-max/area、Gs/area、Rd/area、LCP/area)、LAR和根生物量可塑性指数较高,在主成分上的载荷较大。这是因为三七叶片形态、光合生理及生物量分配对光强的变化是十分敏感的,尤其是光合生理(Pn-max/area、Gs/area、Rd/area、LCP/area)和LAR。而三七根部作为碳储存的器官和经济器官,其生物量的积累对环境的变化也较为敏感。总的来看,阴生植物主要依赖于光合生理及生物量分配的改变来响应光环境的变化。
3.4 10% LT左右的光照条件最有利于三七生长发育
三七作为一种典型的阴生植物,其对光照环境十分敏感[60],在有关三七栽培的资料中,有人认为“三分阳、七分阴”(即透光率在30%左右)的光照条件适宜三七的生长[35]。另外,早期也有研究指出三七生长适宜的透光率为40%—60%[62];而前期研究认为,从光合生理角度判断,当透光率为9.6%—11.5%时,三七的光合生理表现较好[31—33]。本研究结果表明,三七在透光率为9.70%时,株高、茎粗和单株叶面积等形态特征明显优于其他光照条件下的,同时各部分生物量和总生物量积累最高,高光或低光都会显著影响其形态的生长发育和生物量的积累(图2、图3)。此外,与透光率9.70%相比,高光照水平下植株的Pn-max/area、Gs/area、LCP/area虽差异不显著,但仍表现出一个较低的值;透光率低于9.7%时,随着透光率降低,净光合速率明显减小。从形态特征、光合生理及生长特征综合看来,阴生植物三七在透光率9.70%时各特征都表现优良。因此,建议在三七种植实践中应该采取“一分阳,九分阴”(即透光率为10%左右)的光照强度管理措施。
4 结论
综上所述,在光照强度变化的生长环境中,阴生植物三七形态可塑性较小,光合生理和生物量分配可塑性较大,且Pn-max、Gs、LCP、LAR与RGR相关性较强,表明三七光合生理特性、LAR和根生物量的变化是光照强度驱动的主要响应特征。综合看来,9.70%左右的透光率下三七生长发育良好,因此在栽培管理中建议采取透光率为10%左右的光照环境,即“一分阳,九分阴”。对于不利的生长环境光照强度,阴生植物三七通过提升捕光能力和采取缓慢的碳获得和碳消耗的保守策略适应低光环境;在高光条件下则会降低捕光面积、增加根部生物量,并采取快速的碳获得和碳消耗的冒险策略。
猜你喜欢
睿士(2023年8期)2023-08-23 13:07:06
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
河北果树(2022年1期)2022-02-16 00:41:04
北京农学院学报(2021年4期)2021-11-09 00:43:20
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
玩具世界(2021年5期)2021-03-08 08:42:02
发光学报(2019年8期)2019-08-20 10:15:38
家庭影院技术(2017年12期)2017-02-06 02:32:05
西南医科大学学报(2015年1期)2015-08-22 13:01:48
